广西黑叶猴栖息地景观格局破碎化分析及其对种群的影响
2022-03-07管超毅黄乘明周岐海
管超毅,陈 智,黄乘明, 周岐海,*
1 广西师范大学,珍稀濒危动植物生态与环境保护教育部重点实验室,广西珍稀濒危动物生态学重点实验室, 桂林 541004 2 中国科学院动物研究所,北京 100101
生境破碎化一直被认为是造成生物多样性丧失的主要原因之一,它通过减少生物量和改变养分循环的方式,使全球27%的哺乳动物物种面临灭绝的威胁[1—2]。生境破碎化不仅影响栖息地数量和构造,而且还导致适宜生境不再连续,物种的迁移、扩散和交流也会因此受到限制[1—2]。当前,工业建设和农业扩张等人类活动已经改变了50%以上的原始陆地覆盖,大部分野生动物栖息地破碎化日趋严重[2]。对于灵长类动物和其他大型哺乳动物而言,它们更依赖于完整森林生态系统来获取生存所需要的食物和安全场所,栖息地破碎化更容易对这些物种的健康产生一系列连锁反应,并增加种群局部灭绝的可能性[3]。
从生物多样性保护实践来看,物种的保护既要考虑目标物种本身,还要考虑它所在的生态系统及有关生态过程[4]。因此,了解景观空间格局如斑块数量,边缘长度,破碎程度等对于物种及其生态过程的影响具有重要意义。景观空间格局即景观格局,是大小、形状、属性各异的景观要素在空间上的分布和配置,它表现为不同类型的景观斑块在空间上随机、均匀或聚集分布[5]。景观格局分析是研究栖息地破碎化对生物多样性响应的重要方法之一[6],能够有效反映栖息地的环境质量[7]、栖息地适宜性的动态变化[8]、种群对栖息地破碎化的响应[9]。目前,国内外众多学者利用景观空间格局分析栖息地破碎化对物种的影响,如川金丝猴(Rhinopithecusroxellana)[10]、绢毛猴(Saguinusbicolor)[11]、黑白仰鼻猴(R.bieti)[12]。
黑叶猴(Trachypithecusfrancoisi) 属灵长目(Primates) 、猴科(Cercopithecidae) 、疣猴亚科(Colobinae)、乌叶猴属(Trachypithecus),为我国Ⅰ级重点保护野生动物,在中国脊椎动物红色名录中列为濒危动物(endangered,EN),其主要分布在中国西南部和越南北部的热带和亚热带喀斯特地区[13]。黑叶猴曾广泛分布于我国的广西、贵州、重庆。然而,黑叶猴的生存和繁衍受到了前所未有的挑战,非法捕杀和森林砍伐等人类活动导致其种群数量和栖息地面积呈现断崖式下降。