2种羌活药用植物的叶绿体基因组密码子使用偏性分析
2022-03-04王琳璇王高烽谷威赵志敏马永贵
王琳璇 王高烽 谷威 赵志敏 马永贵



摘要 对羌活(Notopterygium incisum)与宽叶羌活(N.franchetii)的叶绿体编码基因的密码子使用偏性(codon usage bias)及其影响因素进行了分析。结果表明:羌活和宽叶羌活的叶绿体基因组总ENC值分别为49.79和49.70,偏性较弱,密码子第3位碱基倾向于使用A/T。PR2-plot分析表明,选择压力和突变都是影响2种羌活叶绿体密码子使用偏性的主要因素;而中性绘图分析和ENC-plot分析表明选择压力是影响2种羌活叶绿体密码子使用偏性的主要因素。相对同义密码子使用度(RSCU)分析确定了2种羌活中使用频率较高的30个高频密码子,几乎都以A或U结尾,只有UUG以G结尾。羌活的最优密码子9个,宽叶羌活的最优密码子7个。2种羌活有5个最优密码子相同,分别是UUG、CAU、CGU、AGU、GGA。该研究结果为后续药用羌活植物的基因表达、蛋白质翻译和遗传育种研究提供了研究依据和基础数据。
关键词 羌活;宽叶羌活;叶绿体基因组;密码子使用偏性;最优密码子
中图分类号 Q 943.2 文献标识码 A 文章编号 0517-6611(2022)03-0095-07
doi:10.3969/j.issn.0517-6611.2022.03.025
Codon Usage Bias in Chloroplast Genomes of Two Medicinal Species of Notopterygium H. de Boissieu
WANG Lin-xuan, WANG Gao-feng, GU Wei et al
(College of Life Sciences, Qinghai Normal University, Xining, Qinghai 810000)
Abstract This study analyzed the codon usage bias and affecting factors of the protein coding genes (CDS) of the chloroplast genomes of two medicinal plants, Notopterygium incisum and N. franchetii. The results showed that the chloroplast genomes of N. incisum and N. franchetii had a total ENC value of 49.79 and 49.70, respectively, indicating weak codon usage bias. The third base of the codon of the two chloroplast genomes preferred to use A/T. The analysis of PR2-plot showed that selection