长山群岛藓类植物多样性研究
2022-03-09宋玉玲李丹丹于晶
宋玉玲 李丹丹 于晶


摘 要: 于2019年10月,对辽宁省大连市长山群岛进行野外实地调查,共采集藓类植物标本259份.对采集到的标本进行鉴定分析,结果表明:长山群岛共有藓类植物96种,14科,38属,藓类植物种类组成分布的科、属比较集中,在科属组成分析中,优势科为丛藓科(Pottiaceae)、灰藓科(Hypnaceae)、曲尾藓科(Dicranaceae)、金发藓科(Polytrichaceae)、葫芦藓科(Funariaceae).土生藓类占总数的81.65%.区系类型以温带成分为主,具有强烈的温带性质.通过与其他地区藓类植物属和种的相似性比较,发现长山群岛藓类植物与其相近的地区植物的相似性更高一些,并且海岛藓类植物之间的相似性比与其他距离较近的陆地环境中的高.
关键词: 长山群岛; 藓类植物; 多样性; 区系分析; 相似性
中图分类号: Q 949.1 文献标志码: A 文章编号: 1000-5137(2022)01-0094-07
In 2019, a field survey was conducted on the Changshan Islands, in which more than 259 moss specimens were collected. There are totally 96 moss species on Changshan Islands, which belong to 14 families, 38 genera, according to specimen identification. The distribution of families and genera composition of mosses are centralized relatively. The analysis on the composition of moss family and genus shows that the dominant families are: Pottiaceae, Hypnaceae, Dicranaceae, Polytrichaceae, and Funariaceae. Soil-born mosses take the main proportion, accounting for 81.65% of the total. The dominated flora element is the temperate one, with a strong temperate nature. Compared with the mosses in other regions, the mosses of the Changshan Islands are more similar to those in its neighboring areas in genera and species. The similarity of mosses from different islands is higher than the same factor of mosses between island and closer terrestrial environment.
Changshan Islands; moss; diversity; floristic analysis; similarity
长山群岛位于辽宁省大连市,四季分明,气候适宜.年平均降水量在550~950 mm之间,降水基本集中在夏季.苔藓植物是特殊的高等植物,与其他种子植物相比,苔藓植物结构简单、形体矮小,经常被人们所忽视,但其在园林设计、药用、能源开发等领域都发挥着很大的作用.从分布上来看,苔藓植物分布范围广泛,从南到北都有其存在,是生物多样性的组成成分之一.目前,大部分关于苔藓植物的研究集中在南方陆地环境的城市,如上海、蘇州等地,有关长山群岛的苔藓植物研究报道相对较少.因此,本研究选取了长山群岛中相对较大的岛屿:大、小长山岛、广鹿岛为研究对象,调查了长山群岛苔藓植物物种多样性及特点,为海岛的苔藓植物多样性保护与利用提供基础资料.
1 研究区域与研究方法
研究区域概况
长山群岛是中国东北地区唯一岛屿,中国八大岛之一,毗邻大连市区,东部与朝鲜半岛相望,西面紧邻辽东半岛,南部与山东半岛相对,整个岛屿由140多个岛砣礁组成,呈东西排列.长山群岛总陆地面积119 km,海域面积7 720 km,岛屿岸线约428.5 km,海岸系数约为2.81.其中大长山岛陆域面积最大,约30 km.
研究方法
1.2.1 标本采集
2019年10月赴大连长山群岛进行野外考察和标本采集,采集地点见图1,共采集苔藓植物标本259余份,自然风干保存.
1.2.2 标本鉴定
在实验室内,参考相关苔藓植物志等有关资料,并借助Olympus SZ40光学解剖镜和Olympus CX21光学显微镜对苔藓植物标本进行了形态学观察及分类鉴定.
1.2.3 数据分析
在整理长山群岛藓类植物名录的基础上,对藓类植物物种组成、优势科属种进行分析.参考相关的苔藓植物区系资料,确定长山群岛的区系成分.并从国内选取其他几个区域,计算长山群岛与其他地区藓类植物属和种的区域相似系数,计算公式为:
其中,为A地区全部种或者属数;为B地区全部种或者属数;为A,B两个地区共有的种属数.
2 结果与分析
藓类植物多样性及特点
2.1.1 藓类植物种类组成
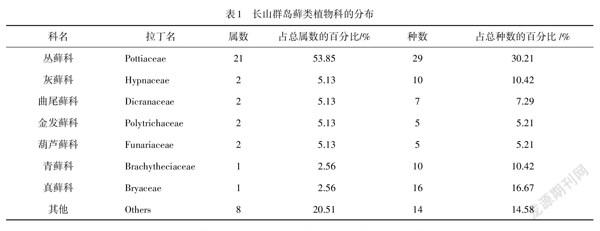
鉴定结果(表1)表明:长山群岛共有藓类植物14科38属96种,其中含5个种以上的共7个科,包含31属、82种,分别占总调查藓类属数和总种数的79.49%和85.42%,所占比例较大,说明长山群岛藓类植物种类分布的科、属比较集中.
