白蜡属种间杂交子代木质素含量变异及FmPAL核苷酸多态性关联分析
2022-03-02尹一卜李吉祥郭樱杰芦子廷刘华领詹亚光曾凡锁
尹一卜 李吉祥 郭樱杰 芦子廷 肖 英 刘华领 詹亚光 曾凡锁*
(1. 东北林业大学生命科学学院,哈尔滨 150040;2. 黑龙江省山河屯林业局,哈尔滨 150232)
木质素是高等陆地植物细胞壁的主要结构成分,它增强细胞的机械支持力和抗压强度,促进了水的运输,并成为抵抗病原体的物理屏障。木质素的含量和组成在植物物种间以及植物体内部有很大差异,并受发育和环境因素的影响。木质素单体(S-,G-和H-木质素的单体)生物合成途径中大多数酶及其功能已被确定。目前,可以利用木质素生物合成途径中关键基因(等)的特定突变、遗传修饰及差异表达来改变木材品质性状。苯丙氨酸途径受到干扰后,途径中间体可能会成功整合到木质素聚合物中,从而影响其理化性质。苯丙氨酸氨解酶(PAL)和酪氨酸氨解酶(TAL)催化苯丙氨酸向反式肉桂酸的转化,并将碳流从莽草酸途径引入苯丙烷代谢的各个分支,这是苯丙烷代谢途径的第一步,也是初级和次级代谢交叉的重要调节点。研究表明,同一植物中不同的基因表达模式及调控机制具有显著的差异性。在拟南芥()中具有4个不同的基因,其表达具有组织特异性,其中和表达模式较为相似,均在拟南芥的根、花、茎中表达水平较高,而在上述部位中几乎没有表达,获得的、、、四突变体中木质素的含量降低,并且大大减少茎中木质素和其他酚类化合物的含量,这些突变体在生长、繁殖力和对环境压力的耐受性方面都受到损害。白梨()基因在木质化组织(根和茎)中的的转录水平高于木质度较低的组织(叶、芽和花),在过表达拟南芥中增加了木质素含量和细胞壁厚度,参与木质素生物合成。本文的前期研究克隆了水曲柳()基因,其与梨树()中参与木质素合成的PAL基因具有极高同源性。基因在木质化程度较高的水曲柳枝中表达量最高,具有组织特异性。表达模式在水曲柳的不同部位显示出与木质素含量相似的变化趋势,初步说明可能参与水曲柳中木质素的生物合成。在非生物胁迫下,的表达量显著上升。植物的苯丙烷类代谢途径能够被非生物胁迫激活,从而增加基因的活性,提高植物抗逆境能力,起到保护和调节的作用。转录水平随环境因素变化,例如高光/紫外线、病原体攻击、低氮、低磷酸盐、低温和伤害都可以改变基因的转录水平。
