西藏色季拉山高寒草甸植被群落坡向分异特征
2022-03-02孙凯歌屈兴乐罗大庆
孙凯歌 ,袁 敏 ,屈兴乐 ,罗大庆 *
(1.西藏农牧学院高原生态研究所,西藏 林芝 860000;2.西藏高原森林生态教育部重点实验室,西藏 林芝 860000;3.西藏林芝高山森林生态系统国家野外科学观测研究站,西藏 林芝 860000)
植物群落是经过环境的长期变化,并通过自身的适应调节而形成的一种维持生态系统稳定和生产力水平的物质基础[1]。生物所组成的群落和生态系统具有多样化、变异程度和复杂性[2],既是对所在环境的响应也是生态学特征的综合表现,能够反映群落的差异性和稳定性[3]。生物量和物种丰富度主要反映草甸群落的生物资源变化和结构功能[4-5],而植物区系是植物界在某个地理和历史环境的综合作用下所演化的结果,也是植物分类的重要依据[6]。
地形是体现时空异质性的外界环境因素之一,它直接影响着植被生理变化和生理周期,间接影响着群落结构及动态变化[7]。特别是在高山草甸区,坡向通过改变太阳辐射强度、风向进而影响温度和降水,使草甸植被呈现一定复杂性[8],通常阳坡多为耐旱植物,阴坡多生长喜阴喜湿植物[9]。近些年已有研究表明,高寒草甸植被群落物种丰富度北坡高于南坡[10-11],生物量阴坡大于阳坡[12-13],但目前东、西坡向对高寒草甸群落特征的影响研究较少,需要进一步探究。
高寒草甸是西藏高原地区的主要植被类型,具有高原地带性和垂直地带性[14],同时发挥着强大的涵养水源和碳汇功能[15]。研究区色季拉山位于藏东南地区,拥有独特的地理和气候条件,林线之上的高寒草甸是该区域垂直带谱中的典型植被类型,草甸植物群落分布复杂,种类齐全[16]。本文通过对西藏色季拉山高寒草甸东、西坡植物组成和区系的探究以及物种多样性和生物量的比较,可为研究区域内不同草甸植物群落保护和利用提供依据,同时对藏东南草地生态系统维持、生产力水平和系统认知具有重要意义。
1 材料和方法
1.1 区域概况
色季拉山位于西藏东南部的林芝县境内,地处喜马拉雅构造带、念唐古拉构造带的东端和横断山脉构造带的交汇处,也是尼洋河流域和帕隆藏布江的分水岭。山体处于藏东南湿润气候和半湿润气候过渡区,冬春少雨,夏秋多雨,年降水量为650~1 130 mm,降雨主要集中在5―9月;年均气温-0.73℃,最暖月(7月)平均气温9.8℃,最冷月(1月)平均气温-13.8℃。色季拉山作为藏东南部典型的山脉体系,植被生长繁茂,种类繁多,垂直谱带明显,其植被沿海拔高度的升高有明显的分布规律。
1.2 试验方法
1.2.1 样地布设及调查
为了解色季拉山高寒草甸植被基本情况,于2019年5月,在生长季来临前,对色季拉山高寒草甸进行踏查,调查草甸植被分布、类型等基本情况。根据踏查情况,以色季拉山顶山脊线为分水岭,分别在其东、西坡两侧草甸植被选择面积大、分布连续和生长良好的草地作为调查与试验样地,样地信息见表1。于2019年7月,在植物生长盛期进行植被群落调查和样品采集。采用典型样地法,在东坡、西坡分别设置10.0 m×10.0 m调查样地3块,在样地中心以及对角线中心设置5个1.0 m×1.0 m样方进行群落调查。使用GPS测定样方的经纬度以及海拔、坡度和坡向等环境因子,同时记录群落特征,并且采集样方内所有出现的植物种带回制作成标本,用于后期物种鉴定。

表1 高寒草甸样地信息Table1 Information of alpine meadow sample plot
1.2.2 植物物种及区系组成鉴定
对所采集的植物标本经过压制、晾干、挂牌、消毒和上台纸等工作后,再对植物标本进行确定到种的鉴定。根据鉴定结果,建立色季拉山高寒草甸区的植物名录,运用Microsoft Excel进行科、属和种的数量分析,分析色季拉山高寒草甸植物物种组成情况。根据吴征镒的《中国种子植物属的分布区类型》[17]、《世界种子植物科的分布区类型系统》及其修订[18-19],查找《西藏植物志》[20],分析确定植物科、属的区系组成。