尤其在广西,20世纪90年代中期,黑叶猴种群数量从20世纪80年代的4500—5000只减少至90年代的2000—2500只,其适宜栖息地零散分布在15个自然保护区中。21世纪初,广西黑叶猴种群数量急剧减少至44群307只[14—15]。目前学者已开展栖息地破碎化对黑叶猴行为生态的影响,主要集中在对觅食活动[16—17]、生境利用[18]等方面的影响,而针对生境破碎化评价以及破碎化对黑叶猴种群数量的影响等方面的研究还相对欠缺。因此,本文以广西黑叶猴为研究对象,结合景观格局与猴群数量关系进行深入分析,旨在探讨以下两个问题: (1)对黑叶猴栖息地的破碎化程度进行量化评估; (2)生境破碎化对黑叶猴种群数量的影响程度。以期为这一珍稀濒危物种的保护及自然保护区的管理提供科学依据。
1 研究区域与对象
2003—2005年,Li等[17]基于前期访问调查的结果,采用样线法结合小区蹲点调查法开展广西黑叶猴数量调查。本研究基于Li等[17]的调查结果,选取9个黑叶猴主要分布区作为研究区域(图1),包括龙州县、扶绥县、隆安县(群吁)、德保县、靖西县(岳圩、南坡和新兴)、隆林县。研究区属于亚热带季风气候,其热量充足,降雨充沛,雨热同期,最冷月平均气温在0℃以上,最热月平均气温大于22℃[19—20]。大部分地区由石灰岩组成的山地和丘陵,地貌主要为峰林谷地和峰丛洼地,植被以亚热带常绿阔叶林为主[19—20]。除广西弄岗国家级自然保护区外,大部分黑叶猴分布区人为干扰严重,许多山间平地被开垦为农业用地[15,19]。
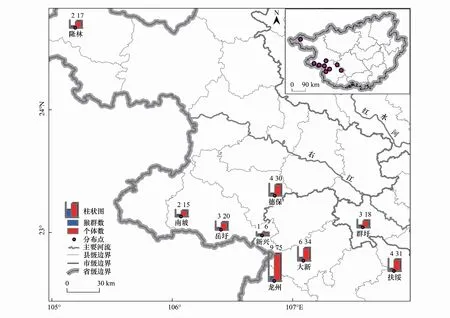
图1 广西黑叶猴数量与分布位置Fig.1 Number and distribution of François′ langurs in Guangxi
黑叶猴分布区边界的划定主要依据以下原则:(1)如果黑叶猴分布在保护区内,依据保护区边界进行划定;(2)如果遇到黑叶猴无法逾越的地理屏障(如河流),依据这些地理屏障进行划定;(3)依据喀斯特石山的走向及其边界进行划定。
2研究方法2.1景观分类影像分类
基于1999—2001年的Landsat TM影像,根据黑叶猴的生态习性,以及考虑不同用地类型对其适宜程度,利用ERDAS IMAGINE软件将研究区域分为7个基本景观类型:林地、灌草丛、裸地、裸岩、农田、居民点、水体(包括泡沼、水湾、水库等)。
2.2 景观分析
