Rhizosphere microbiomes can regulate plant drought tolerance
2022-03-02MehtabMuhammadASLAMEyaliraOKALAishaLawanIDRISZhangQIANWeifengXUJosephKARANJAShabirWANIandWeiYUAN
Mehtab Muhammad ASLAMEyalira J.OKALAisha Lawan IDRISZhang QIAN∗Weifeng XU∗Joseph K.KARANJAShabir H.WANI and Wei YUAN∗
1College of Agriculture,Yangzhou University,Yangzhou 225009(China)
2Joint International Research Laboratory of Water and Nutrient in Crops and College of Life Sciences,Center for Plant Water-Use and Nutrition Regulation and College of Resources and Environment,Fujian Agriculture and Forestry University,Fuzhou 350002(China)
3Juncao Research Center,College of Life Sciences,Fujian Agriculture and Forestry University,Fuzhou 350002(China)
4State Key Laboratory of Ecological Pest Control for Fujian and Taiwan Crops&Key Laboratory of Biopesticide and Chemical Biology of Ministry of Education,College of Life Sciences,Fujian Agriculture and Forestry University,Fuzhou 350002(China)
5Mountain Research Centre for Field Crops Khudwani,Sher-e-Kashmir University of Agricultural Sciences and Technology,Kashmir 192102(India)
ABSTRACT Beneficial root-associated rhizospheric microbes play a key role in maintaining host plant growth and can potentially allow drought-resilient crop production.The complex interaction of root-associated microbes mainly depends on soil type,plant genotype,and soil moisture.However,drought is the most devastating environmental stress that strongly reduces soil biota and can restrict plant growth and yield.In this review,we discussed our mechanistic understanding of drought and microbial response traits.Additionally,we highlighted the role of beneficial microbes and plant-derived metabolites in alleviating drought stress and improving crop growth.We proposed that future research might focus on evaluating the dynamics of root-beneficial microbes under field drought conditions.The integrative use of ecology,microbial,and molecular approaches may serve as a promising strategy to produce more drought-resilient and sustainable crops.
Key Words:biota,drought stress,metabolites,phytohormone,plant growth,root-microbe association
INTRODUCTION
Among natural disasters,drought is one of the most influential abiotic stresses that severely affects crop productivity and yield,thereby threatening global food security(Naylor and Coleman-Derr,2018;Bahaduret al.,2019).It is estimated that one-third of the world’s arable land is affected by water shortage with crop production limited by drought stress(Naseemet al.,2018).However,provision of food to the ever-growing population relies on an improved crop production under drought stress.According to environmental models,the intensity and frequency of drought in the coming years have been predicted to increase due to global climate change(Dai,2013).The intensity and duration of drought substantially threaten crop yield globally.With increasing drought episodes,understanding plant physiological responses to drought is crucial for sustaining the productivity of agricultural ecosystems(Guptaet al.,2020).Despite immense researches aiming at understanding drought-resilient crop production,particularly complex feedback between root-microbial interaction and how these interactions are influenced over time under drought have been largely ignored.Indeed,roots are key to uptake of water and nutrients from soil,and exhibit a range of root developmental modification traits to adapt to several abiotic stresses.Various approaches have been tried to increase drought tolerance in plants,leading to higher crop yield(Aliet al.,2017,2020;Buddolla,2019;Ullahet al.,2019).Plant and microbial interactions are crucial for maintaining terrestrial ecosystem and their responses to rapidly fluctuating environment.Rhizospheric microbiomes protect plants from pathogens(Chialvaet al.,2018),and promote nutrient uptake efficiency(Hunter,2016).Additionally,beneficial microbiomes may provoke plant performance and phytohormone-mediated abiotic stress resistance(Spaepen,2015;Egamberdievaet al.,2017).Recent studies suggest that microbiomes can influence plant morphological traits(e.g.,leaves,shoot,secondary root,root length,and biomass)(Kumaret al.,2018;Steidingeret al.,2019)and plant phenology(e.g.,flowering time)(Luet al.,2018),suggesting great contribution towards plant community dynamics and mediate plant response to environment(Crawfordet al.,2019;Delavauxet al.,2019;Steidingeret al.,2019).Massive efforts have been made to understand plant-microbe interaction;however,the mechanisms controlling rhizospheric microbiome assembly,composition,and structure under drought are largely unknown.To understand the molecular regulatory mechanism involved in mitigating plant responses to abiotic stress,researchers often identify an important section of missing variance related to the ecology,the“environment by genotype”interactions(Naylor and Coleman-Derr,2018).Hence,exploring the relationships or interrelationships between plants,rhizosphere microbes,and the environment would be of great importance in understanding their contributions and responses to drought stress tolerance.Soil microbial biota represents the largest reservoir of biological diversity globally(Buéeet al.,2009).The complex association between root and microbes,considered the plant’s second genome,is crucial for maintaining plant health(Berendsenet al.,2012).Root and microbial interaction is regularly defined as trench warfare,that may significantly inhibit the infection of pathogenic microbes(Berendsenet al.,2012).The rhizosphere is the area around the root influenced by root growth processes(Zhang Yet al.,2020)and all of the microbial population existing in the rhizosphere known as rhizobiome(Lundberget al.,2012;Del Carmen Orozco-Mosquedaet al.,2018).Rhizosphere soil contains almost 1011microbial cells g−1root and>30 000 prokaryotic organisms.The whole genome of rhizospheric microbiome is much greater than that of the plant(Mendeset al.,2011).