pressure and nutation were the main factors affecting the bias of codon usage in N. incisum and N. franchetii CDS, while neutrality plot and ENC-plot analyses both illustrated selection pressure was the main factors affecting codon usage bias in the two species. Relative Synonymous Codon Usage (RSCU) analysis determined 30 high-frequency codons used in N. incisum and N. franchetii, almost all ending with A or U, with only UUG ends with G. There were nine optimal codons identified in N. incisum, and seven optimal codons found in N. franchetii. The five optimal codons shared by N. incisum and N. franchetii were namely UUG, CAU, CGU, AGU, GGA. The results provide important basal data for the future study of gene expression, protein translation and molecular breeding of medicinal Notopterygium plants.
Key words Notopterygium incisum;Notopterygium franchetii;Chloroplast genome;Codon usage bias;Optimal codons
基金項目 青海省基础研究计划项目(2018-ZJ-741)。
作者简介 王琳璇(1994—),女,河北廊坊人,硕士研究生,研究方向:生物化学与分子生物学。
通信作者,教授,博士,从事动植物分子进化与适应研究。
收稿日期 2021-04-03
密码子在蛋白质合成过程中起着传递遗传信息的关键作用,在大多数情况下,编码同一氨基酸的同一密码子的使用频率是不同的,这一现象称为密码子使用偏性(codon usage bias)。某一物种或某一基因通常倾向于使用一种或几种特定的同义密码子,这些密码子被称为最优密码子(optimal codon)。影响密码子使用偏性的因素很多,最主要的因素是定向突变和中性选择[1-3],还包括GC含量、同义替换率[4]、tRNA丰度[5]、有效翻译起始位点的选择[6]、密码子亲水性和DNA复制起始位点[7]、基因长度[8]和表达水平[9]等。在不同物种中,突变和选择的影响强度不同。例如在蓝藻中,影响密码子使用偏性较大的是GC组成与选择压力[10],但在果蝇中则是突变对的密码子使用偏性影响较大[3]。