由表1可知:在长山群岛中丛藓科(Pottiaceae)植物占比较大.GAO等研究发现:丛藓科所含属、种丰富,多为旱生生境,与长山群岛气候干旱、比较缺水的特点相符.由于地下水是海岛重要的供水水源,海岛地下水环境功能极为重要.长山群岛水环境容量小、径流短,地下水系只有季节性溪流,降水分布不均匀,一般集中在夏季,但因地下蓄水条件差,雨水大量流失,加上超量开采地下水,存在海水入侵的危险,群岛境内多数属于贫水区.在长山群岛藓类植物物种组成中,既有灰藓科(Hypnaceae)等北方常见科,也有如丛藓科(Pottiaceae)、真藓科(Bryaceae)的世界广布科.真藓科(Bryaceae)分布范围广,适应能力强,常常分布于高山、丘陵,也分布于比较湿润的荫蔽环境中,比如林下、房前屋后、路边阴湿的土地等.还有曲尾藓科(Dicranaceae)等典型的热带科.北方城市相对于南方城市气候较干燥,丛藓科和真藓科等藓类更适应北方城市的气候.
经查阅资料并结合实地考察,发现大长山岛、小长山岛处于整个长山群岛的中部,岛狭长,山地丘陵区植被整体保持较好,总体的植被景观格局特征为:景观破碎度指数大,景观多样性指数相对比较居中,空间聚集度指数小.广鹿岛位于群岛的西部位置,地势比较平坦,岛上有一定数量的农田植被,植被稳定性低.随着经济的发展,人类活动对植被景观的破坏程度越来越严重.岛屿本身的生存系统存在着一定的脆弱性,抵御外来因素造成的生态污染的能力较弱,一旦遭到破坏,恢复较慢.有研究对苔藓植物多样性的方面进行调查,发现较大面积的地表,其苔蘚植物分布较少,主要原因可能是湿度较低和森林覆盖率不够.而在比较湿润的环境,如树林下等比较干燥的区域,土生和石生的苔藓植物种类多. 近几年,长山群岛整体上降雨量小,地质比较干旱.自然环境条件不稳定,可能对苔藓植物的生长分布起到了关键作用.总体来说,长山岛藓类植物数量和种类比较少,并且各个岛的优势科和优势属相似,说明各岛屿的气候特征、地质历史、人文情貌、经济发展和植被演化的过程有着密切的联系.
2.1.2 生境特点
生物与环境的关系是长期进化的结果,不同的生境有着不同的物种.苔藓植物根据生境一般可以分为土生、石生等类型.通过对在长山群岛中采集的标本的生境进行统计分析,总结了藓类植物在长山群岛分布的主要特点.
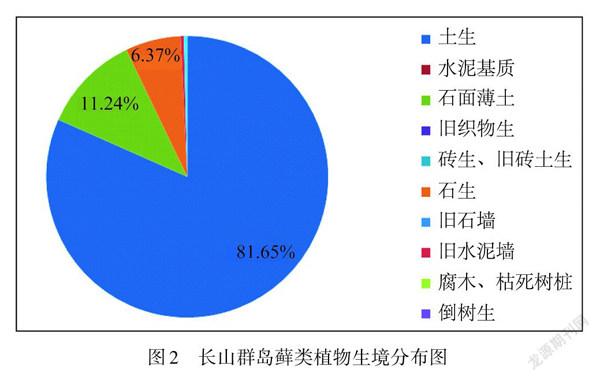
由图2可知:长山群岛以土生藓类为主,占总种数的81.65%,石面薄土和石生种类分别占总种数的11.24%和6.37%,没有树生种类.影响树生苔藓植物分布的主要因素众说不一,包括木本植物的多样性、局部的气候条件、森林郁闭度引起的空气湿度差异等.
经过调查发现:长山群岛植被属于暖温带夏绿阔叶林带,从属于华北植物区系,苔藓植物在树干上的分布高度远没有我国南方山地森林上的高,并且由于长山群岛的开发历史比较早,许多植物遭受到破坏,现在的植被多为天然次生林和人工次生林,因此,在长山群岛几乎没有树附生苔藓分布.
区系成分分析
参照WU等的相关研究资料,将长山群岛藓类植物区系进行划分,统计结果见表2.由表2可知:长山群岛苔藓植物区系成分主要是北温带成分和东亚成分,分别为81种和79种,占总种数的31.76%和30.98%,说明本区域藓类植物属温带性质,与其地理位置和气候特点相一致.部分藓类植物区系的主体具有显著的东亚特点.