Mycorrhizal symbiotic associations between plant root and rhizospheric microbes establish various distinct mechanisms that are beneficial to both the host plant and microbes(Aslamet al.,2019).Microbes can stimulate root growth and thus soil exploration and enhance nutrient acquisition from soil by decomposing plant organic matter.In return,microbes gain 20%—25%of photosynthetic carbon(C)derived from leaf,root litter,soil dead organic matter,and root exudate to carry out their metabolic activities.Plant roots continuously exude a wide array of plant-derived metabolites(e.g.,mucilage,organic acids,free ion radicals,oxygen,water,enzymes,sugars,and phenols)known as root exudates into the rhizosphere,while the magnitude of root exudation relies on plant genotype,soil type,age,and nutrient availability.Recent studies recommended that root exudation plays a critical role in the screening of favorable rhizospheric microbiomes by modulating root exudation patterns,which give new insights into improving plant growth and yield(Williams and De Vries,2020).Mutual interaction of both organisms exudes chemical signals and exchanges nutrients that determine plant growth and maintain soil moisture and structure.These beneficial microbes,such as mycorrhizae,interact with plant in the rhizosphere and promote plant growth by colonizing plant roots(Berendsenet al.,2012).Several studies had reported that drought stress greatly influences plant growth and rhizospheric microbial activity,resulting in alterations of nitrogen(N)and C cycling processes(Gordonet al.,2008;Sanaullahet al.,2011;De Vrieset al.,2016,2018;De Nijset al.,2019).Although studies on rhizospheric microbes and their roles in improving plant fitness and growth are available(Ahmedet al.,2020;Juet al.,2020;Teijeiroet al.,2020),there are fewer studies on the beneficial mechanisms of root microbiomes under drought stress.For example,Aalipouret al.(2020)inoculatedArizona cypresswith arbuscular mycorrhiza fungi(AMF)and rhizobacteria under well-watered(WW)and severe water deficiency(SWD)conditions and showed that AMF-inoculated plant growth was significantly better than non-inoculated plant growth under SWD.Interestingly,the symbiotic interaction between rhizobacteria and AMF co-inoculation can alleviate drought stress and reduce water stress damage.Similarly,Zhanget al.(2020)demonstrated that orange inoculated with AMF under WW and drought stress conditions showed increased polyamine(PA)expression to protect cells from damage and improve drought tolerance.This suggests that the inoculation of beneficial microbiomes may provide superior and better adapted plant varieties under stressed condition.
Root exudates are considered as key determinant for maintaining rhizobiome structure and exerts significant effect on plant performance and survival under stress condition.Recently,various studies on root exudates have revealed their positive correlations with soil microbial community(Gargallo-Garrigaet al.,2018;Guyonnetet al.,2018;De Nijset al.,2019),while the interactive responses of root exudates and rhizospheric microbes to drought remain unidentified.It is proposed that root exudation is directly involved in selecting beneficial rhizosphere microbiomes.Altering root exudation may help to select beneficial microbiomes under drought stress and could help in developing new strategies to improve crop growth,particularly yield(Williams and De Vries,2020).
This review highlights the important plant and microbial traits affected by water shortage,particularly the role of beneficial rhizospheric microbiomes in the improvement of plant drought tolerance.Additionally,drought-induced modifications in plant secondary metabolite production and phytohormone synthesis are explored.
EFFECT OF DROUGHT ON PLANT AND ASSOCIATED COMMUNITIES
Drought effect on plant traits
Drought stress influences multiple plant physiological and biochemical processes(Fig.1),including photosynthesis,respiration,uptake and translocation of ions,and carbohydrate and nutrient metabolism or their translocation to shoot,thereby disturbing hormonal and nutritional homeostasis in plant and reducing plant growth(Naseemet al.,2018).Plant photosynthetic rate is detrimentally affected during drought stress due to decreased chlorophyll content,fluorescence,quantum yield,and decreased stomatal conductance lowering the photosynthetic capacity(Naylor and Coleman-Derr,2018).Additionally,drought stress can cause decreased turgor pressure,limited gaseous exchange,reduced CO2assimilation,impaired photosynthetic apparatus(mainly photosystems I and II),and enhanced metabolite fluxes(Zargaret al.,2017).However,the major determinant of reduced photosynthesis under drought stress is stomatal limitation,lowering both foliar photosynthetic rate and internal CO2concentration,further inhibiting total photosynthetic metabolism.In addition,various nonstomatal effects,including photophosphorylation,ribulose-1,5-bisphosphate(RuBP)regeneration,rubisco activity,and ATP synthesis,are also attributed to stomatal closure under drought condition(Reddyet al.,2004).Drought inhibits photosynthesis by limiting C intake and potential energy reserves,which makes the adaptive strategy energetically intensive.The quality and quantity of root exudates found under drought condition are different to those under normal condition(Williams and De Vries,2020),indicating that root exudates are one of the strategies that could be highly intensive under drought.
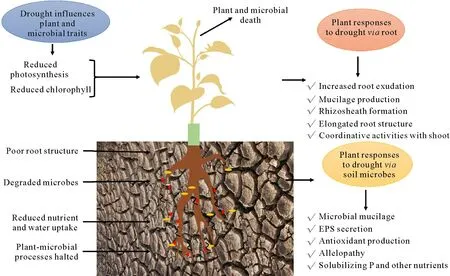
Fig.1 Plant and microbial traits influenced by drought and their responses to drought stress.Upon drought exposure,several plant physiological and microbe-associated mechanisms are downregulated,including reduced photosynthesis and degraded beneficial root microbes.To overcome this stress,plants respond via altering root structure or interacting with drought-resistant beneficial rhizospheric microbes.EPS=exopolysaccharides.
An imbalance between root water uptake and shoot transpiration can cause loss of shoot turgor,thereby inhibiting plant growth and development.Reactive oxygen species(ROS)are highly reactive chemical molecules existing in four forms(1O2,O2,H2O2,and HO)that originate as byproducts of natural oxygen metabolism and play critical roles in plant acclimation process against several abiotic stresses.Reactive oxygen species are induced by drought,resulting in oxidative damages to protein synthesis carbohydrates,and lipid metabolisms,which subsequently lead to membrane damage and necrosis in plant tissues(Bahaduret al.,2019;Duttaet al.,2019).Upon oxidative stress,drought enhances soil O2,increases soil heterogeneity,restricts nutrient availability and mobility,and often imposes a strong reduction in soil microbial biota.It has been described that ROS,such as H2O2,are emitted from root surface into rhizosphere(Zouet al.,2015).H2O2,however,aids in lowering the oxidative burst of host plant trifoliate orange(Poncirus trifoliate)by arbuscular mycorrhiza(AM)as demonstrated by Zouet al.(2015)and Huanget al.(2017).For plant-microbial communication,root exudates act as important pathways that provide photosynthetic C for microbial growth,in addition to mediating communication between plants and microbes through phytohormones and signaling molecules(De Vrieset al.,2020).Future studies must focus on the complex feedback of plant and microbial responses to drought,which may offer great potential for producing drought-resilient crops.