结合景观指数的生态学含义[8]、黑叶猴的生境特征以及参考相关研究[21—22]选取景观指数,选用15个景观变量,其中包括6个景观水平和9个类型水平(表1)。基于此,利用景观分析软件FRAGSTATS 软件对研究区景观格局进行分析,探讨黑叶猴栖息地破碎化现状
式中:TA为景观总面积 Total landscape area;MPS为斑块平均大小 Mean patch size;MNFD为平均斑块分维指数 Mean patch fractal dimension;CONTAG为蔓延度指数 Contagion index;SHDI为Shannon多样性指数 Shannon's diversity index;FN为景观块数破碎化指数 Fragmentation indices of patch number;HDI为人为干扰指数 Human disturbance index;CA为景观面积 Landscape area;PLAND为景观面积占比 Percentage of Landscape
2.3 统计分析
本研究使用R 3.6.1软件计算景观格局(不包括景观总面积)与黑叶猴个体数量的 Pearson 相关系数及其显著性检验,并利用多元逐步回归建模确定引起黑叶猴个体数量差异的关键因子,此外所有检验的差异显著水平设定为P<0.05。
3 研究结果
3.1 景观分析
在景观水平上(表2),平均斑块大小、平均斑块分维指数和蔓延度指数等格局指数的平均值分别为18.73、1.3499和76.35。龙州的平均斑块大小、均斑块分维指数和蔓延度指数最大,分别为32.18、1.3727和91.27;德保的平均斑块大小指数最小,为10.26;而南坡的平均斑块分维指数和蔓延度指数最小,分别为1.3319和71.11。Shannon多样性指数和景观块数破碎化指数等格局指数的平均值分别为0.83和0.00523(表2)。群吁的Shannon多样性指数最大,为0.98;德保的景观块数破碎化指数最大,为0.0087;而龙州的Shannon多样性指数和景观块数破碎化指数等格局指数最小,分别为0.27和0.0028。人为干扰指数的平均值为0.29(表2);其中龙州的人为干扰指数最小,为0.01;而扶绥的人为干扰指数最大,为0.65。而在各个分布区中,龙州的面积最大,其面积为6821.39 hm2;而新兴的面积最小,其面积为1054.61 hm2;面积大小排序为龙州>大新>扶绥>岳圩>隆林>南坡>德保>群吁>新兴。
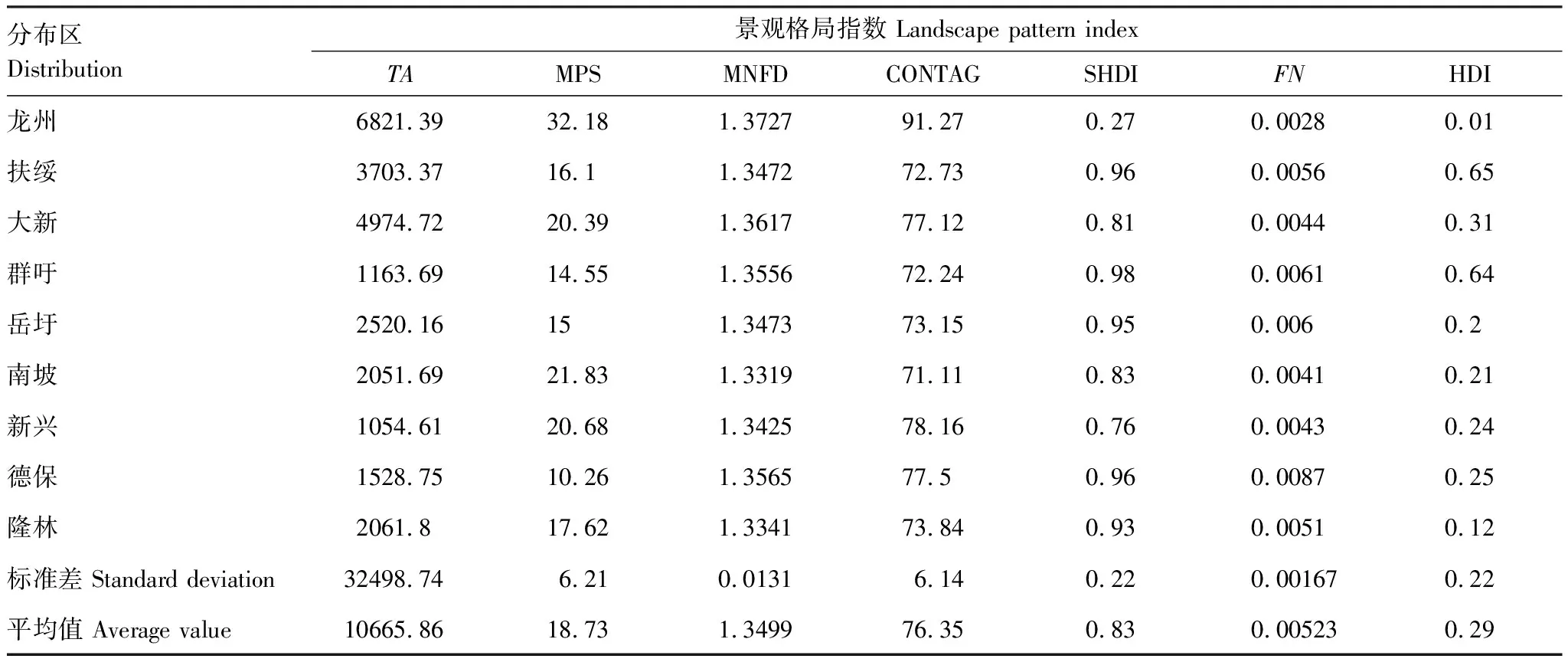