叶绿体是植物进行光合作用的器官,也参与到植物的氨基酸合成等通路中。作为植物重要的细胞器,叶绿体有自己一套独特的遗传信息[11]。针对叶绿体基因组的密码子使用偏性研究,可以帮助了解叶绿体蛋白质编码基因的使用偏好,为后续遗传改造和基因表达研究提供基础数据。研究发现,乳油木叶绿体基因组密码子使用偏性更多受突变影响[12],选择压力对其使用偏性影响较小。杜梨叶绿体密码子使用偏性则主要受选择的影响[13]。胡晓艳等[14]在对酸枣叶绿体基因组的研究中获得了7个最优密码子。
羌活属(Notopterygium Boissieu)植物隶属于伞形科(Apiaceae)美味芹族,是我国特有的多年生草本药用植物,主要分布在我国西南部高山地区[15]。据《中国植物志(英文版)》记载,该属包括宽叶羌活、羌活、羽苞羌活、澜沧羌活、卵叶羌活和细叶羌活6个种[16]。《中国药典》规定的羌活基源植物为羌活与宽叶羌活,入药部位为干燥根及根茎,具有解表散寒、祛风除湿和止痛的功效。羌活主要产于四川、云南,次产于青海及甘肃;宽叶羌活主产于甘肃、青海,次产于四川、山西和内蒙古等地。近年来,由于过度采挖及生境受到严重破坏,野生羌活的自然群体已经面临濒危,被载入《中国生物多样性红色名录(高等植物卷)》[17]。羌活作为一味传统的中药,富含异欧芹素、羌活醇、阿魏酸等活性物质,具有抗炎、抗氧化、抗心律失常、抗菌、抗癌细胞增殖等药效作用[18]。目前有关羌活属叶绿体基因组密码子使用偏性研究鲜见报道。该研究拟对2种可以入药的羌活属植物,即羌活和宽叶羌活进行叶绿体基因组的密码子使用模式分析,研究结果不但可以阐明羌活光合作用相关基因的使用偏好,也可为羌活基因表达研究及遗传育种研究提供理论基础。
1 材料与方法
1.1 数据获取
羌活样品采集于青海省果洛藏族自治州达日县,99.46° E,33.77° N,海拔3 943.24 m。宽叶羌活样品采自青海省玛可河麦浪沟,100.96° E,32.84° N,海拔3 790.78 m。保存于干冰中,送于北京百迈客生物科技有限公司进行叶绿体基因组测序。利用Illumina HiSeq X Ten 平台测序,使用MIRA序列组装软件的MIRA v3.4.1.1模块进行,最终得到羌活的叶绿体全基因组序列长度为 157 050 bp。宽叶羌活的叶绿体全基因组序列长度为 142 184 bp。测序结果提交至NCBI(national center for biotechnology information),登录号:羌活MW820163、宽叶羌活MW820162。使用Geneious11.1.5 提取编码基因(coding sequence,CDS)并筛选,剔除小于300 bp的基因,并选择起始密码子为ATG的编码基因进行后续分析,最终羌活获得51条编码基因,宽叶羌活获得50条编码基因。
1.2 分析方法
1.2.1 密码子相关参数计算。
