油棕Δ9硬脂酰-ACP脱氢酶EgSAD基因的克隆及表达分析
2022-03-07王梦娣李睿曹红星金龙飞李新国
王梦娣 李 睿 曹红星 金龙飞 李新国

摘 要:9硬脂酰-ACP脫氢酶(9 stearoyl-ACP dehydrogenase,SAD)是植物中重要的脂肪酸脱氢酶,在调控不饱和脂肪酸合成中发挥着重要作用。本研究以非洲油棕和杂交油棕为材料,在油棕果实油脂积累期,分别测定花后120、140、160 d 3个时期的脂肪酸组分;从油棕中克隆SAD基因,并对其理化性质、进化关系、启动子顺式作用元件进行生物信息学分析;利用实时荧光定量PCR检测SAD在2个油棕品种果实成熟期的表达特征。结果表明:非洲油棕中总不饱和脂肪酸(棕榈酸和硬脂酸)比例(55.76%~60.27%)显著高于杂交油棕(37.2%~45.43%),杂交油棕中不饱和脂肪酸(油酸和亚油酸)比例(51.61%~56.92%)显著高于非洲油棕(35.54%~38.64%);鉴定了7个基因命名为:~和~;基因编码的肽链平均为413个氨基酸,分子量为43.41~45.92 kDa,等电点为5.99~7.13,蛋白不稳定指数为44.07~53.25,总平均亲水性为‒0.496~‒0.405。基因编码的肽链平均为405个氨基酸,分子量为44.94~49.39 kDa,等电点为6.05~7.14,蛋白不稳定指数为43.44~44.61,总平均亲水性为‒0.554~‒0.489。多序列比对分析,油棕SAD基因氨基酸序列中存在典型SAD特征的保守组氨酸富集区:EENRHG和DEKRHE。在SAD的启动子上鉴定出植物激素响应、逆境胁迫响应、光响应和逆境响应顺式作用元件。在油棕果实成熟过程中呈上调表达趋势,和在杂交油棕中表达量显著高于非洲油棕,s在杂交油棕果实花后140 d中表达量最高,在杂交油棕的3个时期中均呈现先升高后下降的趋势,且在花后120 d的表达量显著高于花后120 d和160 d。本研究结果为进一步研究SAD调控油棕不饱和脂肪酸合成的机制奠定基础。
关键词:油棕;SAD基因;脂肪酸;生物信息学;基因表达
中图分类号:Q949.748.5 文献标识码:A
Identification and Expression Analysis of 9 Stearoyl-ACP Desaturase Gene in Oil Palm
WANG Mengdi LI Rui CAO Hongxin JIN Longfei LI Xinguo
1. College of Horticulture, Hainan University / Key Laboratory of Tropical Horticultural Crop Quality Regulation of Hainan Province, Haikou, Hainan 570228, China; 2 Coconut Research Institute, Chinese Academy of Tropical Agricultural Science / Hainan Key Laboratory of Tropical Oil Crops Biology, Wenchang, Hainan 571339, China
9 Stearoyl-ACP dehydrogenase (SAD), an important fatty acid dehydrogenase in plants, plays an important role in regulating the synthesis of unsaturated fatty acids. Oil palm is an important woody oil crop in the world. In this study, the saturated fatty acid and unsaturated fatty acid components of African oil palm and hybrid oil palm were determined at 120, 140 and 160 days after flowering (DAF) during the oil