心肌致密化不全的分子遗传学机制研究进展
2022-03-08张琪洪楠超王静陈丰原孙锟于昱
张琪 洪楠超 王静 陈丰原 孙锟 于昱
200092 上海交通大学医学院附属新华医院,心血管发育与再生医学研究所
心肌致密化不全(noncompaction of ventricular myocardium,NVM)又称左心室致密化不全(left ventricular noncompaction,LVNC)或非致密性心肌病(noncompaction cardiomyopathy,NCCM)[1-3]。以异常粗大的心室肌小梁和深陷的小梁间隐窝为形态学特征,病变主要累及左心室。可孤立发生,也可伴有其他先天性心脏结构异常,如房室间隔缺损、动脉导管未闭、Ebstein畸形等。最早于1926年由Grant首次报道,最初被称作“海绵状心肌”。1990年由Chin等正式命名为孤立性左室心肌致密化不全。近年来,由于NVM在人群中具有高发病率和高死亡率而受到越来越多的关注,但其病因、发病机制、诊断及管理等方面仍存在诸多争议[4-6]。
随着分子遗传学研究手段的发展,人们对NVM的遗传学背景有了一定了解。家系研究显示,其具有明显家族聚集性,可为常染色体显性遗传(如编码肌节蛋白基因突变)、X连锁隐性遗传(如Barth综合征)、常染色体隐性遗传和母系遗传(由线粒体DNA突变导致)。在个别散发性病例中,遗传可能由新发突变(De novo mutation)导致。此外,NVM还与其他先天性代谢性异常疾病(包括1B型糖原贮积病、丙二酰辅酶A脱羧酶缺乏症和钴胺素C缺乏症)、染色体异常(特纳综合征、21/18/13三体综合征、22q11缺失和1p36缺失等)、神经肌肉性疾病(Duchenne、Becker综合征)和其他遗传综合征(Soto、Marfan和RASopathies综合征)相关[7]。对其遗传背景进行深入了解,从分子遗传学角度解释NVM的发病机制,将是未来全面揭示NVM病因学的关键路径。
1 胚胎发生假说
为了解释NVM的发病异质性,Finsterer[8]将NVM分成原发NVM(primary NVM)和继发NVM(secondary NVM)。原发NVM是指能找到遗传缺陷的类型,该遗传缺陷导致NVM的疾病表型,比如NOTCH信号通路相关基因发生缺陷,干扰心室肌发育导致NVM。继发NVM即没有遗传缺陷,但可以通过缺氧、炎症或以血流动力学改变来解释异常肌小梁形成的NVM类型。对于原发性NVM而言,胚胎发生假说是早期较为公认的致病原因。该假说认为,NVM是胚胎发育阶段心室肌小梁重构(致密化)过程失败所致。心肌细胞在多种遗传因素调控下从中胚层原基分化为心肌。在人类胚胎第3个月末,可以明显观察到肌小梁以许多长突起形式发源于心内膜层。在冠状动脉建立之前,小梁的存在扩大了用于心肌氧供和物质交换的表面积。随着心脏持续的发育和循环系统压力的增加,小梁形态发生重构,呈现心肌纤维螺旋式致密化。在妊娠期第4个月,形成紧实的心肌并构成了心室腔。若此过程受损,心室肌则无法发挥收缩功能进而继发心肌功能障碍,这也是NVM发生的关键环节[9]。与此同时,人们还提出另一种假设,认为NVM的异常小梁是由心室细胞异常增殖引起的,具体机制尚不清楚[10-11]。Choquet等[12]在小梁发育过程的关键阶段删除编码转录因子Nkx2.5的基因,小鼠心脏表现出异常小梁结构和菲薄致密心肌层,而该表型在发育晚期敲除个体中则并未发现,这一结果支持了NVM起源于早期心脏发生缺陷的观点。
对于继发性NVM,非胚胎发生假说(缺氧、炎症、血流动力学改变)似乎更为适用。近年来在运动员[13]、妊娠期妇女[14]、镰状细胞性贫血症患者[15]、某些神经肌肉性疾病患者以及慢性肾衰竭患者中观察到后天获得且部分可逆的NVM病例。这些病例促使胚胎发生假说和获得性非胚胎发生假说的双重假设模式被提出,还引发人们对表观遗传学影响的思考[16]。