表2 广西黑叶猴栖息地景观水平上的景观格局分析
在类型水平上(表3),自然景观是主要的优势类型,其中林地景观具有绝对的优势,其面积占比的平均值为71.50%;人为景观也有较高的比例,其中耕地面积占比的平均值为20.25。黑叶猴的各分布区中,龙州地区的林地聚集,林地景观面积为6423.32 hm2,林地斑块平均大小为2141.11 hm2,拥有最大的林地景观。而群吁的林地景观面积仅为641.12 hm2,林地斑块平均大小仅为64.11 hm2,是最小的林地景观。而扶绥地区耕地密布,耕地面积为1438.19 hm2,其面积占比为38.83%,是最大的耕地景观。人为景观影响最小的地区是龙州,其中耕地景观面积仅为61.41 hm2,其面积占比为0.9%。
3.2 景观破碎化对黑叶猴种群的影响
从景观水平的景观格局与黑叶猴个体数量的相关分析表明(图2):蔓延度指数和平均斑块分维指数与黑叶猴个体数量呈现显著正相关;Shannon多样性指数与黑叶猴个体数量呈现显著负相关。斑块平均大小与黑叶猴个体数量呈现正相关,相关性接近于统计学显著水平,而黑叶猴个体数量与景观破碎化指数和人为干扰指数无显著相关性。可见黑叶猴对景观连通程度的好坏(蔓延度指数)有一定的要求。另外平均斑块分维指数和景观破碎化指数均反映了景观的破碎程度,但是它们对黑叶猴的响应并不一致。

表3 广西黑叶猴栖息地类型水平的分析结果

图2 景观水平的景观格局指数与黑叶猴数量的相关性分析Fig.2 Correlation analysis between landscape pattern and the population of François′ langurs at the landscape scale
从类型水平的景观格局与黑叶猴个体数量的相关分析表明(图3):林地和裸岩等自然景观对黑叶猴有促进作用。其中林地景观面积、林地平均斑块大小、裸岩面积和裸岩面积占比与黑叶猴个数量呈现显著正相关。而黑叶猴个体数量与耕地面积、耕地占比、居民地面积、居民地占比均无显著相关性。可知裸岩的景观格局与黑叶猴个体数量的相关性最强,裸岩景观有利于黑叶猴的生存。
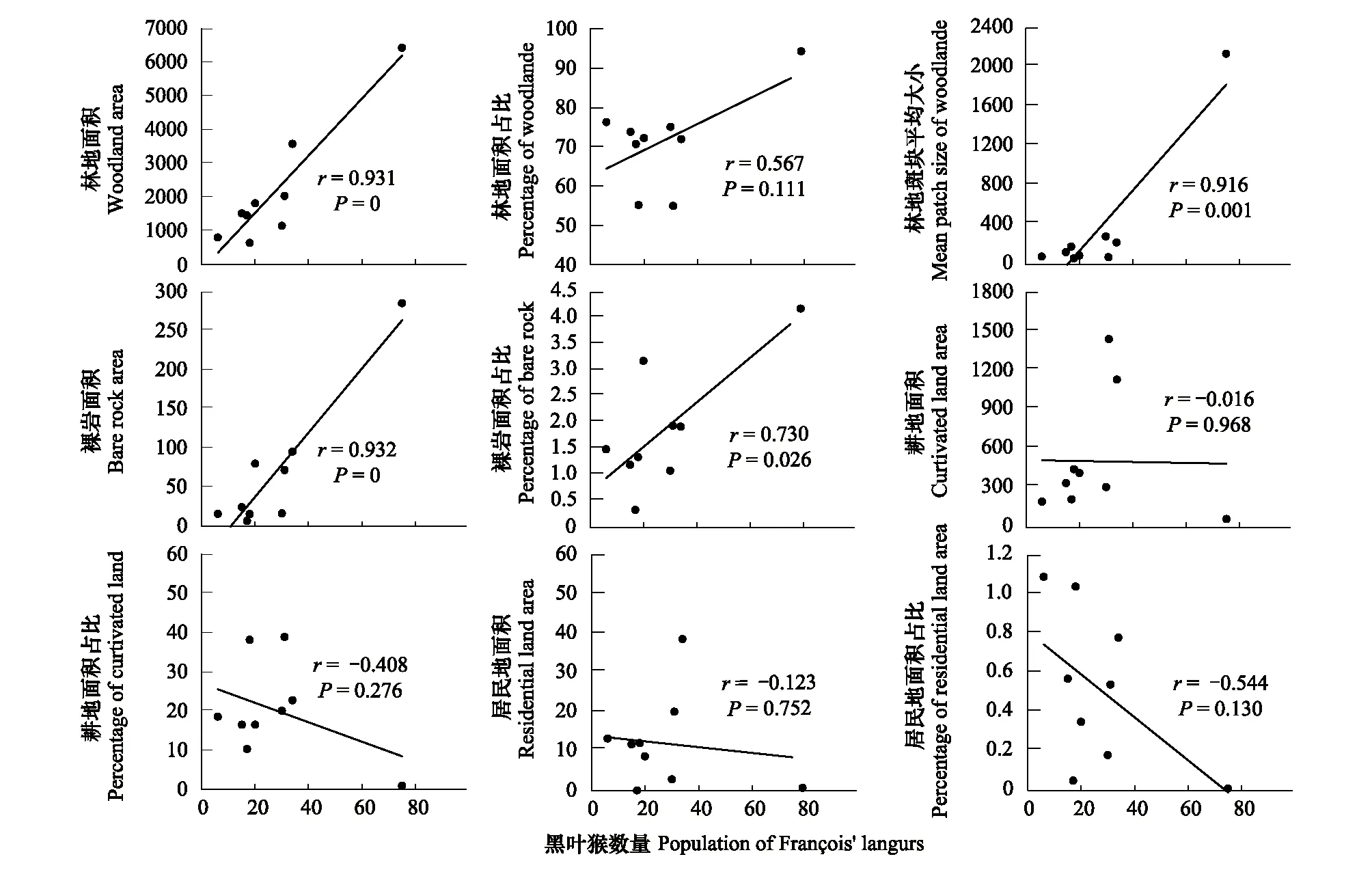
图3 类型水平的景观格局指数与黑叶猴数量的相关性分析Fig.3 Correlation analysis between landscape pattern and the population of François′ langurs at the Class level