长山群岛与其他地区藓类植物属和种的相似性
選择与大连相邻的辽宁省沈阳市、辽宁省仙人洞国家自然保护区,位于长江三角洲地区的上海市,同为海岛的福建海坛岛、浙江嵊泗列岛,分别计算其与长山群岛藓类植物的相似性系数.相似性系数越大,表明物种起源等相似程度越高(表3).
由表3可知:长山群岛与辽宁省沈阳市和仙人洞藓类植物的种属相似性系数较高,分别为25.5%和24.8%,而与上海市的种属相似性系数仅为9.5%.虽然长山群岛与福建海坛岛和浙江嵊泗列岛距离较远,但藓类植物的相似性系数也较高,分别为23.9%和28.5%.
长山群岛与沈阳市和仙人洞自然保护区,均位于辽宁省,地理位置相近,距离较近地区的种属相似性系数相对较高.而长山群岛距离位于长三角地区的上海市较远,因此种属相似性系数较低.同时由于本次在长山群岛调查的范围相对较小,且标本种类较少,辽宁省3个地区之间的共同属和种的数量少,导致了相似性系数不高.还可能因为苔藓植物形体矮小,局部环境的变化可能导致其物种组成上的差异.
海坛岛地处中国东南沿海,岛上的海蚀地貌非常典型.该地区分无人岛和居民岛屿,无人岛上苔藓植物分布主要受森林覆盖率和裸岩面积的影响,居民岛屿上苔藓植物分布除了受自然环境的影响外,还与人为干扰带来的复杂小生境有关.嵊泗列岛位于东亚大陆的东南侧,太平洋的西北部,气候受海陆的共同影响,海岛降水量要少于同纬度的沿海和内陆,降水时段比较集中.沿海岛屿苔藓植物物种丰富与其复杂多样的自然环境有关,并且沿海岛屿具有典型的海蚀地貌,这可能是长山群岛与海坛岛、嵊泗列岛的藓类植物种属相似性系数较高的原因.
3 结 论
本研究对长山群岛(大长山岛、小长山岛和广鹿岛)的藓类植物进行了调查,采集标本259余份,经鉴定共发现藓类植物18科36属96种,其中丛藓科植物占较大比例.长山群岛藓类植物生境是以土生藓类为主,树生藓类几乎没有.由于采样季节的限制,实际采集标本量较少,调查范围不够广泛,导致相关的数据分析不够充分.后续计划在本研究的基础上,进一步扩大调查范围,对长山群岛的苔藓植物进行更加全面的采集和研究,尤其是苔类植物,以期得到长山群岛完整的苔藓植物名录,充分反映长山群岛的苔藓植物多样性.
区系地理成分分析发现,长山群岛的藓类植物以北温带和东亚成分为主,与该区域地处我国东北地区和北温带气候区相一致.另外,通过与其他地区藓类植物种属相似性的比较,发现长山群岛藓类植物与距离较近的辽宁省沈阳市和仙人洞的相似性系数明显高于相距较远的上海市,这符合距离越近物种相似度越高的规律;而与同为海岛的海坛岛和嵊泗列岛的种属相似性系数也较高.由于苔藓植物个体矮小,微生境的变化会导致其物种组成和多样性出现差异,岛屿这样的特殊环境有可能造成物种的相似性较高。本研究对辽宁省大连市长山群岛的藓类植物进行了初步调查,为海岛的苔藓植物多样性提供了基础资料.
参考文献:
[1] CAO T, GAO Q, FU X, et al. Diversity of bryophytes and their conservation [J]. Chinese Journal of Ecology,1997,16(2):47-52.
[2] CHEN Y. Studies on distribution pattern of bryophytes and its relationships with the environments in Shanghai city, China [D]. Shanghai: Shanghai Normal University,2005.
[3] WANG J, CAO T, WANG M, et al. Biodiversity and it’s characteristics of bryophytes in the Suzhou gardens [J]. Acta Botanica Boreali-Occidentalia Sinica,2007,27(6):1239-1246
[4] LUAN W X, WANG H Z. Study of geographical foundation and difference factor on regional development of Changshan islands [J]. Science Geographica Sinica,2005,10(25):124-132.
[5] GAO Q, WU Y H. Flora Bryophytorum Sinicorum, Vol. 10 [M]. Beijing: Science Press,2008.
[6] GAO Q. Moss flora of northeast China [M]. Beijing: Science Press,1977.
[7] GAO Q. Flora Bryophytorum Sinicorum, Vol. 2 [M]. Beijing: Science Press,1996.