使用软件codonW1.4.2分析获得的编码基因,计算有效密码子数(effective number of codon,ENC)。ENC范围一般在20~61,越接近20,表示密码子使用偏性越大。使用在线分析软件CUSP(http://embossgui.sourceforge.net/demo/)对叶绿体基因组密码子的GC含量等碱基组成信息进行研究,并对相关参数进行相关性分析[19]。
1.2.2 中性绘图分析。
分别以CGC1、CGC2、CGC3表示密码子第1、2、3位的GC含量,CGC12表示CGC1与CGC2的平均数,以CGC3和CGC12为横、纵坐标绘制散点图,分析叶绿体基因组密码子第3位和前2位的相关性,从而分析影响密码子偏性的因素[20]。以CGC3、CGC12为横、纵坐标绘制散点图,如果CGC3和CGC12呈显著相关,说明密码子第3位的碱基组成与前2位相似,密码子偏性主要受突变的影响;如果CGC3和CGC12无显著相关关系,趋势线斜率接近0,则说明密码子第3位与前2位碱基组成差异较大,密码子碱基相对保守,这种情况密码子偏性更多受选择的影响[21]。
1.2.3 PR2-plot绘图分析。
为进一步分析影响2种羌活叶绿体基因组密码子偏性的因素,以CA3、CT3、CC3、CG3分别表示密码子第3位A、T、C、G 4种碱基的含量,以CG3/(CG3+CC3)为横坐标,CA3/(CA3+CT3)为纵坐标绘制散点图。如果大部分基因分布在图像上半部,说明密码子第3位碱基含量A>T,反之则T>A,同理如果大部分基因分布在图像右半部,说明密码子第3位碱基含量G>C,反之则C>G。若点在图中均匀分布,则密码子偏性完全受突变影响[20-21]。
1.2.4 ENC-plot绘图分析。
以GC3为横坐标,以ENC值为纵坐标绘制散点图,并以ENC期望绘制标准曲线。
ENC期望=2+CGC32+29CGC32+(1-CGC3)2
标准曲线表示在无选择压力的情况下ENC与CGC3的关系,如果点主要分布于曲线附近,说明密码子偏性更多受突变的影响,如果点分布于距标准曲線较远的位置,则说明密码子偏性更多受选择的影响。根据公式(ENC期望-ENC实际)/ENC期望制作ENC分布频数表,统计ENC实际与ENC期望的差距[22]。
1.2.5 RSCU分析。
使用CodonW计算2种羌活叶绿体基因组的RSCU(relative synonymous codon usage)。RSCU是实际观察到的某密码子使用频率与预期(该密码子编码的氨基酸的同义密码子使用频率相等时)使用频率的比值。理论值与预期值相等时,密码子不存在偏性。所以当RSCU=1.00,表明密码子使用无偏性;RSCU>1.00时,表明该密码子实际使用频率较高;RSCU<1.00时,表明实际使用频率较低[23]。
1.2.6 最优密码子分析。
选出2种羌活叶绿体编码基因密码子ENC值最大的10%和最小的10%,分别建立高低表达库,选出库中ΔRSCU>0.08且整体RSCU>1.00的密码子[24]。
2 结果与分析
2.1 密码子组成
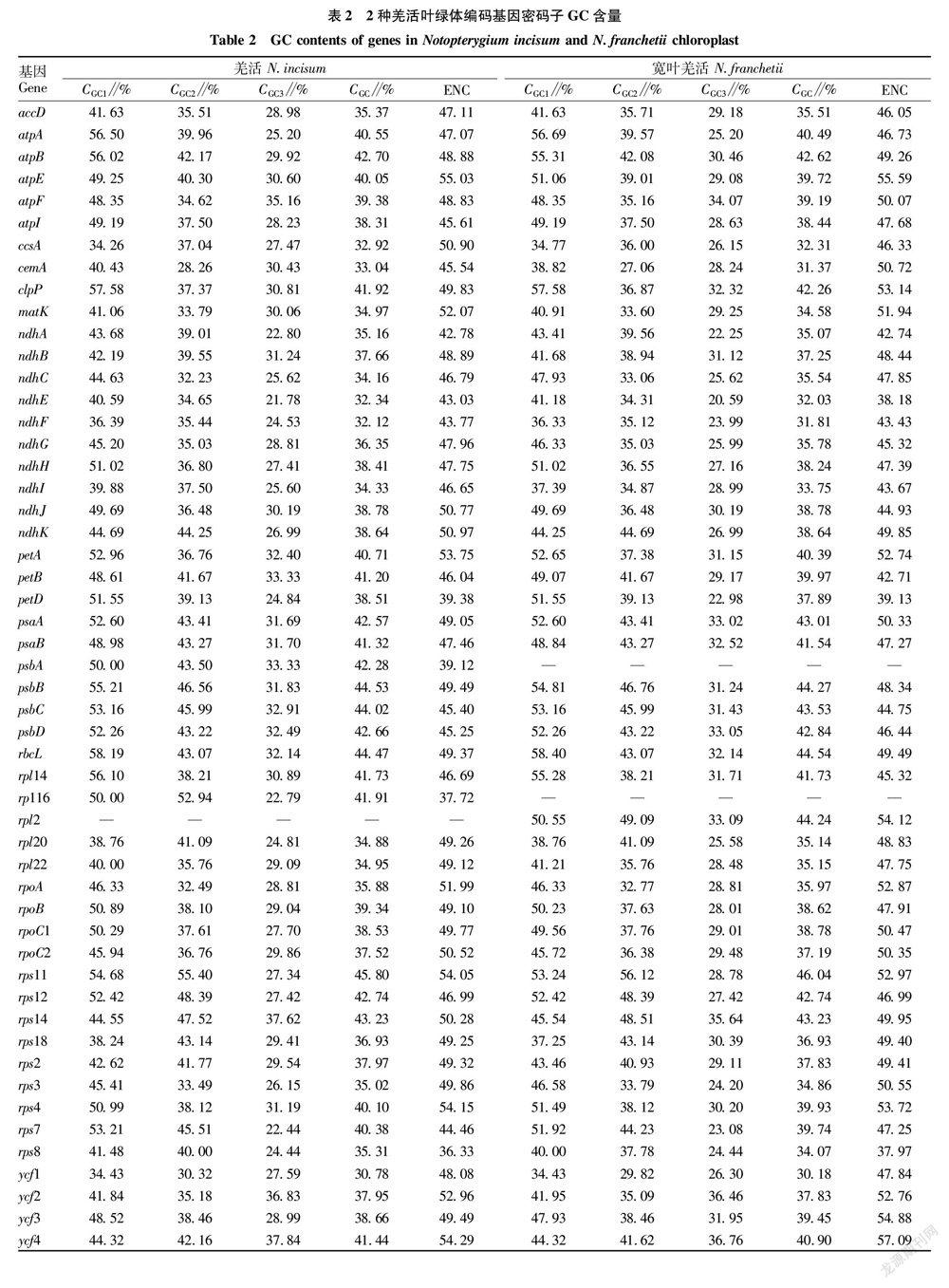