2 分子遗传学机制
NVM的分子遗传致病机制是复杂的。NVM患者常被发现具有其他心肌病亚型[扩张型心肌病(dilated cardiomyopathy,DCM)、肥厚型心肌病(hypertrophic cardiomyopathy,HCM)]家族史,表明NVM可能与其他心肌病具有共同的遗传机制[17]。因此,与其他心肌病类似,NVM也具有“共同致病途径”[18]。不同的是,NVM的致病途径不是单一的,在大多数情况下,NVM的发生可能是主要途径(例如肌节基因)和干扰发育过程的发育途径(例如NOTCH信号通路)的共同结果[19]。且临床证据表明,发生多基因突变的病例更为普遍,致病基因之间可能具有协同性。几个常见且重要的候选致病基因见表1。

表1 NVM致病途径分类的代表性候选基因
2.1 主要致病机制
主要途径是指涉及编码心肌结构蛋白成分等基础结构的基因发生致病性改变,包括编码肌节、细胞骨架蛋白、离子通道蛋白等。
2.1.1 肌节蛋白基因 肌节基因的突变是NVM最常见的致病原因,在最新涵盖国内外、具有最大病例数(7 598例)的分析报告中,肌节基因突变率最高。以TNN为首,其突变率为11%;MYH7次之(9%);MYBPC3紧随其后(7%)[20]。一些肌节基因如TPM1、TNNC1或ACTC1突变虽不常见却导致较差预后[21]。Takasaki等[21]通过人诱导多能干细胞证明了肌节基因在NVM发生中的“遗传触发”角色,影响了心脏发育相关的众多基因的表达,从而导致心肌细胞增殖和分化过程失衡。然而,多数肌节基因缺乏特异性,其同时参与其他心脏疾病的发生,例如编码肌球蛋白-β的MYH7基因的突变,还与DCM有关[22];而编码肌球蛋白结合蛋白C的MYBPC3突变,也与HCM相关[23]。新近报道的致病候选基因SORBS2似乎更具特异性。该基因仅在NVM心肌中高表达,在HCM或致心律失常性右室心肌病(arrhythmogenic right ventricular cardiomyopathy,ARVC)心肌中表达均完全正常。上调的SORBS2通过介导β-微管蛋白形成、聚合、致密化异常,Junctophilin 2(JP2)异位表达和兴奋收缩偶联(E-C偶联)功能障碍,最终导致NVM小鼠心肌细胞收缩功能障碍和心力衰竭。该研究证实了SORBS2增加NVM患者发生心力衰竭的概率,对制订NVM靶向治疗策略以延缓心脏重构和心力衰竭的进展具有参考价值[24]。
2.1.2 细胞骨架蛋白基因 7 598例NVM报告中,细胞骨架蛋白候选基因也较为常见,以LDB3(3%)、LMNA(2%)和DTNA(1%)最为多见。LDB3也称为ZASP或Cypher,可编码心肌和骨骼肌细胞质中的特异性Z带蛋白,与肌动蛋白共定位。该基因通常导致NVM合并DCM、HCM或ARVC的复杂表型[25-26]。而编码核膜蛋白基因的LMNA,不仅与NVM相关,还与其他心肌疾病以及心脏传导系统疾病相关联。此外,α-肌营养不良蛋白DTNA也是NVM的重要致病候选基因,其主要功能是结合肌营养不良蛋白及肌营养不良蛋白相关糖蛋白复合物,在肌肉收缩和松弛过程中起到维持细胞膜稳定性的作用,其异常表达可导致心肌收缩舒张功能受限,心脏负荷不良,最终诱发NVM。已报道的NVM细胞骨架致病候选基因也可导致其他类型的心肌病,因而部分学者认为尚不能确定NVM为一种独立的心肌病还是其他心肌病所共有的形态学特征[6]。