[8] GAO Q. Flora Bryophytorum Sinicorum, Vol. 9 [M]. Beijing: Science Press,2003.
[9] GAO Q. Flora Bryophytorum Sinicorum, Vol. 1 [M]. Beijing: Science Press,1994.
[10] HU R L, WANG Y F. Flora Bryophytorum Sinicorum, Vol. 7 [M]. Beijing: Science Press,2005.
[11] HU R L. Bryology [M]. Beijing: Higher Education Press,1987.
[12] LI X J. Flora Bryophytorum Sinicorum, Vol. 3 [M]. Beijing: Science Press,2000.
[13] LI X J. Flora Bryophytorum Sinicorum,Vol 4 [M]. Beijing: Science Press,2006.
[14] WANG H S. Floristic Geography [M]. Beijing: Science Press,1992.
[15] WU P C, JIA Y. Flora Bryophytorum Sinicorum, Vol. 8 [M]. Beijing: Science Press,2004.
[16] WU P C, JIA Y. Flora Bryophytorum Sinicorum, Vol. 5 [M]. Beijing: Science Press,2004.
[17] WU P C, JIA Y. Flora Bryophytorum Sinicorum, Vol. 6 [M]. Beijing: Science Press,2011.
[18] ZHANG Y L. Cofficient of similarity: an important parameter in floristic geography [J]. Arid Zone Research,1998,17(4):429-434.
[19] ZHANG Y H. The sustainable development capacity study of Chang-Shan Archipelago’s survival and environmental supporting system [D]. Dalian: Liaoning Normal University,2005.
[20] SUO A N, SUN Y G, LI B Y, et al. Vegetation landscape health assessment in Changshan Archipelago, North Yellow Sea [J]. Chinese Journal of Applied Ecology,2015,26(4):1034-1040.
[21] GUO S L, CAO T. Study on community distribution patterns of epiphytic bryophytes in forest ecosystems in Changbai Mountain [J]. Acta Phytoecologica Sinica,2000(4):442-450.
[22] LI G Y. Relationship between environment and morphology and genetic diversity of bryophytes [D]. Chengdu: Sichun University,2004.
[23] WANG J, CAO T, WANG M, et al. Floristic analysis of bryophytes from urban area of Suzhou [J]. Journal of Shanghai Normal University (Natural Sciences),2008,37(1):105-110.
[24] STRINGER S M H L. A quantitative study of corticolous bryophytes in the vicinity of Winnipeg, Manitoba [J]. The Bryologist,1974,77(4):551-560.
[25] ZHAO D X, WANG C, HAO Z Z. Epiphytic bryophyte diversity and its influencing factors [J]. Acta Ecologica Sinica. 2020,40(8):2523-2532.
[26] SLACK N G. Host specificity of bryophytic epiphytes in eastern North America (Geography and Ecology of Bryophytes) [J]. Journal of the Hattori Botanical Laboratory,1976,41:107-132.
[27] WU Z Y, WANG H S. Natural geography of China [M]. Beijing: Science Press,1983.
[28] WU Z Y, ZHOU Z K, LI D Z, et al. The areal-types of the world families of seed plants [J]. Acta Botanica Yunnanica,2003,25(3):245-257.
[29] WU Z Y. The areal-types of Chinese genera of seed plants [J]. Acta Botanica Yunnanica,1991,13(4):1-139.
[30] CHEN F. Landform and evolution of Haitan Island, Fujian Province [J]. Marine Science Bulletin,1994(6):62-68.
[31] LI Y Y. Diversity and biogeography of bryophytes in Haitan Island and its adjacent islands, Fujian Province [D]. Shanghai: Shanghai Normal University,2020.
[32] CAI J R. Studies on flora and geographical distribution of bryophytes in Shengsi Archipelago, Zhoushan, Zhejiang [D]. Shanghai: Shanghai Normal University,2017.
(責任编辑:顾浩然,包震宇)
收稿日期: 2021-07-07
基金项目: 国家自然科学基金(31570208);上海市自然科学基金(21ZR1447400);上海植物种质资源工程技术研究中心项目(S17DZ2252700)
作者简介: 宋玉玲(1997—),女,硕士研究生,主要从事苔藓植物系统分类方面的研究. E-mail: 857544750@qq.com
* 通信作者: 于 晶(1972—),女,教授,主要从事苔藓植物系统分类与生物地理学方面的研究. E-mail: yujing@shnu.edu.cn
引用格式: 宋玉玲, 李丹丹, 于晶. 长山群岛藓类植物多样性研究 [J]. 上海师范大学学报(自然科学版),2022,51(1):94‒100.
SONG Y L, LI D D, YU J. Study on the diversity of mosses on Changshan Islands [J]. Journal of Shanghai Normal University(Natural Sciences),2022,51(1):94‒100.