水曲柳为木犀科(Oleaceae)白蜡属(又称为梣属)落叶乔木,是东北珍贵的阔叶用材树种之一。该树种是中国木材生产的主要商业树木之一,在东北的生态和环境保护中发挥着重要作用。前期研究中,本团队已经获得水曲柳与大叶白蜡()、小叶白蜡()和绒毛白蜡()的杂种F1,并营建了对比试验林。苯丙氨酸解氨酶(PAL)是苯丙烷类代谢中的关键酶和限速酶,解析的等位基因变异对水曲柳生长和木材品质性状的影响机制具有重要意义。分析各杂交组合F1 木质素含量,并与其母本自由授粉子代进行比较,可揭示水曲柳各杂交组合木质素含量的变异规律。通过序列测定,筛选F1 代各无性系中的SNP 位点,与木质素含量进行关联性分析,探究基因内部碱基变化对水曲柳木质素含量的影响,为从分子水平对水曲柳的杂交子代的选育和木材改良研究奠定坚实的基础。
1 材料与方法
1.1 试验材料与试验方法
1.1.1 试验材料
本研究材料取自东北林业大学帽儿山林场,71 个白蜡属种间杂交组合子代及16 个母本自由授粉子代。其中,亲本为水曲柳×大叶白蜡的杂交组合共有25 个、亲本为水曲柳×小叶白蜡的杂交组合共有18 个、亲本为水曲柳×绒毛白蜡的杂交组合共有28 个、母本水曲柳的对照组合共有16 个。每个杂交组合取2~3 株,共176 个无性系,平均每棵树上取3~4 个当年新生枝条。将水曲柳枝用固体粉碎机搅碎呈粉末状,装入信封,标明其对应的水曲柳无性系,放于干燥通风处以备后续试验使用。另一部分取每株新生枝剥取树皮,放入-80 ℃冰箱保存,用于总DNA提取。
1.1.2 水曲柳新生枝木质素含量测定
取一定量烘干的水曲柳粉末(80 目),将样品放在65 ℃下烘干至恒质量。取0.05 g 样品溶于1 mL溴乙酰冰醋酸溶液(25%)中,70 ℃水浴30 min,冷却后加入0.4 mL(2 mol·L)NaOH 终止反应,再加入1 mL 冰醋酸和0.04 mL(0.5 mol·L)羟氨盐酸,离心收集上清液,加入蒸馏水稀释50 倍,测量280 nm处OD值A,以计算木质素的含量。