Drought effect on rhizospheric microbiomes
Rhizosphere is a natural ecosystem that hosts numerous kinds of microorganisms,including bacteria,archaea,actinobacteria,algae,fungi,and protozoa(De Vries and Wallenstein,2017).Various compounds,including carbohydrates,amino acids,and organic acids,are exuded by roots into soil through diffusion,ion channels,and vesicular transport.In addition to changing soil chemistry,the compounds also supply nutrients to the microbes in rhizosphere(Lareenet al.,2016),hence serving as a distinctive competitive environment for microbes,including N-fixing bacteria(Liuet al.,2019).The rhizosphere regions of 20 selected tropical gardens showed a significant diversity in bacterial communities.The microbial number of the various rhizospheres ranged from 6.25×107to 7.9×106cells g−1soil,which is incomparable to that of the unplanted(1.8×107cells g−1soil)bulk soil communities.This had been attributed to the abundance of nutrients resulting from root exudation(Reubenet al.,2013).However,drought stress modulates microbial abundance and activities in rhizosphere(Naseemet al.,2018).Reubenet al.(2013)demonstrated that 60%of selected plants showed a 60%—90%reduction in bacterial counts of the rhizosphere soil samples following exposure to drought stress.This implies that drought could have severe effects on rhizosphere microbiomes,and special attention is required to maintain the ecological niches of microbes within rhizosphere.
Carbon sources exuded by roots are utilized for bacterial growth.Composition and abundance of bacterial community alter under drought stress(Fig.1),and rely not only on the direct effect of drought but also on the variations in C availability resulting from changes in root exudation patterns reflecting plant stress(Reubenet al.,2013).It has been hypothesized that drought diverts the translocation of C from shoot to root due to target protein channel closure and shift of C towards production of osmolytes and storage compounds by plants.As such,bacteria groups that largely rely on plant C reduce or die,and remaining bacterial activity is limited to certain areas of moisture,such as soil pores,further restricting their ability to interact with plants.However,when radiolabeled C was supplied to plants,less C appeared in soil bacteria under drought stress than under WW condition(Naylor and Coleman-Derr,2018).
Drought-adaptive strategies,such as improving the uptake of water by enhancing root system development,reducing transpiration loss or water storage in plant tissues,and recovering from drought by osmotic adjustments or the maintenance of cell wall elasticity may result in changes in the belowground C input.These may affect the functional structure and activity of the microbial community in rhizosphere(Sanaullahet al.,2011).Arbuscular mycorrhiza fungi are widely known as soil-born symbiont fungi for upgrading soil structure and their capacity to enhance water and nutrient uptake from rhizosphere,particularly under abiotic stress.As rhizospheric microbiomes,they produce spores and hyphae network in rhizosphere to connect with plant roots,which significantly increases the access of roots to enlarged soil area,resulting in improved plant growth(Bowleset al.,2018).They not only improve nutrient and water acquisition to overcome the drought effect,but also ameliorate stomatal regulation.They favor enhanced tolerance-related responses to drought stress,subsequently improving plant growth and crop production.Consequently,drought directly affects AMF development cycle,which adversely affects spore germination,colonization capacity,sporulation,and extra-radical hyphal elongation,ultimately lowering AMF community in soil,which reduces soil moisture and limits nutrient mobility(Bahaduret al.,2019).
RESPONSIVE MECHANISMS OF PLANTS TO DROUGHT STRESS
Plant responses to drought via roots
Rhizosheath.Drought tolerance is one of the main strategies employed by plants to withstand drought by modifying structural and morphological plasticity of root system(Ullahet al.,2019;Guptaet al.,2020).These architectural changes are attributed to synchronized root apical meristem development and differentiation(Guptaet al.,2020).Rhizosheath serves as a mechanism that improves drought tolerance and protects the roots under arid conditions(Brownet al.,2017).It is the layer of soil surrounding the root adhering upon excavation of root system.Rhizosheath is formed as a result of soil particles that attach to the root hairs and mucilage secreted by roots or microbes.Preferably,it is formed in drier soil where the adhesive capacity of the mucilage and root hair growth are enhanced and stimulated,thereby improving the firmness of the rhizosheath(Zhang Yet al.,2020).Rhizosheath is effective in relieving such stresses because it helps to sustain good contact between root and soil,which is necessary for nutrient and water acquisition(Brownet al.,2017).This has been measured by studying the regulation of root hair growth and rhizosheath size in response to the decrease in soil moisture.Indeed,the formation of rhizosheath may represent a significant trait that can improve drought tolerance since the soil moisture level of 10%—14%(weight/weight)stimulates higher rhizosheath production and greater root hair density and length as compared to other soil moisture levels,which was validated by measurement of the expression of root hair-related genes(Liuet al.,2020).
Root exudates.Root exudates are a medium of communication between plants and rhizospheric microbes that are used by plants to encourage and recruit microorganisms with plant-beneficial traits during drought(Ullrichet al.,2018;Williams and De Vries,2020).Plant and microbe association has the potential to influence plant tolerance and survival during severe abiotic stress(De Vrieset al.,2019).It has been found that the rate of root exudation and root exudate composition in plants is adjusted to withstand drought stress.This is achieved by inducing higher soil respiration and increasing microbial activity after drought and is likely to enable plant regrowthviaincreasing nutrient mineralization.Root exudates significantly enhance the colonization of roots with beneficial rhizosphere microbiome to compete with drought stress and stimulate plant growth.It is reported that plants exude more organic acids upon drought that play critical roles in phosphorus(P)mobilization,thus improving the capability of nutrient release and uptake from rhizosphere(Songet al.,2012).
Deep root system.Under drought conditions,an enlarged root system architecture facilitates efficient water and nutrient uptake,higher crop yield,and improved drought tolerance(Fig.1).Additionally,small diameters of roots help plants to enhance their hydraulic conductivity by increasing root surface area with soil moisture,as well as increasing the soil volume that can be explored for capturing water(Ullahet al.,2019).Plants with complex rhizomatous roots exhibit flexible responses to episodic water shortage and the development of fibrous roots on the top soil dedicated to maximizing water requirement(De Vrieset al.,2019).