accumulation period, 120 to 160 (DAF). The physicochemical properties, evolutionary relationships and -acting elements of promoters were analyzed through bioinformatics. The expression characteristics of SAD genes in two oil palm varieties at fruit maturity stage were detected using real-time fluorescence quantitative PCR. Physiological measurement results showed that the proportion of total unsaturated fatty acids (palmitic acid and stearic acid) in African oil palm (55.76%‒60.27%) was significantly higher than that in hybrid oil palm (37.2%‒45.43%). The proportion of unsaturated fatty acids (oleic acid and linoleic acid) in hybrid oil palm (51.61%‒56.92%) was significantly higher than that in African oil palm (35.54%‒38.64%). Seven SAD genes were cloned from oil palm and were named ‒and ‒The results of physicochemical properties analysis showed that genes encoded a peptide chain with an average of 413 amino acids, a molecular weight of 43.41‒45.92 kDa, an isoelectric point of 5.99‒7.13, a protein instability index of 44.07‒53.25, and a total average hydrophilicity of ‒0.496 to ‒0.405. The average peptide chain of genes was 405 amino acids, the molecular weight was 44.94‒49.39 kDa, the isoelectric point was 6.05‒7.14, the protein instability index is 43.44‒44.61, and the total average hydrophilicity is ‒0.554 to ‒0.489. The conserved histidine rich regions with typical SAD characteristics were EENRHG and DEKRHE in the amino acid sequence of oil palm SAD genes. The results of bioinformatics analysis showed that the Cis-acting elements on plant hormone response, stress response, and light response were identified in the SAD promoter. The expression of was up-regulated during the ripening process of the oil palm fruit. The expression levels of and in the hybrid oil palm