1.1.3 水曲柳总DNA的提取及克隆
将水曲柳新生枝树皮材料加入液氮研磨成细粉末状后加入到已预热(65 ℃)的CTAB 缓冲液中,放入金属浴仪器(65 ℃)中保温30 min,每5 min 摇动离心管,后把样品管插到冰上冷却2 min,加入700µL 氯仿,离心取上清液再次加入氯仿离心,此过程重复2次。取上清液加入等体积的异丙醇,轻轻摇匀后插到冰上,沉淀至少30 min。离心取沉淀用70%的乙醇洗涤2次,最后加入50µL的ddHO进行溶解,溶解后离心1 min,进行电泳。设计基 因 全 长 引 物 序 列(F:5′CTGCCA‐CATTCAACAACAAGAG3′,R:5′GTTTTAGTCCCAAAGTCTTGTTACA3′)并以提取水曲柳DNA 为模板进行扩增。电泳检测利用1%的琼脂糖凝胶检测扩增产物,选择特异性条带且大小在1 800 左右的扩增产物进行测序。
1.2 数据统计分析
采用SPASS19.0 软件对所测得的木质素含量进行统计分析。根据木质素含量进行差异显著性分析。
所需应用的变异系数的计算公式:

式中:代表所测含量或性状的平均值,代表代表所测含量或性状的对应于平均值的标准差。
对遗传力进行计算研究所需公式:

式中:值代表遗传力,代表样本进行放长分析时的输出项。
1.3 核苷酸多态性及SNP关联性分析
将序列比对结果进行统计,筛选出SNP 位点,对突变频率极低(<3%)的位点进行排除,计算SNP 频率及发生突变过程中转换和颠换的数量。利用DnaSP5.0 软件分析基因核苷酸多样性并进行中性检测;利用MEGA 软件对不同无性系间基因的遗传距离进行分析;再利用DnaSP5.0软件对基因的SNP位点进行连锁不平衡分析;最后结合其对应无性系的木质素含量,利用SPSS 软件进行差异检测及关联性分析。
2 结果与分析
2.1 白蜡属种间杂交组合木质素含量分析