Coordinative activities with shoot.The optimal distribution of resources and biomass is a functional equilibrium exacted by the coordinative activities of root and shoot,which are altered under changing environmental conditions(Sanaullahet al.,2011).Plants undergo various morphoanatomical changes to help them manage drought stress,including leaf size reduction to avoid transpiration,root expansion to absorb moisture from soil,and stem elongation to avoid water loss(Zargaret al.,2017).
Role of plant metabolites in reducing drought stress
Stomatal closure is one of the most primitive plant response to drought,which slows down cell division and elongation,resulting in accumulation of metabolites.The affected plants downregulate metabolic activities owing to alterations in physiological and biochemical processes,that initiate the mobilization of special metabolites in plant cells.The production of unique primary and secondary metabolites in drought-stressed plants is important for the synthesis of protective compounds which are necessary for osmotic adjustment(Aroca,2012;Blum,2017).Osmotic adjustment enables plants to acclimatize to water-deficient conditions by generating increased numbers of osmotically active molecules in plant cells.High solute contents increase the negative osmotic potential,which results in improved cell hydration and maintenance of turgor pressure in active plant cells.Several studies have reported that proline accumulation in newly grown leaves is higher compared to older ones in various plants,such as Arabidopsis(Sperdouli and Moustakas,2014),soybean(Fukutoku and Yamada,1984),cowpea(Zegaouiet al.,2017),and alfalfa(Girousseet al.,1996),suggesting plant responses to developmental drought stress control.A previous study indicate that severe drought induces increased CO2concentration leading to stomatal closure while it was not influenced by moderate water stress(Meidner and Mansfield,1968;Hsiao,1973;Boyeret al.,2008;Gaoet al.,2020).Drought stress therefore induces turgor loss that downregulates photosynthetic metabolism and stimulates multiple cellular responses related to plant physiological,biochemical,and molecular mechanisms(Grandaet al.,2014).Several studies on drought tolerance in plants suggest a strong link between changes in photosynthetic C metabolism and the accumulation level of primary metabolites and other osmoprotectants that are pivotal for stress tolerance.
Normally,primary metabolites,such as amino acids,lactic acid,and carbohydrates,are useful in maintaining plant survival and growth(Fig.2).Drought is,however,known to induce oxidative stress in plants owing to the production of ROS,including singlet oxygen(1O2),superoxide(),hydroxyl radicals(·OH),and hydrogen peroxide(H2O2).Presence of ROS in cells prompts modifications in osmotic chemicals,leading to the production of both primary and secondary metabolites to strengthen plant defense mechanisms(Niinemets,2016).For instance,increased exudation of organic acids plays a critical role in nutrient mobilization,thereby improving nutrient uptake from soil in secondary metabolites,e.g.,flavonoids and anthocyanin caused improved plant drought resistance and plant growth in chili andChenopodium quinoa,respectively,indicating their potential role in plant drought-tolerance mechanism(Akula and Ravishankar,2011).Moreover,plants under osmotic stress produce organic osmoles from sugars and amino acids,such as sucrose,fructose,trehalose,proline,glycine betaine,and quercitol(Perlikowskiet al.,2016).These primary metabolic compounds were reported to act as chaperones,advancing proper cell functioning through the stabilization of enzymes,cell membranes,and subcellular structures(Xoconostle-Cazareset al.,2010;Shahbazyet al.,2020).Although the accumulation of amino acids,particularly proline,is strongly associated with stress tolerance,their role in osmotic adjustment remains a debatable issue,mainly because their contribution to water potential is generally insignificant(Harbet al.,2013).
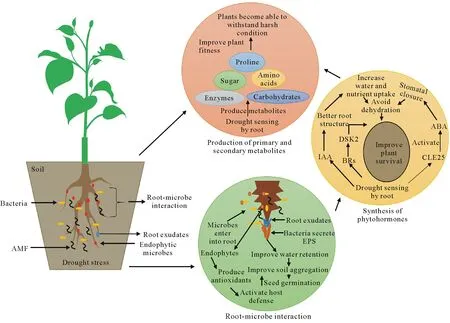
Fig.2 Interactions of rhizospheric microbes with plant roots stimulating plant growth and significantly improving drought tolerance:i)root-microbe interaction,ii)synthesis of phytohormones,and iii)production of primary and secondary metabolites.Root-microbe interaction leads to the secretion of root exudates and exopolysaccharides(EPS),which results in improved water retention around plant root.Drought sensing by plant roots leads to the activation of CLE25 peptide that triggers abscisic acid(ABA)accumulation,inhibiting stomatal opening to avoid dehydration.Similarly,indole acetic acid(IAA)and brassinosteroids(BRs)improve water and nutrient uptake under drought stress.Primary and secondary metabolites also participate in improving plant fitness and yield under stress.AMF=arbuscular mycorrhiza fungi.
Besides primary metabolites,plants also possess defense systems that employ antioxidant enzymes,such as superoxide dismutase(SOD),catalase(CAT),peroxidase(POD),ascorbic acid,and glutathione,as well as a non-enzymatic antioxidant system composed of secondary metabolites(mainly phenols and flavonoids)(Gaoet al.,2020).Several studies have reported an increase in the activities of CAT,SOD,POD,and ascorbate peroxidase when plants are exposed to drought stress(Gusainet al.,2015;Caoet al.,2017).Most drought-tolerant plants have been shown to possess strong scavenging systems that can maintain low levels of ROS and enable plants to prevent membrane lipid peroxidation.Similarly,secondary metabolites act as the first layer of defense to water deficit environment by offering protection against oxidative stress and elimination of ROS by producing phytochemicals such as flavonoids and carotenoids(Akula and Ravishankar,2011).