2种羌活叶绿体基因组密码子第3位的碱基组成结果显示,羌活的叶绿体基因组密码子第3位的CA3、CT3分别为42.75%与46.97%(表1),明显高于CC3、CG3,说明羌活叶绿体基因组密码子多以AT结尾;宽叶羌活的密码子第3位碱基组成与羌活类似,AT含量较高。 羌活与宽叶羌活的叶绿体密码子第1位GC含量从第1位到第3位逐渐下降,第1位分别为46.16%、45.99%,第2位38.17%、37.91%,第3位29.97%、29.67%。2种羌活的ENC均大于49.00,说明2种羌活叶绿体基因组密码子偏性较弱。
使用CUSP统计2种羌活叶绿体基因组编码基因的CGC、CGC1、CGC2、CGC3、ENC,结果表明,羌活和宽叶羌活的叶绿体基因组编码基因的CGC平均值为38.64%和38.44%(表2),差异不大。羌活和宽叶羌活中CGC最高的基因皆为rps11,CGC最低的基因为ycf1。羌活和宽叶羌活编码基因的ENC值范围分别是36.33~55.03、37.97~57.09,而且羌活约有83%编码基因的ENC值大于45.00,宽叶羌活有79%编码基因的ENC值大于45.00,说明多数编码基因密码子使用偏性较弱。2种羌活的rps8的ENC值最小,分别为36.33和37.97,表明rps8在2种羌活叶绿体编码基因中偏性最强。
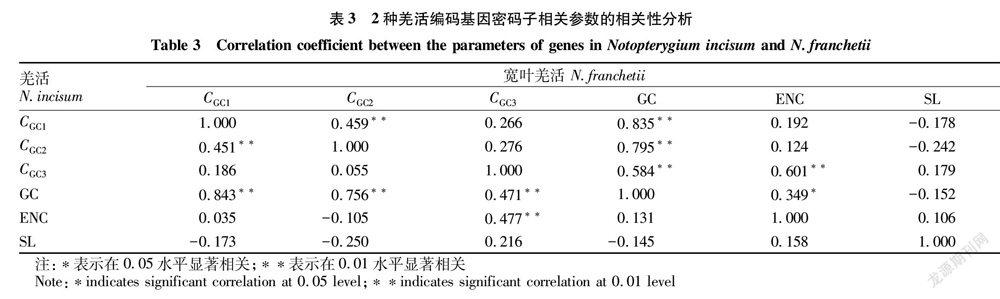
对2种羌活的叶绿体编码基因的CGC1、CGC2、CGC3、GC、ENC、叶绿体编码基因序列长度(sequence length,SL)等参数进行相关性分析,结果表明2种羌活的碱基组成相关性较接近,CGC1与CGC2皆为极显著相关(表3),且CGC1、CGC2都与CGC3无显著相关性;2种羌活的GC含量与第1、2、3位组成均极显著相关。羌活与宽叶羌活的SL与CGC1、CGC2、CGC3、GC、ENC均无显著相关性。2种羌活的相关性分析结果也存在一定差异,羌活的ENC与CGC3极显著相关,而宽叶羌活的ENC除了与CGC3极显著相关外,还与GC显著相关。
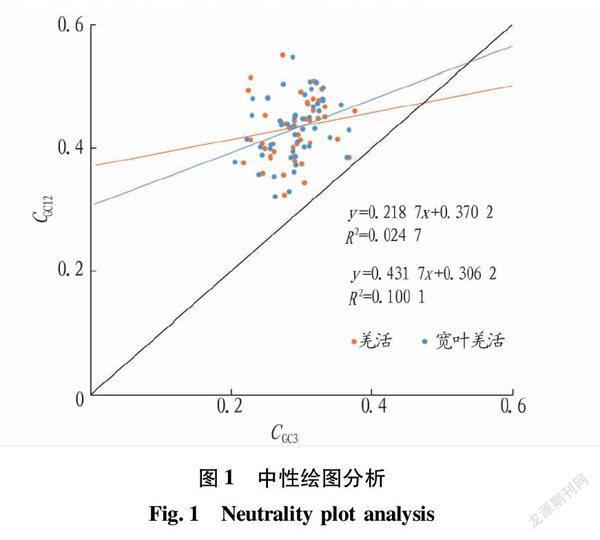
2.2 中性绘图分析
羌活的CGC12取值范围为 0.324~0.550(图1),CGC3与CGC12的回归系数为0.218 7,皮尔森系数为0.147,无显著相关性,说明羌活的叶绿体密码子第3位碱基组成与第1、2位差异较大,密码子偏性主要受选择的影响。宽叶羌活的CGC12的取值范围0.321~0.547(图1),CGC3与CGC12的回归系数为0.431 7,皮尔森系数为0.316,相关性较低,说明宽叶羌活的叶绿体密码子第3位碱基组成与第1、2位差异较大,密码子偏性受选择影响较大。
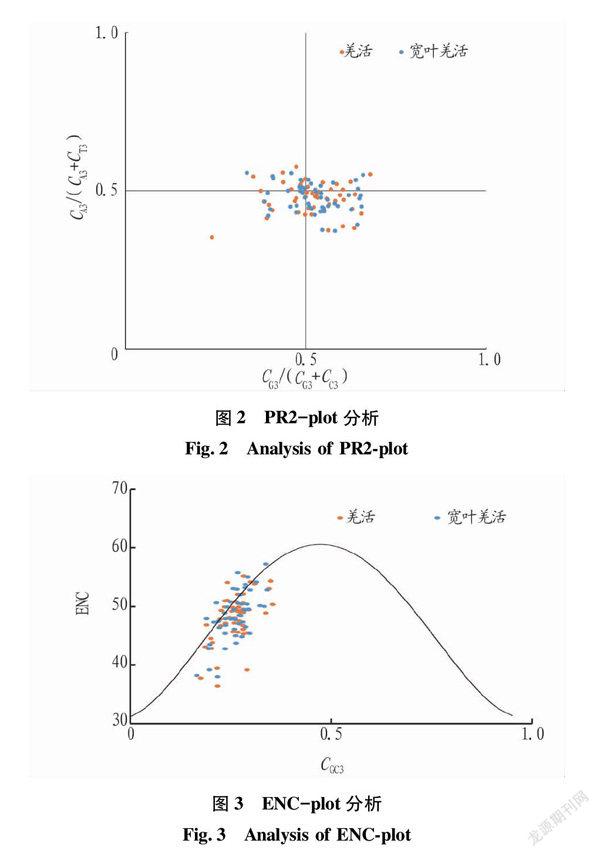
2.3 PR2-plot分析
PR2-plot分析以CG3/(CG3+CC3)为横坐标,CA3/(CA3+CT3)为纵坐标绘制散点图,结果表明,2种羌活大部分基因分布在右下方位置(图2),说明羌活和宽叶羌活的叶绿体基因组密码子不仅受到突变的影响,还受到选择的影响,并且密碼子第3位的碱基频率T>A,G>C。