were significantly higher than those in the African oil palm. The expression levels of s were the highest in the 140 (DAF) of the hybrid oil palm fruit and the expression level of hybrid oil palm at 120 days after anthesis was significantly higher than that at 120 days and 160 (DAF). This study would lay a foundation for further study on the unsaturated fatty acids synthesis mechanism of SAD gene in oil palm.
oil palm; SAD gene; fat acid; bioinformatics; gene expression
10.3969/j.issn.1000-2561.2022.02.002
油棕是重要的热带木本油料作物,其果肉压榨的棕榈油广泛应用于餐饮业、食品工业、化工、机械润滑和生物能源等领域。棕榈油的脂肪酸组分主要有棕榈酸、硬脂酸、油酸和亚油酸,其中饱和脂肪酸含量较高,达48%以上。已有研究结果表明,过量摄取饱和脂肪酸增加帕金森症和心脑血管疾病的风险,通过遗传改良培育高不饱和脂肪酸油棕品种,降低棕榈油的棕榈酸和软脂酸等饱和脂肪酸含量,提高油酸、亚油酸和亚麻酸等不饱和脂肪酸含量,对提升棕榈油品质和扩大消费市场至关重要。
植物脂肪酸的合成主要在质体中进行,以糖酵解产生的丙酮酸为底物从头合成,生成16~18个碳的饱和脂肪酸,饱和脂肪酸在脱氢酶的作用下合成不饱和脂肪酸。质体中的硬脂酰酰基载体蛋白(ACP)在9硬脂酰-ACP脱氢酶(stearoyl- ACP desaturase,SAD)的作用下,去饱和生成含油不饱和双键的油酰基ACP,然后在硫酯酶的作用下解离,转运至内质网进行三酰甘油的组装,或者继续延长、脱氢合成其他不饱和脂肪酸。SAD是植物脂肪酸合成中的关键节点,是不饱和脂肪酸合成中的关键酶,对调节脂肪酸的组分具有重要意义。近年来,在拟南芥、花生、油菜、可可、棉花、大豆和油橄榄等植物中鉴定了编码SAD。在烟草中超量表达黄羽扇豆的SAD基因,能显著提高叶片油酸的含量。采用反义链降低棉花中SAD的表达量,棉籽油中硬脂酸含量由2%升高到40%。在莱茵衣藻中也发现,采用人工合成miRNA介导基因沉默的方式降低SAD的表达量,与野生型相比,转基因系的硬脂酸含量增加1倍。研究人员也在非洲油棕中克隆了2个SAD基因,表达分析发现其在果实发育中后期的表达量较高,但对于SAD调控油棕不饱和脂肪酸合成的機制还不明确。随着油棕基因组数据的公布,基于全基因组数据挖掘功能基因成为可能。本研究在非洲油棕中挖掘了4条,在杂交油棕中挖掘了3条,分析其理化性质、进化关系、启动子顺式作用元件,并采用荧光定量PCR分析其在油棕果实成熟过程中的表达特征,为后续研究SAD基因在油棕的生物学功能和遗传改良奠定基础。
材料与方法
材料
实验材料采自中国热带农业科学院椰子研究所基地(19°33′ N,110°47′ E)。分别采集‘热油4号’非洲油棕()和杂交油棕(× )花后120 d(S1)、140 d(S2)和160 d(S3)的果肉。样品采集后液氮速冻,储存在‒80℃冰箱中以备RNA提取。
方法
1.2.1 油棕脂肪酸含量 油棕果肉的脂肪酸采用GC-MS色谱法测定。