In a study to assess induced drought resistance inSolanum lycopersicum,Monaet al.(2017)reported increased accumulation of phenols,flavonoids,and proline,which exhibited direct correlation with increased stress tolerance.Elevated levels of phenols and flavonoids protect plants against drought stress by scavenging on free radicals and mediating cell wall formation(Surekhaet al.,2014;Monaet al.,2017).Elevated levels of flavonoids under drought stress have been shown to play a key role in mitigating drought stress in several crops,including wheat(Maet al.,2014),Oryza sativa(Quanet al.,2016),Arabidopsis thaliana(Wanget al.,2019),Capsicum annuum(Rodríguez-Calzadaet al.,2019),andCarthamus tinctorius(Chavoushiet al.,2020).This suggests that the production of metabolites protects plants from drought stress and positively affects plant performance and growth.
Furthermore,flavonoids modulate transportation and signalling of auxins by acting as auxin inhibitors,thus mediating appropriate development during drought.Recent studies have shown that flavonoids,such as flavone,flavanone,flavonol,iso-flavone,and anthocyanin,could reduce indole acetic acid(IAA)-generated radicals during IAA oxidation whereby they inhibit the activity of dioxygenase,and also absorb strong wavelengths that are normally accompanied with high insolation during dry seasons(Brunettiet al.,2018).We surmise that plant metabolites could play a primary role in the successful acclimation of plants to environmental variance during drought stress by interacting with phytohormone pathways.
Amongst secondary metabolites in drought-stressed plants,flavonoids are the most appreciated in exhibiting protective roles due to their special structures consisting of hydroxyl groups and double C bonds.Flavonoids are so diverse and their hydroxyl groups are prone to modifications such as glycosylation,methylation,and prenylation,which are known to counter stress-induced oxidative cell damage(Wanget al.,2018).For instance,Yildiz-Aktaset al.(2009)observed an increase in the production of flavonoids,cinnamic acid,and isoprene derivatives in drought-stressed cotton plants.Flavonoids and cinnamic acid compounds are associated with high abilities of cotton plants to scavenge on ROS,while isoprenes,phenylamines,and tannins were indicated to counter heat-induced stress by acting as antioxidants that quench oxygen radicals in tobacco,carrots,and sorghum,respectively(Tattiniet al.,2014;Isah,2019;Van Doornet al.,2020).According to Maet al.(2014),flavonoid biosynthesis-related genesTaCHS,TaCHI,TaF3H,TaFNS,TaFLS,TaDRF,andTaANSare upregulated in wheat cultivars under drought stress.Furthermore,more than 9 000 different types of flavonoids occur in plants,and the majority of them play a role in the reduction of abiotic stress.
Notably,root exudates under water deficit conditions consist of phenolic compounds,flavonoids,alkaloids,and terpenoids,which act as signaling compounds between plants and symbiotic microorganisms in rhizosphere(Makoi and Ndakidemi,2007).For instance,flavonoids are crucial in root nodulation and are required for the biosynthesis of signaling molecules,referred to as Nod factors,which are detected by plants,thus allowing symbiotic microbes to infect plant roots(Liu and Murray,2016;Mathesius,2019).In essence,flavonoids and phenols from drought-stressed plants stimulate the colonization of symbiotic microbes,such as AM andRhizobia,thereby allowing rhizospheric microbes to colonize roots and provide stress-related benefits to drought-stricken plants.Rhizospheric microbes may respond to drought-related secondary metabolite in plants by secreting useful molecules such as choline or polyamines,which mitigate osmotic stress and promote root growth.Apparently,the primary and secondary metabolites secreted by plants may vary depending on plant genotype and species,development stage,and drought intensity.Future studies may be needed to provide more detailed metabolomics profiles,particularly in plants that are well known to thrive in dry areas.This could provide a suitable link between physiological and genetic studies in deciphering target molecules and pathways in improving drought tolerance in plants.
Role of phytohormones in reducing drought stress
Besides regulating growth and development,phytohormones play a key role in plant responses to abiotic stresses such as drought(Waniet al.,2016).During drought stress,the first plant response generated is stomatal closure.Abscisic acid(ABA)is the key phytohormone that initiates stomatal closure when plants are exposed to water deficient conditions(Lindet al.,2015).Water deficiency in plants activates stress response pathways that involve phytohormone signaling and metabolite production(Bailey-Serreset al.,2019).Drought stress usually stimulates the secretion of ABA hormone in various plant organs.Studies have shown that drought stress can induce responses that increase ABA concentration up to 30 times compared to normal levels(Fig.2)(Outlaw,2003).Abscisic acid is synthesized in the phloem and guard cells that regulate plant responses to water deficiency,and its production is more coherent in leaf mesophyll cells owing to their abundant chloroplast stores(McAdam and Brodribb,2018).
During drought stress,ABA levels significantly increase,causing ion efflux and inhibiting sugar uptake through guard cells,which decreases the stomatal pore size.The ABA mode of action starts when it binds to its receptors(RCARs/PYR/PYL)in the guard cells,blocking the activity of protein phosphatase 2Cs(PP2Cs)such as ABA-insensitive 1(AB1)and 2(AB2).The PP2Cs are negative regulators of ABA and their inhibition was indicated to result in the phosphorylation and activation of proteins that manage drought stress in plants(Zhanget al.,2014;Kauret al.,2016).High levels of ABA during drought stress induce the efflux of anions and K+ions,which causes guard cells to lose their turgor pressure,resulting in stomatal closure.The ABA also induces the guard cells to express NADPH oxidase,which mediates ROS production,hence resulting in stomatal closure.In addition,ABA was reported to move from leaves to roots acting as stress messenger(Limet al.,2015).Therefore,the majority of studies focused on improving drought resistance and water-use efficiency have primarily focused on ABA pathways.For example,Parket al.(2015)demonstrated that engineering of ABA receptorPYR1genes can significantly enhance drought resistance in Arabidopsis and tomato.Since stomatal closure as a result of ABA not only affects transpiration but also CO2influx,there is a need for deeper research to elucidate how ABA influences photosynthesis and how plants overcome drought stress to improve their growth and yield.Molecular studies employing gene manipulation strategies that target the ABA pathway could be employed to enhance ABA-induced responses in crops that are less tolerant to drought stress,thereby promoting food security.