2.1.3 离子通道蛋白基因 离子通道病相关基因也称作心律失常基因,是一组与离子通道转运相关的基因。编码电压-门控钠通道的SCN5A基因突变最具代表性,常表现为长QT间期综合征等一系列心律失常[27]。近年来NVM的报道更多集中在基因型与不良预后的相关性上。心律失常连同心力衰竭和血栓栓塞事件是NVM的三大致病性表现,离子通道蛋白与NVM患者的心律异常密切相关,这对伴发心律失常NVM的靶向治疗以及通过基因筛查进行早期预防具有重要指导意义。
2.1.4 线粒体疾病相关基因 Tang等[28]在NVM患者的线粒体基因组序列中发现了ND1基因同一位点Met31的不同突变。由于线粒体复合体对心肌细胞增殖、分化、功能实现的重要性,这种高度保守的残基改变以及随之而来的复合体缺陷可能是NVM的潜在致病机制。而核编码线粒体蛋白基因的典型代表为TAZ基因,该基因也是最早发现也最为重要的NVM候选基因之一。TAZ/G.5编码Tafazzins蛋白,在线粒体内具有酰基转移酶的功能,是组成线粒体内膜的主要成分。1997年,Bleyl等[29]描述了独立发生于Xq28区域的TAZ基因突变可导致孤立性NVM,共有6个表现出X连锁隐性遗传的家庭被纳入分析,这是NVM的第一个遗传学证据。并且,在之后的报道中,人们在该基因中发现了150多个致病突变能够导致NVM,主要为点突变[30]。
2.2 干扰发育过程的致病机制
发育途径主要涉及调控心肌发育的转录因子和信号通路组分异常,发育过程的中断或异常都可能导致NVM的发生。
2.2.1 NOTCH信号通路相关基因 NOTCH是高度保守的信号传导途径,可调节细胞的生长、分化和衰亡[7]。NOTCH信号通路组成成分的表达异常在NVM病例中的发生率极高,包括MIB1、FKBP12等。编码E3泛素连接酶的MIB1是 NOTCH通路中的重要调节组分,可调节Notch配体Delta和Jagged的内吞作用。对小鼠心肌的MIB1或Jagged1进行靶向失活、干扰NOTCH信号传导最终诱发了NVM的发生[31]。FKBP12(也称Fkbp1a)是内皮Notch1受体的活性调节剂,表达于发育阶段的心内膜。心内膜特异性敲除FKBP12诱发了小鼠心肌NVM表型,且检测到N1ICD(Notch1胞内段)活性增强。此外,抑制N1ICD活性可部分逆转FKBP12敲除鼠的NVM表型,FKBP12缺失突变诱发NVM可能与Notch1受体胞内段N1ICD活性增强有关。同时,人们还观察到FKBP12的下游调控因子BMP10的变化,其不仅在FKBP12缺失小鼠中显著上调,在Nkx2.5缺失小鼠和Numb或类似Numb缺失小鼠中也被观察到显著且一致的改变。利用人心钠素(human atrial natriuretic peptide,hANF)启动子诱导小鼠胚胎心脏BMP10的过表达,可显著增加心肌的异常小梁出现[18,32]。有趣的是,由BMP10缺失导致的NVM心室壁中p57kip2上调且异位表达;而在FKBP12缺失导致的NVM心室壁p57kip2水平显著降低。目前仍然认为心肌细胞的增殖水平的改变是NVM发生的重要原因之一。
2.2.2 Wnt/PCP信号通路相关基因 Wnt/PCP信号的关键作用为调控极化组织中细胞的排列和方向,其在胚胎发育和器官发生中起着至关重要的作用[33-34]。科学家对Wnt/PCP信号通路上重要的效应分子Daam1激活子进行基因敲除小鼠模型的构建,不仅观察到伴发右室双出口和室间隔缺损的NVM[35],更重要的是,缺乏Daam1的心脏具有正常的心肌细胞增殖和Bmp10表达,表明细胞增殖水平的改变的确不足以解释NVM的发生[35]。在该研究中,心肌的肌纤维生成、肌节形成、心肌细胞极化、细胞-细胞排列和细胞-细胞连接均发生明显异常,进一步证实除细胞增殖外其他NVM的可能致病机制。截至目前,已发现数个Wnt/PCP信号传导通路的核心组分基因突变能够诱导出NVM表型。