2.4 ENC-plot分析
对2种羌活进行ENC-plot绘图分析,结果见图3。标准曲线表示在无选择压力的情况下,ENC与CGC3的关系。如果点主要分布于曲线附近,说明密码子偏
性更多受突变的影响,如果点分布于距标准曲线较远的位置,说明密码子偏性更多受选择的影响。2种羌活的编码基因在标准曲线两侧均有基因分布,大部分基因位于标准曲线下侧,部分基因距标准曲线较远。
为了进一步分析ENC实际值与标准曲线的偏离情况,制作ENC分布频数表,结果见表4,羌活有24个基因的比值在-0.05~<0.05,表明有24个基因的ENC实际值与期望值接近(ENC比值-0.05~<0.05),其余27个基因的ENC实际值与期望值相差较大(ENC比值>0.05或<-0.05),其中psbA、rps8、rp116、petD、psbC、psbD、petB的ENC实际值距标准曲线较远(ENC比值>0.15)。宽叶羌活有23个基因的比值在-0.05~<0.05,ENC实际值与期望值接近,27个基因ENC实际值与期望值差距较大,其中rps8、petB、petD、psbC、ndhE、ndhI的ENC实际值距标准曲线较远。说明2种羌活的叶绿体基因组密码子偏性受突变的影响较小,更多受选择的影响。
2.5 RSCU分析
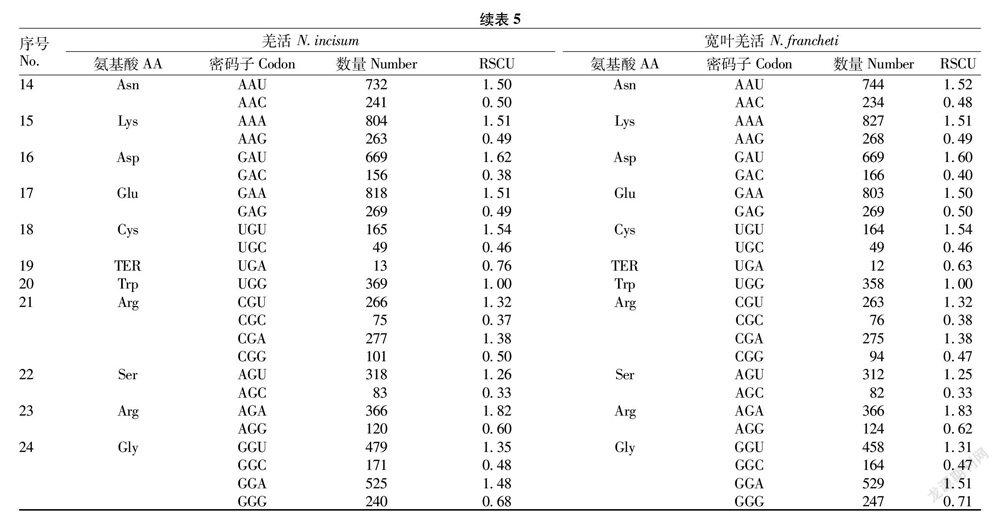
由表5可知,羌活和宽叶羌活各有30个密码子RSCU>1.00,说明各有30个密码子的使用有偏好性。实际使用频率偏高(RSCU>1.00)的密码子中结尾为U的有16个,结尾为A的有13个,只有UUG以G结尾。说明2种羌活叶绿体基因组密码子偏好以A、U结尾,其中U的偏好性略强于A。
2.6 最优密码子确定
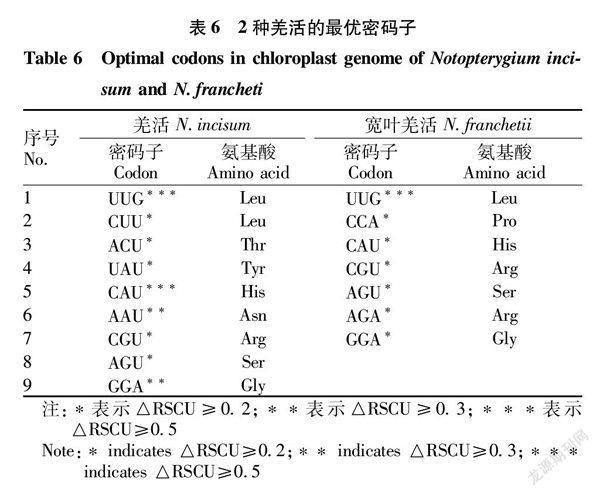
2种羌活的最优密码子均符合总RSCU>1.00,ΔRSCU≥0.08。2种羌活的最优密码子都多以A/U结尾,只有UUG以G结尾。确认了羌活的最优密码子9个(UUG、CUU、ACU、UAU、CAU、AAU、CGU、AGU、GGA),宽叶羌活的最优密码子7个(UUG、CCA、CAU、CGU、AGU、AGA、GGA)。2种羌活有5个最优密码子相同,分别是UUG、CAU、CGU、AGU、GGA(表6)。
3 讨论
该研究对羌活和宽叶羌活基因组密码子使用偏好性及其主要影响因素进行了探讨和比较。结果表明,ENC小于45的基因较少,多数基因ENC较高,说明2种羌活基因组密码子使用偏性均较弱。相关性分析结果表明,2种羌活的叶绿体基因组ENC都与CGC3显著相关,说明密码子第3位碱基组成对密码子偏性的影响较大,这与原晓龙等[25]对云南蓝果树的研究结论一致。而尹为治等[26]研究结果表明,坡垒的叶绿体基因组密码子第1位碱基组成对密码子偏性的影响更大。密码子组成分析显示,2种羌活的总GC和CGC3较低,且CGC1>CGC2>CGC3,密码子的第3位倾向于使用A/T,第3位碱基使用频率T>A,G>C。