1.2.2 油棕SAD基因家族的挖掘与生物信息学分析 油棕全基因组数据从NCBI数据库(https://www.ncbi.nlm.nih.gov/,非洲油棕ID:2669,美洲油棕ID:24188)中下载,拟南芥SAD蛋白质序列从TAIR数据库(https://www.arabidopsis.org/)中下载。以拟南芥的SAD蛋白质序列作为参考序列,通过BLSATP在油棕基因组数据库中进行比对。基因全长采用生工生物工程(上海)股份有限公司的高保真PCR Mix预混液克隆。利用在线工具ExPAsy(http://www.expasy.org)分析油棕SAD基因家族的蛋白质分子量、等电点和总平均亲水性等理化性质。从NCBI数据库中下载油棕SAD基因CDS上游2000 bp的序列,利用Plant Care进行顺式作用元件分析,采用Tbtool进行可视化分析。使用在线分析工具 Clustal Omega(http://www.ebi.ac.uk/Tools/ msa/ clustalo/)比对油棕SAD基因编码氨基酸序列,使用在线工具 BoxShade显示比对结果(http:// www.ch.embnet.org/software/BOX_form.html)。
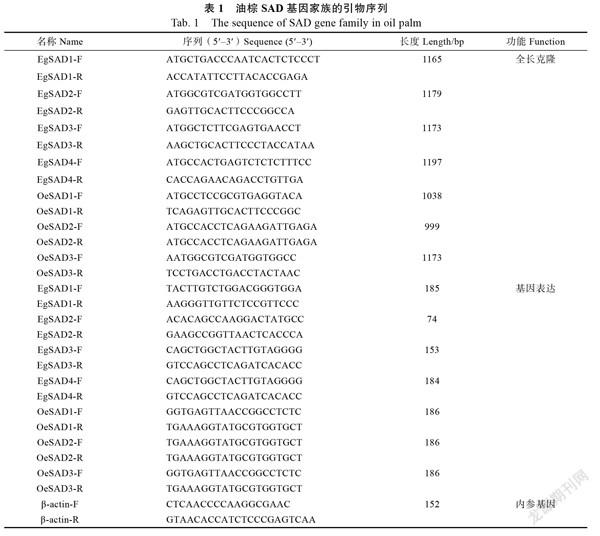
1.2.3 RNA提取及基因表达分析 油棕果肉的总RNA提取采用生工生物工程(上海)股份有限公司的Spin Column Plant Total RNA Purification Kit试剂盒,cDNA合成采用北京全式金生物技术有限公司的HiScript II 1st Strand cDNA Synthesis试剂盒进行,具体步骤参照试剂盒的说明书。实时荧光定量PCR采用赛默飞世尔科技(中国)有限公司的SYBR Select Master Mix进行分析,以β-actin作为内参基因,反应体系参照试剂盒的说明书,设置3个生物学重复,引物见表1,采用2法计算目标基因的相对表达量。
数据处理
采用SPSS 26.0软件进行数据分析,用Duncan’s检测法进行脂肪酸含量和基因表达的差异显著性分析。
结果与分析
油棕果肉的主要脂肪酸种类和含量
由表2可知,油棕果肉的主要脂肪酸种类有棕榈酸、硬脂酸、油酸、和亚油酸,月桂酸、豆蔻酸、亚麻酸等含量较低归入其他类,非洲油棕中不饱和脂肪酸(棕榈酸和硬脂酸)比例(55.76%~ 60.27%)显著高于杂交油棕(37.2%~45.43%),杂交油棕中不饱和脂肪酸(油酸和亚油酸)含量比例(51.61%~56.92%)显著高于非洲油棕(35.54%~38.64%)。
油棕基因的克隆、编码蛋白质的理化性质分析及亚细胞定位预测
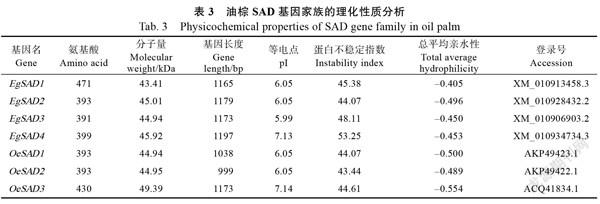
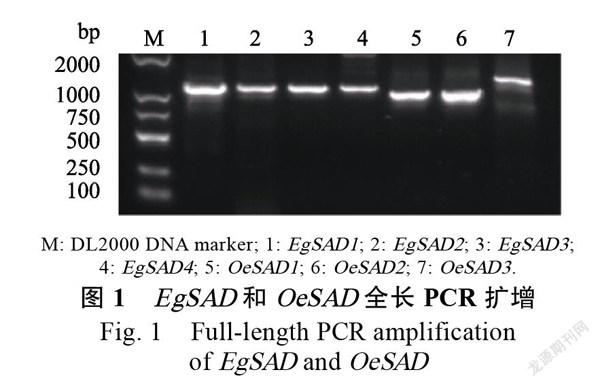
通过BLASTP比对在非洲油棕和美洲油棕的基因组中鉴定了7个SAD候选蛋白质,根据其与拟南芥的同源性顺序进行命名,依次命名为~、~。分别以油棕果实和杂交油棕果实的cDNA为模板,克隆
SAD基因(图1)。基因长度为1165 bp,编码471个氨基酸;基因长度为1179 bp,编码393个氨基酸;基因长度为1173 bp,编码391个氨基酸;基因长度为1197 bp,编码399个氨基酸;基因长度为1038 bp,编码393个氨基酸;基因长度为999 bp,编码393个氨基酸;基因长度为1173 bp,编码430个氨基酸。非洲油棕EgSAD1~EgSAD4蛋白质不稳定系数分别为45.38、44.07、48.11和53.25;总平均亲水性为‒0.405、‒0.496、‒0.450和‒0.453。杂交油棕OeSAD1~OeSAD3蛋白质不稳定系数分别为44.07、43.44和44.61;总平均亲水性为‒0.500、‒0.489和‒0.554(表3)。
非洲油棕和杂交油棕的进化分析
将油棕的SAD蛋白序列与其他作物进行同源性比对,并采用MEGA 6.0进行进化关系分析。结果发现,EgSAD1和AtSAD1、TcSAD5聚在一起,EgSAD2、EgSAD3、EgSAD4、OeSAD1、OeSAD2、OeSAD3和TcSAD9聚在一起(图2)。
启动子顺式作用元件的鉴定
在非洲油棕和杂交油棕SAD基因的启动子中鉴定出7种顺式作用元件(图3),其中包含光响应(25个)、茉莉酸甲酯响应(4个)、脱落酸响应(8个)、逆境响应(2个)、干旱响应(4个)、水杨酸响应(4个)和赤霉素响应(1个)。
油棕蛋白氨基酸序列比对
由图4可知,油棕SAD蛋白氨基酸序列中存在典型的保守组氨酸富集区:EENRHG和DEKRHE(图4中的红色框),天冬氨酸(D)和组氨酸(H)为催化活性中心的二价铁离子提供结合位点,保证了油棕SAD脱氢酶具有一定的催化活性。