Several studies have reported that microorganisms,such asAzospirillum brasilense,A.lipoferum,Bacillus subtilis,Burkholderia phytofirmans,Achromobactersp.,B.licheniformis,andPseudomonas putida,can alleviate drought effects in plants through the production of ABA(Cohenet al.,2008;Zhang H Met al.,2008;Cohenet al.,2009).In a study on how endophytic bacterial strains affect ABA and jasmonic acid in plants,Forchettiet al.(2007)indicated that strains belonging toAchromobacter xiloxidansandB.pumiluscould increase ABA production up to seven folds inHelianthus annusunder drought stress.Cohenet al.(2015)reportedA.brasilenseto increase ABA level inA.thaliana,resulting in higher plant biomass,increased lateral roots,activated photosynthetic pigments,elevated proline content,and decreased water loss.Inoculation ofA.thalianawith bacteria was shown to improve seed yield,reduce stomatal conductance,and improve plant survival and performance under drought.Furthermore,Salomonet al.(2014)showed thatB.licheniformisandP.fluorescensremarkably enhanced ABA level and defense-related terpenes in grape vine.Similarly,B.licheniformisandP.fluorescensincreased ABA content by 70-and 40 folds,respectively,in the leaf tissues of grape vine.Maize seedlings inoculated withP.putidaunder drought conditions were shown to be more tolerant to stress than uninoculated seedlings.Bacteria were reported to regulate IAA and ABA production and enhance root and shoot lengths and relative water content of maize seedlings(Skzet al.,2018).It is suggested that further studies should be conducted in order to elucidate the mechanisms involved in inducing ABA secretion by rhizospheric microbes and approaches that could be used to harness their beneficial roles under drought.
Brassinosteroids are another important phytohormones whose main function is to promote seed germination,stem elongation,fruit ripening,and vascular differentiation.Furthermore,several studies have indicated the role of BRs in regulating specific responses in plants under drought stress(Nolanet al.,2020).Negative regulation of BRs was shown to promote stomatal closure inVicia faba,whereas its exogenous application toA.thalianaandBrassica napusseedlings was reported to induce drought resistance.Studies have shown that drought-responsive brassinosteroid-insensitive 2(BIN2)can phosphorylate and activate the ubiquitin receptor protein DSK2,thereby improving plant growth coordination and survival(Nolanet al.,2017).Brassinosteroids enhance ABA-induced stomatal closure and resistance to drought stress.Studies have shown that both BRs and ABA regulate stomatal opening and repress stomatal development in some plants,and BRs could further play a role in stomata formation.Moreover,BRs have been reported to induce stomatal closure in Arabidopsis through the signal transduction pathway of ethylene hormone(Shiet al.,2015).Several studies reported an interaction between BR and ABA whereby both induced stomatal closure during drought stress.In addition,BRs were reported to increase the synthesis of ethylene,resulting in the accumulation of phenolic and terpenoid compounds which scavenge on ROS.Further studies are required to elucidate the transcriptional regulation of BRs and their molecular interaction with ABA pathway.A clear understanding of the signal transduction pathway of BRs could be employed to enhance the degree of stress response in crops cultivated in drought-prone areas.
Other important phytohormones that induce plant responses against drought include auxins,cytokinins(CKs),jasmonic acid,and ethylene hormones.High concentrations of auxins normally induce unidirectional efflux of K+channels which result in stomatal closure(Shiet al.,2014).In addition,water deficiency significantly reduces the production of CK and induces its transport to roots,leading to the closure of stomata.Notably,research has indicated that plants which naturally produce low levels of CKs likely exhibit higher drought stress tolerance,with enhanced ABA sensitivity and high cell membrane integrity.Jasmonic acid,on the other hand,is induced by drought stress and also interacts with ABA,resulting in decreased stomatal aperture.Strigolactone and ethylene hormones also interact with the ABA signal transduction pathway to generate stress responsesviastomatal closure.Further studies need to focus on the interaction of the signal transduction pathways of respective hormones during drought in order to understand their individual and combined effects on alleviating drought.Studies may also focus on the mechanisms of individual plants in generating resistance against drought stress through the production of phytohormones.
RHIZOSPHERIC MICROBES ALLEVIATING THE EFFECT OF DROUGHT ON PLANTS
There is great demand for efficient and environmentfriendly agricultural approaches under biotic and abiotic stresses(Ullahet al.,2019).Although plants may adapt to drought stress through various mechanisms that involve phenotypic resilience,studies have reported their interactions with rhizosphere microbiomes to generate enhanced resistance against the adverse effects of drought(Farraret al.,2014).Naturally,there exists a tightly knit mutual relationship between plants and rhizospheric microorganisms.Rhizosphere contains billions of microbial cells per gram root and is strongly influenced by secretions from plant roots(Egamberdievaet al.,2008).Because of the close genetic interactions between plants and their microbiomes,rhizospheric microbial genome is sometimes considered as the plant second genome(Vandenkoornhuyseet al.,2015).Owing to global warming effects,including rising temperatures,disruptive dry seasons,and reduced yield,there has been a surge in studies seeking to understand the role of rhizospheric microbes in drought stress tolerance(Ahkamiet al.,2017).