突变和选择压力是2个影响密码子偏性的主要因素。PR2-plot分析结果发现,基因分布在图中发生了偏移,可知2种羌活的叶绿体密码子使用偏性不仅受到了突变的影响,还受到了选择压力的影响。中性绘图结果显示,2种羌活CGC12与CGC3相关性不显著,说明密码子第3位的碱基组成与前两位差异大,密码子相对保守,表明选择压力对密码子的影响较大。ENC-plot结果显示,2种羌活的多数叶绿体基因偏离标准曲线进一步证实选择压力是影响2种羌活叶绿体密码子使用偏性的主要因素。综上所述,2种羌活的叶绿体基因组密码子使用偏性受到突变和选择压力的影响,最主要的影响因素是选择压力。影响竹亚科的巴山木竹、印度簕竹、Olyra latifolia L.以及苦荞的叶绿体基因组密码子使用偏性的主要因素也是选择压力[27-28]。而突变对槲蕨属的川滇槲蕨、栎叶槲蕨的叶绿体基因组密码子使用偏性的影响更大[29]。
羌活和宽叶羌活的叶绿体基因组密码子使用模式较为相似。2种羌活的总GC含量与密码子各位的GC含量差异不大;叶绿体基因组密码子偏性都较弱;密码子第3位都倾向于使用A/T,且碱基含量T>A,G>C;影响密码子使用偏性的主要因素都是选择压力。
通过RSCU分析,确定了2种羌活中使用频率较高的30个高频密码子(RSCU>1.00),几乎都以A或U结尾,只有UUG以G结尾。确认了羌活的最优密码子9个(UUG、CUU、ACU、UAU、CAU、AAU、CGU、AGU、GGA),宽叶羌活的最优密码子7个(UUG、CCA、CAU、CGU、AGU、AGA、GGA)。2种羌活有5个最优密码子相同,分别是UUG、CAU、CGU、AGU、GGA。
该研究通过对2种羌活药用植物的叶绿体基因组编码基因的使用偏性进行比较分析,得出主要结论:①2种药用羌活叶绿体基因组的密码子使用偏性整体较为相似,且偏性较弱;②密码子的第3位倾向于使用A、T;③突变和选择压力在2种羌活的密码子使用偏性中起主要作用,其中选择压力对密码子使用偏性影响更大。该研究结果为后续药用羌活植物的基因表达、蛋白质翻译和遗传育种研究提供了研究依据和基础数据,有潜在的经济价值和应用价值。
参考文献
[1] MARAIS G,MOUCHIROUD D,DURET L.Neutral effect of recombination on base composition in Drosophila[J].Genetical research,2003,81(2):79-87.
[2] PRABHA R,SINGH D P,SINHA S,et al.Genome-wide comparative analysis of codon usage bias and codon context patterns among cyanobacterial genomes[J].Marine genomics,2017,32:31-39.
[3] VICARIO S,MORIYAMA E N,POWELL J R.Codon usage in twelve species of Drosophila[J].BMC evolutionary biology,2007,7:1-17.