测量分析不同杂交组合子代木质素的含量(见表1),在水曲柳×大叶白蜡组中70 号杂交组合木质素含量最高为(32.96±5.25)%。水曲柳×小叶白蜡组和水曲柳×绒毛白蜡组中129号和7号杂交组合的木质素含量为(35.29±1.71)%,(39.79±7.15)%达到组合内最高水平。而母本自由授粉子代中135 号木质素含量达到了最高值为(37.20±3.73)%,但是其中大多数杂交组合水曲柳的木质素含量是高于母本对照水曲柳组的平均含量。70号、129 号,7 号杂交组合木质素遗传增益分别13.59%,25.57%,36.33%。
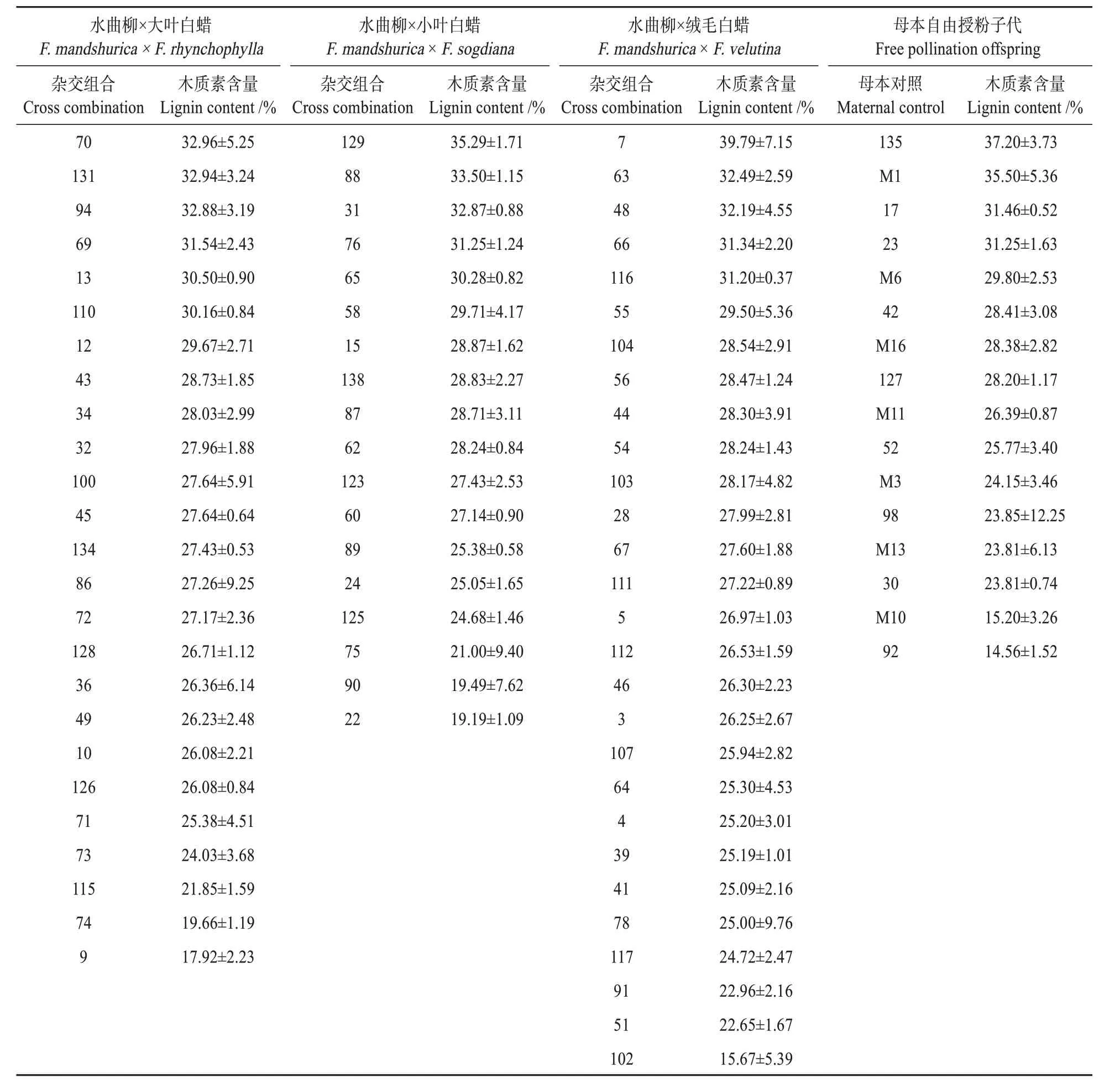
表1 白蜡属种间杂交组合木质素含量均值统计Table 1 Mean statistics of lignin content in Fraxinus spp.hybrid combinations
分析71 个杂交组合和16 个母本自由授粉子代间木质素的含量差异(见表2),根据SPSS 统计学分析表明,各杂交组合间木质素含量差异极显著(<0.01)。水曲柳×大叶白蜡组、水曲柳×小叶白蜡组、水曲柳×小叶白蜡组及母本自由授粉子代的木质素含量的变化范围分别为17.28%~36.07%、11.98%~35.29%、11.80%~39.79%和11.59%~37.20%(见表2)。各组合内木质素含量差异显著,其各组间的均值范围为25.53%~27.38%,各杂交组合内木质素含量的均值分别为:水曲柳×大叶白蜡(27.31±4.72)%;水曲柳×小叶白蜡(26.50±6.47)%;水曲柳×绒毛白蜡(27.38±5.20)%;母本自由授粉子代水曲柳(25.53±7.16)%。各杂交组的木质素含量均与母本自由授粉子代水曲柳差异显著,遗传力范围为0.657~0.895。
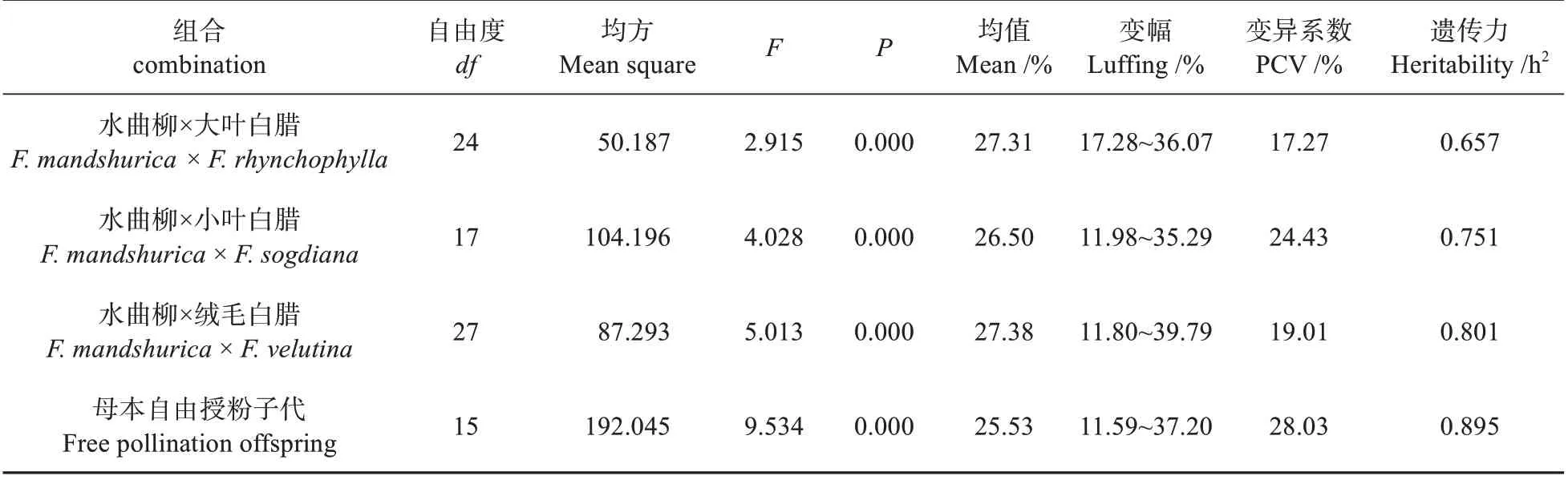
表2 白蜡属各组合木质素的方差分析及含量变异参数分析Table 2 Variance analysis and content variation parameter analysis of lignin in various combinations of Fraxinus spp.