将得到了5个景观水平的景观格局变量和9个类型水平的景观格局变量与黑叶猴个体数量(K)进行逐步回归分析(表4)经过模型自动识别,最终林地面积、居民地占比和平均斑块分维指在模型中,得到了“最优”模型:K=-787.041+ 0.006×林地景观面积-12.284×居民地占比+598.354×平均斑块分维指数 。该模型R方值为0.957,解释性优秀,拟合度效果较好。且模型通过F检验(F=37.488,P=0.001),说明模型有效。林地景观面积的回归系数值为0.006(t=3.620,P=0.015),意味着林地景观面积会对黑叶猴个体数量产生显著的正向影响关系。居民地面积占比的回归系数值为-12.284(t=-2.266,P=0.073),意味着居民面积占比并不会对黑叶猴个体数量产生影响关系。平均斑块分维指的回归系数值为598.354(t=2.914,P=0.033),意味着平均斑块分维指(MNFD)会对黑叶猴个体数量产生显著的正向影响关系。
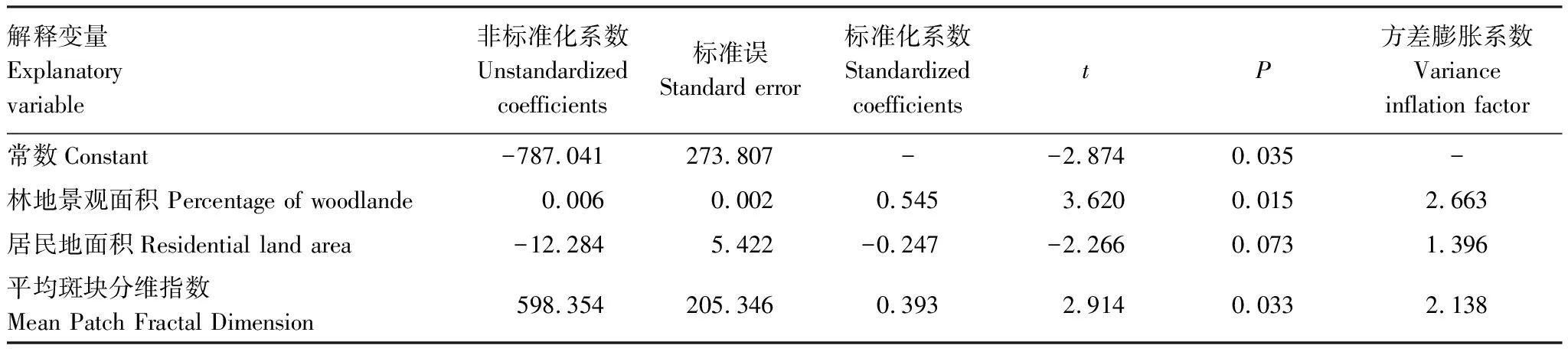
表4 景观格局与黑叶猴种群数量的逐步回归分析
分析结果表明:在景观水平上,平均斑块分维指数是主要造成黑叶猴个体数量差异的主要原因。在类型水平上,林地面积是是主要造成黑叶猴个体数量差异的关键变量。从标准化系数Beta的绝对值来看,林地面积>平均斑块分维指>居民地占比。可见林地面积和平均斑块分维指对黑叶猴个体数量的影响尤为突出,而林地面积>的影响力最强。
4 讨论
本研究对广西黑叶猴分布区进行景观格局分析,探究广西黑叶猴分布区的栖息地破碎化现状。在景观水平上,除龙州外,黑叶猴栖息地呈现破碎度严峻、丰富度多元化、斑块形状复杂化、斑块团聚程度较弱且分散化的现象。研究表明,栖息地破碎化主要来自于自然和社会的双重影响[22]。黑叶猴栖息地岩溶作用强烈,造就了地表破碎化;在地表破碎化的基础上人为干扰又造成的原生植被破碎化。随着社会发展、人口增加,当地居民需要更多的耕地,而贫瘠的喀斯特地区土地难以提供更高的生产力,当地居民只好通过开垦山脊和坡地来扩大耕地面积以维持平衡[17,23]。不合理的森林砍伐和农业开垦加剧了的黑叶猴栖息地破碎化,许多栖息地被大量的农耕地分隔开,其中扶绥地区尤为明显[15—16]。在斑块类型水平上,黑叶猴在广西主要分布区的景观配置以林地景观占据主要优势,但各分布区的林地面积占比差异明显。这种差异主要是取决于人为干扰的强弱。