[4] SHARP P M,LI W H.The rate of synonymous substitution in enterobacterial genes is inversely related to codon usage bias[J].Molecular biology and evolution,1987,4(3):222-230.
[5] OLEJNICZAK M,UHLENBECK O C.tRNA residues that have coevolved with their anticodon to ensure uniform and accurate codon recognition[J].Biochimie,2006,88(8):943-950.
[6] ZALUCKI Y M,POWER P M,JENNINGS M P.Selection for efficient translation initiation biases codon usage at second amino acid position in secretory proteins[J].Nucleic acids research,2007,35(17):5748-5754.
[7] HUANG Y,KOONIN E V,LIPMAN D J,et al.Selection for minimization of translational frameshifting errors as a factor in the evolution of codon usage[J].Nucleic acids research,2009,37(20):6799-6810.
[8] 孫铮,马亮,MURPHY R,等.Wolbachia pipientis wMel 基因组水平上的密码子使用分析[J].中国科学(C辑:生命科学),2009,39(10):948-953.
[9] HIRAOKA Y,KAWAMATA K,HARAGUCHI T,et al.Codon usage bias is correlated with gene expression levels in the fission yeast Schizosaccharomyces pombe[J].Genes to cells,2009,14(4):499-509.
[10] SUBRAMANIAN S.Nearly neutrality and the evolution of codon usage bias in eukaryotic genomes[J].Genetics,2008,178(4):2429-2432.
[11] 黄琼林.高良姜叶绿体基因组测序与特征分析[J].热带作物学报,2021,42(1):1-6.
[12] 原晓龙,李云琴,张劲峰,等.乳油木叶绿体基因组密码子偏好性分析[J].分子植物育种,2020,18(17):5658-5664.
[13] 辛雅萱,董章宏,瞿绍宏,等.杜梨叶绿体基因组密码子偏好性分析[J].河北农业大学学报,2020,43(6):51-59.
[14] 胡晓艳,许艳秋,韩有志,等.酸枣叶绿体基因组密码子使用偏性分析[J].森林与环境学报,2019,39(6):621-628.
[15] 刘文武,蒋晓文,张帅,等.羌活中香豆素类化学成分及其抗氧化活性研究[J].中草药,2019,50(6):1310-1315.
[16] 杨娇.羌活属叶绿体基因组进化及物种界定研究[D].西安:西北大学,2018:5-6.
[17] 阎梦颖.珍稀濒危羌活属药用植物的谱系地理学研究[D].西安:西北大学,2016:5-6.
[18] 郭培,郎拥军,张国桃.羌活化学成分及药理活性研究进展[J].中成药,2019,41(10):2445-2459.
[19] 赵森,邓力华,陈芬.秋茄叶绿体基因组密码子使用偏好性分析[J].森林与环境学报,2020,40(5):534-541.
[20] 吴宪明,吴松锋,任大明,等.密码子偏性的分析方法及相关研究进展[J].遗传,2007,29(4):420-426.
[21] 尚明照,刘方,华金平,等.陆地棉叶绿体基因组密码子使用偏性的分析[J].中国农业科学,2011,44(2):245-253.
[22] WRIGHT F.The‘effective number of codons’used in a gene[J].Gene,1990,87(1):23-29.
[23] SHARP P M,LI W H.The codon Adaptation Index—A measure of directional synonymous codon usage bias,and its potential applications[J].Nucleic Acids Res,1987,15(3):1281-1295.
[24] 陸奇丰,骆文华,黄至欢.两种梧桐叶绿体基因组密码子使用偏性分析[J].广西植物,2020,40(2):173-183.
[25] 原晓龙,康洪梅,王毅.云南蓝果树叶绿体基因组密码子偏好性分析[J].西北林学院学报,2020,35(4):26-31,124.
[26] 尹为治,方正,黄良鸿,等.海南2种龙脑香科植物叶绿体基因组密码子偏好性分析[J].林业调查规划,2020,45(6):19-26.
[27] 吴妙丽,陈世品,陈辉.竹亚科叶绿体基因组的密码子使用偏性分析[J].森林与环境学报,2019,39(1):9-14.
[28] 胡莎莎,罗洪,吴琦,等.苦荞叶绿体基因组密码子偏爱性分析[J].分子植物育种,2016,14(2):309-317.
[29] 沈宗芳,陆添权,张志荣,等.槲蕨属叶绿体基因组密码子偏好性分析[J].广西植物,2021,41(2):266-273.