油棕基因的表達特征分析
采用实时荧光定量PCR对4个基因和3个基因分别在非洲油棕和杂交油棕花后120、140、160 d的果中进行表达特征分析(图5)。基因在2个品种中的表达量随着果实发育时期逐渐增加,在花后140 d和160 d,基因在杂交油棕种中的表达量显著高于非洲油棕。基因在2个品种中均有表达,且在杂交油棕种中的表达量显著高于非洲油棕。基因在非洲油棕花后160 d的表达量显著高于花后120 d和140 d,在杂交油棕中仅在花后120 d时有少量表达。基因在非洲油棕和杂交油棕中均有表达,但在非洲油棕中的表达量显著高于杂交油棕。、和基因在杂交油棕的3个时期中均呈现先升高后下降的趋势,且在花后120 d的表达量显著高于花后120 d和160 d。
讨论
油棕是重要的热带油料作物,产油效率极高,但棕榈油中主要是饱和脂肪酸,过量摄取有增加患心脑血管疾病的风险。因此,降低油棕中饱和脂肪酸含量,提高油酸、亚油酸和亚麻酸等不
饱和脂肪酸含量的比例,对提升棕榈油品质至关重要。SAD主要在质体中催化与ACP结合的硬脂酸或软脂酸的9号和10号碳原子之间去饱和形成油酸,在植物细胞不饱和脂肪酸的合成中发挥重要作用。本研究在非洲油棕和杂交油棕中克隆了7个SAD基因,并初步探究了SAD的功能。MANUEL 等研究发现亚麻荠和澳洲坚果中的SAD是可溶性酶。在本研究中,7个基因总平均亲水性均为负值,表现为亲水性。典型SAD含有特征的保守组氨酸序列富集区:EENRHG和DEKRHE。多序列比对结果显示,油棕SAD蛋白氨基酸中含有特征的保守组氨酸,保证脱氢酶具有催化活性。进化分析结果发现,油棕的SAD基因与可可的SAD聚在一起,可能是因为油棕和可可都是热带木本油料作物,有着相似的进化关系。
油棕果实脂质积累主要在成熟期,从花后120 d开始迅速积累,在花后160 d达到峰值。因此,本研究分析了7个SAD基因在油棕果实花后120、140、160 d的表达特征,发现在非洲果实成熟过程中表达逐渐升高,在花后160 d表达量最高,这一结果与油棕本身油脂积累规律一致。在油茶种子发育过程中油酸含量持续上升,SAD基因持续高表达。在油橄榄果肉油脂快速积累期,2个SAD基因的表达量达到峰值。在本研究中,7个SAD基因在油棕果肉成熟期均呈逐渐升高或者先增高后降低的趋势,这表明SAD基因在油棕油脂快速积累期起重要的调控作用。在山核桃中,研究发现基因的表达与油酸含量密切相关,随着山核桃油酸含量上升,基因表达上升,山核桃不饱和脂肪酸比例增加。在本研究中,杂交油棕的不饱和脂肪酸含量显著高于非洲油棕,和在杂交油棕中的表达量显著高于非洲油棕,同时杂交油棕还存在3个来自于美洲油棕亲本的。进而推测杂交油棕的不饱和脂肪酸高的原因可能是由于SAD基因的高表达。油棕果肉发育的不同阶段也受不同激素的调控,本研究也对和启动子顺式作用元件进行预测和分析,发现包含逆境响应、脱落酸响应、茉莉酸甲酯响应等在内的7种顺式作用元件。已有研究发现,菊花在盐胁迫下SAD基因表达量升高,并且伴随饱和脂肪酸含量的减少和不饱和脂肪酸含量的升高,推测在油棕不饱和脂肪酸的积累也受到激素或非生物逆境的影响。
本研究从油棕中克隆出7个SAD基因,通过生物信息学和表达分析发现油棕SAD基因在油棕不饱和脂肪酸合成中发挥作用,为提高油棕的不饱和脂肪酸含量并改良油棕提供了思路。SAD基因在油棕不饱和脂肪酸合成中的功能和逆境响应作用机制有待进一步研究,后续可通过基因过表达和基因敲除等实验进行验证。
参考文献
- KAZAZ S, BARTHOLE G, DOMERGUE F, ETTAKI H, TO A, VASSELON D, DE V D, BELCRAM K, LEPINIEC L, BAUD S. Differential activation of partially redundant 9 stearoyl-ACP desaturase genes is critical for omega-9 monounsaturated fatty acid biosynthesis during seed development in [J]. The Plant Cell, 2020, 32(11): 1-25.