2.2.3 其他调控心室小梁发育的基因 在内皮细胞特异敲除Sema受体Plxnd1基因,不仅验证了Plxnd1的缺失可通过减少细胞外基质蛋白的水解,致使细胞外基质过度沉积,进而诱发NVM的发生,还揭示了Sema3E/PlexinD1信号通路在心肌小梁发育过程中的重要作用[36]。而在针对调节心室发育的ang1-Tie信号通路探索中,发现富集于心内膜的Tie2具有两个互补功能,一是通过促进心内膜细胞增殖和发芽来确保正常小梁,另一方面通过抑制小梁心肌细胞中的RA信号通路来防止过度小梁。该研究发现了一个新的具有潜力的NVM致病基因[37]。此外,其他一些较为罕见的候选基因也陆续被发现,包括维持心脏功能和结构正常所需的关键分子RBM20;调节淋巴细胞早期免疫应答的NF-AT转录因子;神经调节蛋白Nrg;酪氨酸激酶受体ErB2;血管生成素1(Ang-1)等[37-40]。
2.3 染色体缺陷
由染色体缺陷所导致的先天性疾病常常累及心脏,其中以携带lp36微缺失最为多见。在这些主要的染色体缺陷综合征类型中,均可见明显的NVM改变,但具体致病机制尚未可知。
3 基因工程鼠模型
在过去的几年中,基因工程鼠模型被广泛用于NVM的潜在机制研究[18],尤其是针对调控发育的信号网络,以克服难以在人群中溯源胚胎早期心脏发育过程的困难[7]。FKBP12缺失小鼠是第一个心肌致密化不全的特异小鼠模型,因此,也被广泛用于研究调节心室小梁形成和致密化的潜在机制[34]。小梁发育早期敲除Nkx2.5的小鼠模型,不仅提供首个NVM成年小鼠模型,还支持了NVM的胚胎起源[41]。一些在人类中与NVM相关的基因未能在小鼠体内验证和开展研究。例如,人类TNNT2的外显子第96位密码子(从GAG到AAG)错义突变被报道与NVM家族性病例有关,但在小鼠心脏过表达TNNT2却无法诱导NVM表型[42]。除了种属的巨大差异外,还可能是由于遗传变异并非该病的唯一致病因素,修饰基因的调控(表观遗传学)以及环境因素也与之相关;以及过表达的构建系统无法真实模拟体内致病途径,这是未来工作需要突破的地方[16]。
除了前文提到的小鼠模型外,还有许多NVM小鼠模型已被构建应用,例如Jarid2基因敲除小鼠、14-3-3/Ywhae基因敲除小鼠[43-44]、过表达SHP2-Q79R(Noonan综合征)小鼠[45]和过表达βMHC-Met531Arg小鼠[46]等。目前主要的NVM基因工程鼠模型种类,见表2。
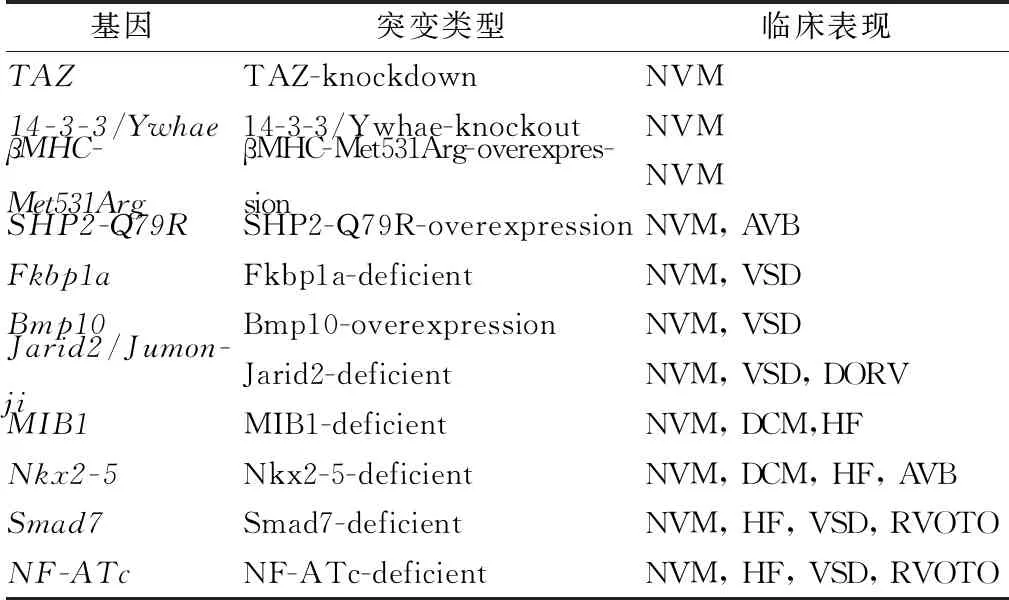
表2 NVM基因工程鼠模型
4 小结
NVM的发病机制涉及分子和细胞水平的多种机制,随着基因测序技术的发展和基因工程鼠模型的运用,人们对NVM分子遗传学致病机制的研究取得了一定进展。目前已报道的NVM致病候选基因集中在心肌结构蛋白成分和调控心脏发育的信号通路上,涉及心室小梁的形成发育过程,心室肌细胞的增殖异常和心肌纤维极化异常。然而,现有研究多基于已知候选基因的筛查,覆盖范围窄且易受其他疾病表型影响而缺乏特异性。除TAZ、FKBPl2等一些通过实验动物验证了基因突变与NVM的相关性外,大部分相关基因信息多来自遗传连锁和突变的分析,缺乏与NVM直接相关的基因型表型实验证据及进一步的功能分析及机制研究。染色体缺陷导致NVM发生的机制也尚存许多知识空白。值得注意的是,在临床病例中,尤其是在高危NVM表型个体中,发生多基因突变的情况更为普遍,是否提示NVM致病候选基因之间可能具有协同作用。未来研究工作中,应更多关注于致病候选基因之间的相互影响,最终为解决NVM医学难题提供新策略。
利益冲突:无