Notably,many bacterial strains isolated from rhizospheres have been identified as plant growth-promoting(PGP)bacteria.They possess the ability to enhance plant growth through nitrogen fixation,nutrient solubilization,induction of systemic resistance,generation of plant growth regulators,organic acids,and volatile organic compounds,and secretion of protective enzymes,such as ACC-deaminase and glucanase(Ahkamiet al.,2017).In addition,PGP rhizobacteria(PGPR)facilitate plants to develop systemic resilience to drought stress(Reubenet al.,2013).Marascoet al.(2018)investigated the ability of various bacterial microbiomes normally associated with the rootstocks of grapevine to enhance plant growth during drought stress.Eight isolates containing over 510 strains were identified and assessed for their ability to support grapevine growth under water deficient conditions.The selected bacterial strains exhibited a wide variety of PGP traits,whereas the observation of GFP-labelledPseudomonasandAcinetobacteriaisolates revealed their ability to attach to and colonize roots of both Arabidopsis and grapevine.An experiment on pepper plants fertilized with the selected strains under both optimal irrigation and drought condition shows that PGP activity was a stress-dependent and not intrinsic feature of the strains.The bacterial isolates exhibited the ability to increase shoot length,leaf,shoot-root biomasses,and photosynthetic activity in drought-stressed grapevines.Essentially,the results showed that PGPR enhanced the adaptation of plants to drought through the stress-induced promotion of droughttolerance ability(Rolliet al.,2015).Furthermore,PGPR were reported to alter root elasticity of cell membranes and enhance tolerance against water deficiency(Dimkpaet al.,2009).Rhizospheric PGP bacteria also enhance drought stress tolerance in plant hosts by producing hormones that can induce the expression of diverse plant genes associated with drought tolerance(Lau and Lennon,2011).Inoculation of PGP isolates from generaBacillus,Paenibacillus,Brevibacillus,andMicrococcusexhibited important changes in Arabidopsis gene expression under drought stress.The PGPB isolates were indicated to induce the expression ofAtRAB18andAtLT178genes in Arabidopsis,which are involved in plant drought resistanceviaABA signalling pathway(Sukweenadhiet al.,2015).
Additionally,rhizospheric bacteria produce hormones or exhibit the ability to induce plant hormones which are important in promoting drought stress resistance.Plant hormones,such as IAA,produced by bacteria in the rhizosphere enhance water and nutrient uptake by stimulating root growth and lateral root formation.For instance,Azospirillumbacteria are known to produce IAA and were shown to enhance plant tolerance to drought stress(Dimkpaet al.,2009).The plant hormones produced by the bacteria in the rhizosphere confer beneficial effects on drought-stressed plants.A study by Dimkpaet al.(2009)further reportedA.brasilenceto promote the root growth ofP.vulgarisunder water-deficient conditions.Wheat and maize seedlings inoculated withA.brasilenseunder drought stress showed increased tolerance,with increased relative and absolute water contents and grain yield(Arzaneshet al.,2011).In the future,drought protective agents such as commercial phytohormones and microbial products will be of great importance for plants due to the global climate change scenario.Therefore,beneficial microbes could be a stress-protecting agent for plants and a promising solution for sustainable and environment-friendly agriculture(Ullahet al.,2019).Further studies may be required to provide a detailed molecular understanding of the complex plant-microbe interaction within the rhizosphere.This could be achieved by elaborating on the comprehensive regulatory mechanisms that are involved in the selection and activity of rhizospheric microbes by plant roots,which may provide new opportunities to promote crop production.
To mitigate the effects of drought,however,plants have been reported to solely rely on root-associated microflora(De Vrieset al.,2017,2020;Cavicchioliet al.,2019;Nayloret al.,2020).Microorganisms existing within the plant interact with them by various means,depending on a large number of signal perception mechanisms adapted by both partners.For instance,the capabilities of generating and sensing signals make the entire population spread out as a biofilm on the root surface to begin an intensive action.As such,microbes reach a certain density,a process of communication known as quorum sensing.By means of root exudation,the rhizospheric bacteria then chemotactically stick to colonizing the root surface,further becoming endophytes(Naseemet al.,2018).Various species of rhizospheric microbes,includingRhizobiumsp.,Bacillussp.,andPseudomonassp.,produce exopolysaccharides(EPS),an important class of polymeric materials that remarkably enhance the ability of bacteria to colonize special ecological niches under drought stress(Putrieet al.,2013).Exopolysaccharides found in the rhizosphere are generally appreciated due to their role in improving the moisture-holding capacity of drought-stricken soil(Kaushal and Wani,2016).Moreover,the presence of EPS-producingRhizobiumsp.has been reported to significantly promote soil aggregation and lead to water retention in the rhizosphere(Fig.2).In a study aimed at evaluating EPS production by drought-tolerantBacillusspp.,Vardharajula(2014)noted that EPS-producingB.amyloliquefaciens,B.subtilis,andB.licheniformissignificantly improved soybean seed germination,and growth of seedling under drought stress,and also induced soil aggregation(Table I).Furthermore,EPS-producingP.putidastrains form biofilm on the root surface of maize and sunflower seedlings have indicated to improve soil structure and the ability of soil to hold roots during drought.When maize was inoculated withP.putidaunder drought stress,the seedlings showed increased root and shoot growth,higher dry biomass,andimproved cellular metabolites and stomatal conductance(Vurukondaet al.,2016).In Arabidopsis,P.chlororaphisO6 was found to colonize the root,thereby inducing drought tolerance.It has been shown that genes upregulated under water deficiency may be highly influenced by microbial colonization,leading to plant protection and the activation of the plant defense signal pathway.It is,however,hypothesized that microbial colonization is a key element for induced systematic tolerance through soil biota.Exopolysaccharides mediate bacterial survival against water stress by facilitating water retention and regulating diffusion of organic C sources.They also aid the irreversible attachment and colonization of microbes to the rootsviaa network of fibrillar material that permanently links the bacteria to the root(Aliet al.,2014).Future studies are needed to provide more detailed insights into the mechanisms of microbial EPS.Genetic approaches can be used for the large-scale production of microbial EPS,which could be used to treat germinating seedlings that are to be grown in water-deficient areas.