- KACHROO A, SHANKLIN J, WHITTLE E, LAPCHYK L, HILDEBRAND D, KACHROO P. The stearoyl-acyl carrier protein-desaturase family and the contribution of leaf isoforms to oleic acid synthesis[J]. Plant Molecular Biology, 2007, 63(2): 1-16
- 東金玉, 万勇善, 刘风珍. 花生9-硬脂酰-ACP脱氢酶基因(SAD)的序列分析[J]. 作物学报, 2012, 38(7): 1167-1177.DONG J Y, WAN Y S, LIU F Z. Analysis of 9-stearoyl- ACP dehydrogenase gene in peanut[J]. Acta Agronomica Sinica, 2012, 38(7): 1167-1177. (in Chinese)
- 贾艳丽, 吴 磊, 卢长明. 甘蓝型油菜9硬脂酰ACP脱氢酶(SAD)基因的克隆与表达分析[J]. 中国油料作物学报, 2014, 36: 135-141.JIA Y L, WU L, LU C M. Cloning and expression analysis of 9 stearyl ACP dehydrogenase (SAD) gene in [J]. Chinese Journal of Oil Crops, 2014, 36: 135-141. (in Chinese)
- ZHANG Y, MAXIMOVA S N, GUILTINAN M J. Characterization of a stearoyl-acyl carrier protein desaturase gene family from chocolate tree, L.[J]. Frontiers in Plant Science, 2015, 6: 239.
- 邓咪咪, 刘宝玲, 王志龙, 薛金爱, 张红梅, 李润植. 大豆硬脂酰-ACP Δ9脱氢酶()基因家族的鉴定及功能分析[J]. 生物工程学报, 2020, 36(4): 716-731.DENG M M, LIU B L, WANG Z L, XUE J A, ZHANG H M, LI R Z. Identification and functional analysis of soybean stearoyl-ACP Δ9 dehydrogenase () gene family[J]. Biological Engineering Journal, 2020, 4(4): 716-731. (in Chinese)
- KNUTZON D S, THOMPSON G A, RADKE S E, JOHNSON W B, KNAUF V C, KNDL J C. Modification of seed oil by antisense expression of a stearoyl-acyl carrier protein desaturase gene[J]. Proceedings of the National Academy of Sciences of the United States of America, 1992, 89(7): 2624-2628.
- JAEGER L D, SPRINGER J, WOLBENS E J H, MARTENS D E, EGGINK G, WIJFFELS R H. Gene silencing of stearoyl-ACP desaturase enhances the stearic acid content in [J]. Bioresource Technology, 2017, 245: 1616-1626.
- AKMAR A, CHEAH S C, AMINAH S. Characterization and regulation of the oil palm () stearoyl-ACP desaturase genes[J]. Journal of Oil Palm Research, 1999: 1-17.
- SHAH F H, RASHID O, SAN C T. Temporal regulation of two isoforms of cDNA clones encoding delta 9-stearoyl-ACP desaturase from oil palm ()[J]. Plant Science, 2000, 152(1): 27-33.
- KRUTKAEW V, SRIRAT T. Cloning and characterization of stearoyl-ACP desaturase gene (SAD) in oil palm (Jacq.)[J]. Thai Journal of Genetics, 2013, 6(1): 60-64.
- MANUEL F R, ALICIA S G, JOAQUIN J S, RAFAEL G, ENRIQUE M F. Characterization of soluble acyl-ACP desaturases from , and [J]. Journal of Plant Physiology, 2015, 178: 35-42.
-
收稿日期 2021-08-12;修回日期 2021-09-19
基金项目 中国热带农业科学院基本科研业务费专项资金(No. 1630152021004)。
作者简介 王梦娣(1996—),女,硕士研究生,研究方向:热带木本油料作物遗传育种。*通信作者(Corresponding auther):
金龙飞(JIN Longfei),E-mail:jlf_0511@163.com;李新国(LI Xinguo),E-mail:lixinguo13@163.com。