1.2.3 植物地上、地下生物量测定
生物量采集采取全挖法采集。根据植被调查数据得到的植物种类,在样地内分种以平均大小分别挖取10~20株植株,挖取全部植株带回实验室,采用水冲洗的方式洗去泥土并吸干植株表面水分。将干净的、完整的植株分为地上与地下部分两个部分,置于80℃恒温干燥箱内烘干至恒重,用分析天平称量干重,计算每种植物平均单株干重,结合调查数据中植物多度计算群落生物量。
1.2.4 植物物种多样性计算
根据调查的植物群落数据,包括物种在群落中的多度、盖度、频度、生殖高度和营养高度等信息,计算各物种在群落中的重要值,并根据重要值计算群落多样性指数(丰富度指数、多样性指数和均匀度指数)。
重要值的计算公式为:IV=(Dr+Pr+Hr+Fr+Br)/5
算式中,其中Dr为相对多度,Pr为相对盖度,Hr为相对高度,Fr为相对频度,Br为相对生物量,IV为重要值。
Margalef丰富度指数d=(S-1)/lnN
其中:Pi代表物种的相对重要值,S代表样方内物种数目,N代表样方内所有物种个体数之和。
Pielous均匀度指数J=H′/H′max
式中,H′为实测样方的Shannon-Wiener指数。
1.2.5 数据处理
采用Microsoft Excel 2013统计分析色季拉山高寒草甸物种组成、物种多样性指数和生物量,以东、西坡两个样地数据计算算数平均值。用SPSS 25.0软件进行差异性分析,采用Origin 2018绘图。
2 结果与分析
2.1 高寒草甸植物群落的物种组成
在色季拉山高寒草甸区域共调查到种子植物53种,分别隶属于22科37属(表2)。群落中包含5种以上的仅菊科(Compositae)一科,共10种,占总种数的18.87%;包含3~4种的有禾本科(Gramineae)、玄参科(Scrophulariaceae)、报春花科(Primulaceae)、虎耳草科(Saxifragaceae)、龙胆科(Gentianaceae)、蔷薇科(Rosaceae)和莎草科(Cyperaceae)共7科23种,占总种数的43.40%;含有2种及以下植物的有兰科(Orchidaceae)、毛茛科(Ranunculaceae)和灯心草科(Juncaceae)等共14科20种,占植物总种数的37.74%。含5属以上的仅有菊科,共有7个属,占总属数的18.92%,包含3~4属的有禾本科、玄参科,占总属数的18.92%;含有2属及以下的有报春花科、虎耳草科等共19科23属,占总属数的62.16%。结合种与属的分析,研究区域内菊科植物占优势,禾本科、玄参科植物为重要组成科,虎耳草科、兰科等植物丰富了该区域的植被组成。区域单种科、属代表了该区域物种发展的两个阶段,一方面是处于新产生物种的阶段,还未形成新的种、属;另一方面是处于物种的消亡阶段,该区域中科内的种、属逐渐消失,留下残遗种类,色季拉山高寒草甸区域内物种科、属存在单种的数目较多,占有一定的比例(15.12%),与物种受环境影响有关,区域特征较为明显,独特性突出,是植物对高山高寒气候环境的适应,也是进化过程的反映。

表2 种子植物组成情况Table 2 Composition of seed plants
色季拉山东、西坡群落的物种组成的差异由重要值(IV>0.05)分析可知,见图1和图2,东坡高寒草甸区优势种植物为丛生荽叶委陵菜(Potentilla coriandrifoliaD.D)和圆穗蓼(Polygonum macrophyllumD.D),亚优势种为高山嵩草(Kobresia pygmaeaC.B.Clarke)、硬 叶 风 毛 菊(Saussurea ciliaris.)、尼泊尔香青(Anaphalis nepalensisHand.-Mazz.)和 狮 牙 草 状 风 毛 菊(Saussurea leontodontoides);东坡草甸区的偶见种为垫状点地梅(Androsace tapeteMaxim.)、银叶委陵菜(Potentilla leuconota)、刺参(Oplopanax elatusNakai.)