除了岳圩和新兴以外,其他分布区均在自然保护区内,由于保护区的保护,其植被条件较好,灌草丛景观和人为干扰景观类型较少。然而很多保护区的并没有得到有效的管理,使得周边居民的生产活动尤其是资源利用、种养殖等经济行为对森林景观产生了不同程度的人为干扰[23—24]。例如在南坡片区,虽然该地分布于南坡乡底定省级自然保护区附近,但保护区成立时间较晚,农户的保护意识依然薄弱,由于不合理耕作,大量的土地被荒废。而在岳圩和新兴等地由于地形复杂、山高谷深使得人为干扰活动较弱,即使不是在保护区内也能有很大的林地面积占比。
景观格局与黑叶猴数量的相关性研究中发现:在景观水平上,Shannon多样性指数对黑叶猴个体数量呈现负向效应,该结论表明栖息地的破碎化不利于黑叶猴的生存发展。研究表明,栖息地破碎化会沿着生境斑块的边界产生有害的边缘效应,随着生态条件的变化动物运动和基因流动将会受到限制,最后导致当地物种的减少[1,10,25]。而蔓延度指数和平均斑块分维指数与黑叶猴个体数量呈现显著正相关,结果表明保持良好的景观连通性对黑叶猴的生存和繁衍具有重要意义。对于栖息地破碎化加剧的现状,许多国内外研究表明,提高景观的连通性有利于促进种群的迁移扩散、提高种群延续的可能,以避免种群的灭绝[25]。例如Macdonald等利用GIS的分析框架为马来云豹(Neofelisdiardi)构建合理的生态连通网络,以减弱栖息地破碎化的影响[26]。Liu等利用最小成本路径优化了川金丝猴的适宜栖息地的潜在连通性,以改善人为干扰和破碎化景观对物种的影响[10]。然而,黑叶猴个体数量与景观破碎化指数并不存在显著相关性。生境破碎化是一个复杂过程,具有高度可变的影响,不同物种对生境破碎化的响应并不一致[22,27]。Galán-Acedo等[27]总结分析景观结构对灵长类动物的影响,发现对其影响最大的景观组成(如森林覆盖度和景观质量指数),而不是景观配置(如破碎化指数)。因此,有必要从景观组成和景观配置全面评估生境破碎化对黑叶猴的影响[27—28]。此外,黑叶猴个体数量与人为干扰指数并不存在显著相关性,这可能与黑叶猴种群密度较低有关,目前的栖息地仍能满足其生存的需求 。