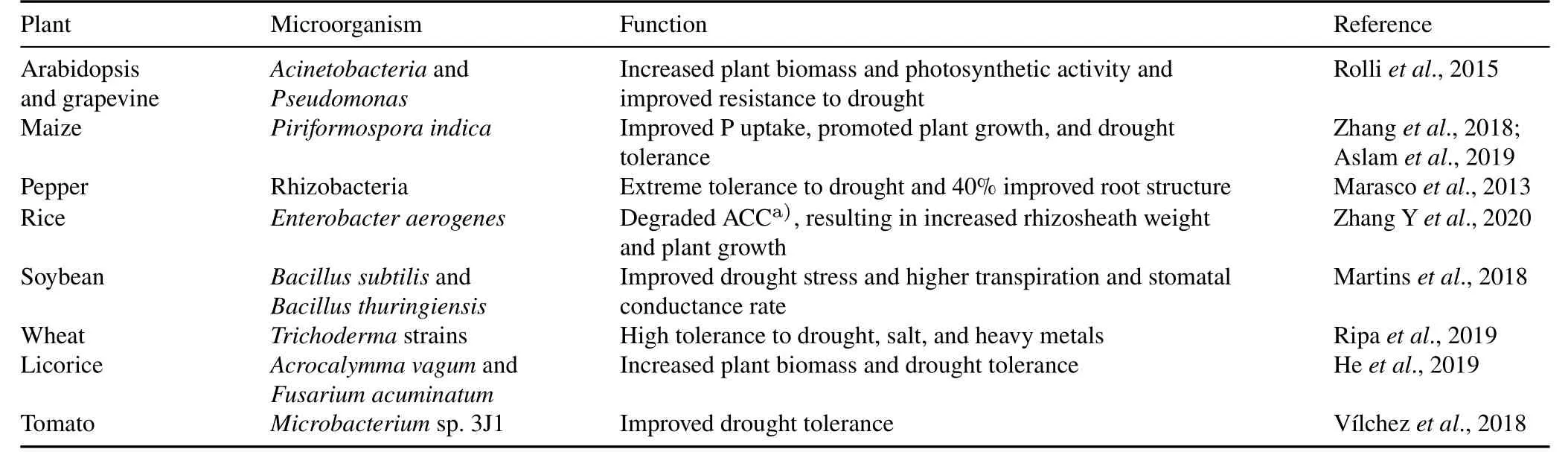
TABLE I Role of soil microbial community in crop tolerance to drought stress
Plants that adopt various morpho-anatomical,physiological,biochemical,and molecular responses to survive under drought are,in part,mediated by several endophytic bacteria.The interactions between plant and endophytic bacteria are currently gaining attention.They play effective roles in increasing crop production and providing drought tolerance(Ullahet al.,2019).Both root exudates and microbial secretions in the rhizosphere under both WW and moderate drought conditions are speculated to exert effects on plant physiological responses to subsequent drought conditions(Fig.1)(Ullrichet al.,2018).Recent studies have outlined the ability of endophytic microorganisms in enhancing drought and heat tolerance in crops such as wheat and sugarcane.Endophytes often originate from rhizosphere and have been shown to synthesize anti-stress biochemicals or activate host stress response systems when exposed to drought(Lataet al.,2018).
Moreover,symbiotic AMF were reported to exponentially replicate under drought conditions and convey drought tolerance to the host plants by enhancing antioxidant production.An increased antioxidant enzyme activity is known to reduce oxidative stress and promote efficient water usage in plants(Thirkellet al.,2017;Ahammedet al.,2020).Furthermore,Kouret al.(2020)identifiedStreptomyces laurentiiandPenicillium sp.as drought-adaptive and P-solubilizing microbes that can efficiently enhance the accumulation of diverse osmolytes and increase the chlorophyll content in millet under drought.Researches have shown that the majority of stress-tolerant rhizospheric microorganisms in droughtprone areas exhibit thick peptidoglycan cell walls,osmolyte production,dormancy,and sporulation(Schimelet al.,2007;Xu and Coleman-Derr,2019).Such microbial trait responses protect the microbes against desiccation and further enable them to thrive in harsh conditions as they continue to confer their benefits to plant hosts.Many of these beneficial traits conferred by microbes during drought are common among several rhizospheric microorganisms,such as grampositiveActinomycetes,Streptomyces,andGlomeromycetes(Finkelet al.,2017).However,several factors,including stress duration and intensity and bacterial abundance,determine bacterial response to drought at various organizational levels(Naseemet al.,2018).Although several studies have highlighted these beneficial traits of rhizospheric microbes to plants under drought,future studies may be needed to provide the explicit mechanisms of individual microorganisms in helping their plant hosts to alleviate the negative effects of drought stress.Further studies may also be required to investigate whether these microbes can be applied to drought-stressed plants under realistic field conditions.
FUTURE PERSPECTIVES
There is growing interest among agricultural scientists to understand the realistic mechanisms of plant-microbe interactions for drought-resilient crop production.Selection of potential genetic traits that are directly involved in improving plant growth under stressed conditions may come from selective breeding or bioengineering of crops.Exogenous application of peptides,phytohormones,or bacterial inoculation can be used for fine tuning of drought responses to sustain future agricultural yields.However,understanding of complex feedback mechanisms of plant-microbe responses,particularly under field conditions,will provide new insights into enhancing crop yield and productivity to meet global food demand and improve agricultural sustainability.
CONTRIBUTIONS OF AUTHORS
Mehtab Muhammad Aslam conceived the study and designed the figures.Mehtab Muhammad Aslam,Eyalira J.Okal,and Aisha Lawan Idris wrote the first draft.Mehtab Muhammad Aslam,Zhang Qian,Joseph K.Karanja,Shabir H.Wani,Weifeng Xu,and Wei Yuan critically improved and reviewed the manuscript.All other authors read and approved the final version.
ACKNOWLEDGEMENTS
We are grateful for the grant support from the National Key R&D Program of China(Nos.2017YFE0118100 and 2018YFD02003025),National Natural Science Foundation of China(Nos.31761130073,31872169,and 31600209),a Newton Advanced Fellowship,UK(NO.NA160430),Fujian Province Education Department Funding,China(No.JK2017015),Research Grant of Fujian Agriculture and Forestry University,China(No.KXGH17005),and European Union’s Horizon 2020 Research and Innovation Programme under Project SHui(No.773903).Special thanks are given to Prof.Ian Dodd from Lancaster University,UK,for his insightful comments and contribution to this manuscript.
杂志排行

Pedosphere的其它文章
- Elevated carbon dioxide stimulates nitrous oxide emission in agricultural soils:A global meta-analysis
- Hydrogen cyanide production by soil bacteria:Biological control of pests and promotion of plant growth in sustainable agriculture
- Effects of different continuous fertilizer managements on soil total nitrogen stocks in China:A meta-analysis
- Microplastics in soil:Impacts and microbial diversity and degradation
- Difficult-to-culture bacteria in the rhizosphere:The underexplored signature microbial groups
- Effects of plant growth-promoting rhizobacteria on the molecular responses of maize under drought and heat stresses:A review