和裂叶蓝钟花(Cyananthus lobatusWall.ex Benth.),东坡群落为丛生荽叶委陵菜+圆穗蓼群丛。西坡高寒草甸区的优势种为珠芽蓼(Polygonum viviparumL.),亚优势种为大花嵩草(Kobresia macranthaBocklr.)和丛生荽叶委陵菜(Potentilla coriandrifoliaD.D);西坡草甸区的伴生种为白心球花报春(Primula atrodentataW.W.Smith),偶见种为倒锥花龙胆(Gentiana obconicaT.N.Ho),西坡群落为珠芽蓼+大花嵩草群丛。
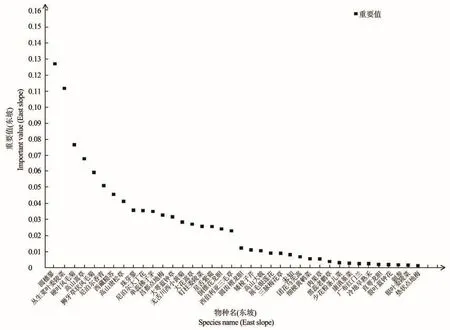
图1 东坡群落物种重要值Figure 1 Important value of species in east slope
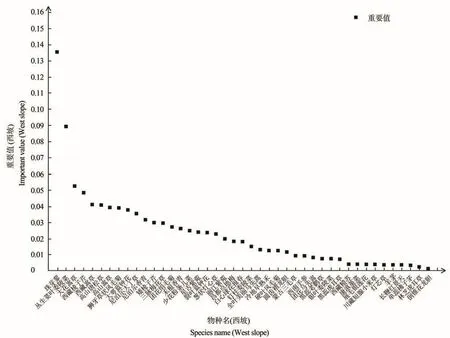
图2 西坡群落物种重要值Figure 2 Important value of species in west slope
2.2 高寒草甸植物群落的区系组成
2.2.1 科的区系组成
根据吴征镒的方法[18-19],将世界种子植物的科划分为18个大分布区类型。照此方法,将研究区域内22个植物科的分布划分为4个类型及1个变型(见表3),其中世界分布型最具优势,共有18个科,占研究区总科数的81.82%,包含科有菊科(Compositae)、伞 形 科(Umbelliferae)、禾 本 科(Gramineae)、蓼 科(Polygonaceae)和 龙 胆 科(Gentianaceae)等;泛热带分布型有1个科,为大戟科(Euphorbiaceae),占研究区总科数的4.55%;东亚(热带、亚热带)及热带南美间断型有1个科,为五加科(Araliaceae),占研究区总科数的4.55%;北温带分布型有1科,为百合科(Liliaceae),占研究区总科数的4.55%;北温带和南温带间断分布型有1科,为牻牛儿苗科(Geraniaceae),占研究区总科数的4.55%。从表中分析可知,色季拉山高寒草甸植物科多具有世界广布特征,没有地区特有科分布。

表3 种子植物科的区系分布类型Table 3 Floristic distribution types of seed plant families
2.2.2 属的区系组成
按照吴征镒关于中国种子植物属划分的15个分布区类型[17],研究区域内植物属的分布可划分8个类型及3个变型(见表4),其中北温带分布型最具优势,共18个属,占研究区总属数的48.65%,包含属有点地梅属(AndrosaceL.)、紫菀属(AsterL.)、羊茅属(FestucaL.)、马先蒿属(PedicularisL.)、委陵菜属(Po⁃tentillaL.)和报春花属(PrimulaL.)等;世界分布型共8个属,占研究区总属数的21.62%,包含有早熟禾属(PoaL.)、蓼属(PolygonumL.)、龙胆属(Gentiana(Tourn.)L)和堇菜属(ViolaL.)等;其次是旧世界温带分布型,共有3个属,占研究区总属数的8.11%,包含有糙苏属(PhlomisL.)、棱子芹属(Pleurospermum camtschaticumHoffm.)和匹菊属(PyrethrumZinn.);泛热带分布型有1个属,为大戟属(EuphorbiaLinn.),占研究区总属数的2.70%;热带亚洲至热带非洲分布型有1个属,为大丁草属(GerberaCass.),占研究区总属数的2.70%;北极-高山分布型有1个属,为红景天属(RhodiolaL.),占研究区总属数的2.70%;北温带和南温带间断分布型有2个属,包含唐松草属(Thalic⁃trumL.)和三毛草属(TrisetumPers.),占研究区总属数的5.41%;东亚及北美洲间断分布型有2个属,包含刺参属(OplopanaxMiq.)