本研究发现,林地景观面积和林地平均斑块大小对黑叶猴个体数量产生显著的正效应。黑叶猴的食物来源主要是乔木和藤本以及少量的草本植物,宽阔林地能够为黑叶猴提供大量的隐蔽场所和食物选择[29]。林地斑块大小直接影响了食物的可获得性和家域范围,较小的林地斑块可能造成食物资源的高度分散,黑叶猴必须花费更多的时间和能量去寻找更多食物,以达到能量平衡[30]。Arroyo-Rodríguez等分别对吼猴的选择偏好研究也发现面积较大森林对猴群更加有吸引力,较大栖息地面积会为动物捕食、栖息和生长繁殖提供更大的安全场所[31]。灵长类动物依赖于具有完整的生物多样性的森林景观来获取营养均衡的饮食,非生境景观的面积占比过大会改变了原有的营养均衡从而导致当地物种灭绝和生物数量的减少[32]。此外,还发现裸岩面积和裸岩面积占比与黑叶猴数量产生显著正相关。黑叶猴是一种石山灵长类,主要栖息在陡峭的石山中,陡峭的石山可提供大量的遮盖物,以及更容易发现捕食者[33]。
本研究通景观格局指数与黑叶猴个体数进行多元逐步回归建模,分析景观格局对黑叶猴数量的影响。其结果表明林地面积和平均斑块分维指数都对研究区的黑叶猴种群产生影响,但是林地面积对黑叶猴的影响更大。在破碎化景观中,黑叶猴的丰度主要是受到林地的影响,同时景观空间格局需要有良好的连通性以保证黑叶猴能顺畅地在斑块间移动。研究栖息地破碎化和森林退化对灵长类的影响,即要考虑景观属性也需要考虑景观空间格局[28]。在森林地带中,林地面积限制了猴群的最大容纳量,因此,迁徙到新的栖息地是灵长类动物保持种群长久延续的有效策略,个体的迁移能防止在食物不足时导致的种群灭绝以及缓解种群内部的竞争压力[28,34]。正如Galán-Acedo等研究发现,微小森林中的蜘蛛猿(Atelesgeoffroyi)可能会被迫更频繁地使用基质进行觅食或迁徙[34]。迁徙后的灵长类动物如果能从邻近的斑块中补充食物,它们就能在资源相对匮乏的生境中持续生存[28,35]。Asensio等发现吼猴(AlouattapalliataMexicana)会穿过阻碍斑块到邻近的景观元素(孤立的树木、被隔离的山峰)中补充饮食,这些景观元素被视为其家域的潜在延伸[36]。因此对于生态保护的相关从事人员,单单保护森林面积的范围是远远不够的,还应该考虑提高生境景观的连通性以降低森林破碎化对物种的影响。
由于广西黑叶猴各分布区的景观格局配置有明显差异,因此在进行保护规划时,应该有不同的侧重点。如果当地的栖息生境破碎程度严重,那么管理规划应集中在景观廊道的建立上以提高自然景观的连通性。但如果是分布区内森林覆盖率严重不足,则保护规划应优先考虑建立大型森林保护区和增加景观中森林覆盖率。此外也要加强公众保护意识宣传教育,对黑叶猴分布地区的当地农民灌输保护意识,普及野生动物保护法,使其充分认识保护黑叶猴和其他珍稀动植物及其栖息地的重要性。