和粉条儿菜属(AletrisL.),占研究区总属数的5.41%;地中海区、西亚至中亚分布型有1个数,为菊苣属(CichoriumL.),占研究区总属数的2.70%;东亚分布型有1个属,为黄鹌菜属(YoungiaCass.),占研究区总属数的2.70%;中国-喜马拉雅分布型有2个属,包含蓝钟花属(CyananthusWall.ex Benth.)和肉 果 草属(LanceaHook.f.et Thoms.),占研究区总属数的5.41%。从植物属的分布来看,该区植物具有明显的温带性质,并且以北温带成分为主,且无中国特有属分布。

表4 种子植物属的区系分布区类型Table 4 Floristic distribution types of spermatophyte genera
2.3 高寒草甸植物群落物种多样性特征
物种多样性是丰富度和均匀度的综合体现,能够反映群落物种丰富程度和个体在群落中的分布状况。通过分析,色季拉山高寒草甸群落Shannon-Wiener指数为3.23,Margalef指数为4.79,Pielou指数为0.90。由表5可见,色季拉山东、西坡Shannon-Wiener指数存在显著差异(P<0.05),西坡(3.330)>东坡(3.137),说明西坡群落有较多植物种共同生长,且每一物种的数目分布相对比较均匀。Margalef指数表现为西坡(5.270)>东坡(4.326);Pielou指数表现为西坡(0.911)>东坡(0.893),表明色季拉山高寒草甸西坡群落中全部物种个体数目分布较东坡更均衡,物种多样性高,物种数量更多。
2.4 高寒草甸群落生物量特征
由表5所示,色季拉山草甸群落总生物量为443.24~626.19 g/m2;群落地上生物量为153.12~253.03 g/m2;群落地下生物量为209.12~373.16 g/m2,总生物量东坡高于西坡41.28%(P<0.05)。从草甸群落东、西坡地上、地下生物量比较来看,东坡地上生物量为253.03 g/m2,西坡地上生物量153.12 g/m2,东坡较西坡高65.25%(P<0.05),差异显著;东坡地下生物量为373.16 g/m2,西坡地下生物量为290.12 g/m2,东坡比西坡高28.62%,但未达到显著差异(P>0.05),东坡与西坡总体表现为地下部分的生物量高于地上部分。

表5 高寒草甸群落特征指数及方差分析Table 5 Community characteristic index and variance analysis of alpine meadow
从群落物种生物量总体来看,图3-4可见,丛生荽叶委陵菜既是东、西坡草甸区生物量最高的物种,也分别是地上、地下生物量上最具优势的物种。而东坡狮牙草状凤毛菊、圆穗蓼、硬叶凤毛菊在地上、地下生物量上优势显著,西坡则是珠芽蓼和狮牙草状凤毛菊占主要优势。

图3 东坡群落物种生物量Figure 3 Species biomass of the community on the east slope
通过对群落优势种生物量进行比较,由表6可见,色季拉草甸东坡优势种生物量占群落总生物量的50.87%,西坡优势种生物量仅占16.43%,表明东坡优势种的生物量在群落中的贡献度远高于西坡。通过根冠比值的比较,东坡根冠比为1.47,西坡根冠比为1.89,东、西坡植物地下生物量明显高于地上生物量。而东坡优势种的根冠比是2.13,非优势种根冠比是1.04;西坡优势种的根冠比是2.16,非优势种根冠比是1.85,东、西坡优势种地下生物量明显高于地上生物量,但东坡非优势种地上、地下生物量接近,而西坡非优势种地下生物量明显高于地上生物量。
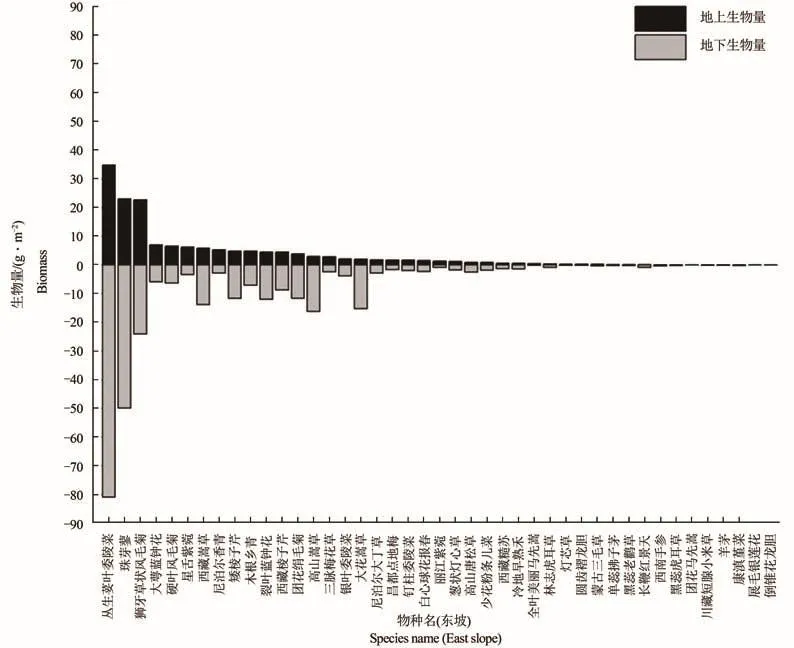
图4 西坡群落物种生物量Figure 4 Species biomass of community on west slope

表6 优势种生物量贡献值Table 6 Biomass contribution value of dominant species
3 讨论与结论
色季拉山东、西坡高寒草甸区共调查到种子植物22科,37属,53个种,分布在研究区的优势科多数植株矮小、根系发达,同时多具毛被的形态特征,能够抵御冻害和强烈辐射,显然已经成为该区自然条件下的适生类群。在研究区分布2种以下的科共有14个,占总科数的63.63%,然而种数却仅占总种数的37.74%,说明该区的恶劣环境使多数科的植物分布到该区的种类数量受到了限制。从科的分布类型来看,菊科、伞形科、禾本科和蓼科等18个大科是世界分布,就其科的层次来说,对于显示一个地区的植物区系特点的意义不大,但在色季拉山高寒地带的代表属却多以北温带分布为主,如点地梅属、紫菀属、羊茅属和马先蒿属等。色季拉山高寒草甸植物在属的分布区类型上成分较复杂,但其地理成分却是比较单一,主要以北温带分布为主,具有明显的温带特征,在分布上由世界分布向北温带分布过渡。可能由于该区域是世界陆地垂直地貌落差最大的地带,植物区系过渡性应该与青藏高原抬升和气候环境变迁有关。
植物群落多样性依赖于植物物种在一定区域内分布的疏密程度,直接或间接的体现群落的稳定性、生境差异等。色季拉山高寒草甸物种数量及多样性均高于藏北草甸[15,21],由于地处藏东南半干旱半湿润地区,年平均温度与藏北草甸区相似,但降水却更丰富,所以水分是影响色季拉高寒草甸植物生长的主要因素[22],因此植物生长繁育更优。而物种多样性表示一定区域内所含物种数目的多少以及物种分布的均匀程度[23]。其中Shannon-Wiener指数和Pielou指数存在极显著差异(P<0.01),指数值均表现为西坡>东坡,西坡植被的物种数量和均匀程度明显高于东坡,表明西坡的气候和土壤条件利于更多高寒草甸植被物种的生长繁育,而西坡植被均匀程度更高表明群落内部间的生态关系排斥性更强[24-25];Margalef指数差异显著,表明西坡植被物种更丰富的同时,其物种数量也高于东坡,这与祁连山和青海尖扎县高寒草甸区的相关研究的结果相似[11,13]。同时,相关研究表明,通常西坡物种多样性更高,群落结构也更复杂,多物种共存使群落物种发展更均衡,稳定的群落结构也能更加适应外部环境的变化[10,26]。
植物群落生产力水平是生态系统功能的主要表现形式,而生物量是反映能流、物质流和描述第一性生产力的重要指标[27]。色季拉山东、西坡草甸群落地下生物量均高于地上,一般认为,植物会把更多生物量分配到获取资源限制最严重的器官上[28-30],由于色季拉山高寒草甸区全年气温和土壤温度低,辐射强,水分蒸发快,所以该区域草甸植被利用雨热同期的气候条件,增强地下根系的发育来获取土壤中的水分和养分,以此应对气候环境对根系养分吸收的限制。同时,东坡地上生物量高于西坡65.25%,地下生物量高于西坡28.62%,东、西坡地上生物量存在显著差异。研究表明在同样的温度条件下,地下生物量与土壤含水量有显著的正相关关系[31],而色季拉东坡高寒草甸区为山地迎风坡,降水多于西坡,土壤含水量更高,因此东坡将更多生物量分配到地上部分。本次研究还发现东坡优势种的生物量对总生物量的贡献约为50%,表明东坡优势种的生物量是总生物量的主要贡献者,而西坡优势种生物量对总生物量的贡献值仅为16.43%,表明西坡优势种对于群落生物量的贡献较低。除此之外,东、西坡优势种的地下生物量远高于地上生物量,说明优势种把更多生物量分配到根系,这种分配方式使它们能更好地适应高寒气候。
综上所述,色季拉山东、西坡高寒草甸植物虽然分布区类型相对复杂,但地理成分单一,具有明显温带特征,与色季拉山海拔高差和藏东南的特殊气候有关;色季拉山高寒草甸物种多样性丰富,以珠芽蓼+大花嵩草群丛为优势种群落的西坡物种多样性优于以丛生荽叶委陵菜+圆穗蓼群丛为主的东坡植被群落;研究区地下生物量高于地上生物量,同时东坡生物量高于西坡,且东坡优势种生物量对总生物量的贡献值更高。因此,对植物群落结构、物种多样性和生物量开展监测研究,可为色季拉山高寒草甸植被群落可持续发展和生产力水平提供科学依据。