2.2 各杂交组合内无性系间木质素的含量及变异参数均值分析
对176 个无性系木质素含量进行分析(见表3),其中亲本为水曲柳×大叶白蜡母本水曲柳杂交子代无性系53 个;水曲柳×小叶白蜡杂交子代无性系28 个;水曲柳×绒毛白蜡杂交子代无性系66个;母本自由授粉子代水曲柳无性系28 个。根据统计学分析,水曲柳杂交组合内各无性系木质素含量差异显著,具有进一步研究价值。
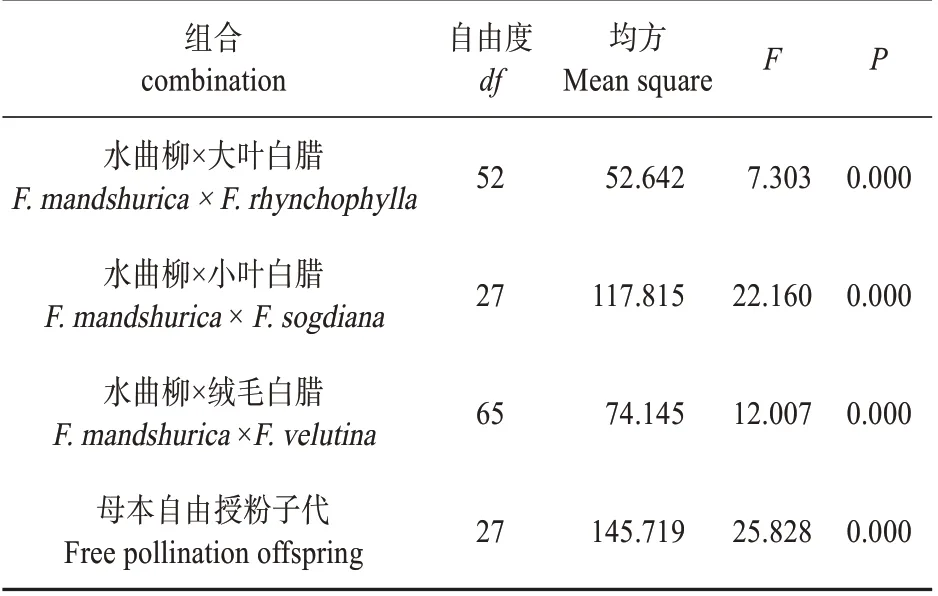
表3 各组合内无性系木质素含量方差显著性分析Table 3 Significance analysis of variance of lignin con‐tent in clones of different combinations
对176 个无性系木质素含量变异情况进行分析,发现水曲柳×大叶白蜡组、水曲柳×小叶白蜡组、水曲柳×小叶白蜡组,母本自由授粉子代组内各无性系间遗传力(h)分别为0.863、0.955、0.917、0.961。
对各杂交组合无性系进行统计分析,其中三个不同杂交组合中水曲柳木质素含量最高的无性系分别为:水曲柳×大叶白蜡组Ⅱ-100-3(36.07±0.86)%,水曲柳×小叶白蜡Ⅱ-129-6(35.29±1.71)%,水曲柳×绒毛白蜡Ⅱ-7-10(39.79±7.15)%。母本自由授粉子代水曲柳中木质素含量最高的为I-135-8(37.20±3.73)%。根据统计结果对其中前5%优良生长性状无性系进行筛选,结果为水曲柳×大叶白蜡组I-86-5 和Ⅱ-100-3 的木质素遗传增益分别为24.38%和27.69%;水曲柳×小叶白蜡Ⅱ-129-6的木质素遗传增益为31.67%;水曲柳×绒毛白蜡Ⅱ-7-10、I-48-4 和I-55-1 的木质素遗传增益分别为41.59%、34.64%和29.17%。
2.3 水曲柳基因组总DNA 的提取及FmPAL 基因的扩增
分别对176 株不同无性系水曲柳总DNA 进行提取,电泳有清晰DNA 条带,且OD值约为1.8,样品质量较高,能够满足后续对基因的扩增。基因的全序列共有1 893个碱基,其中编码区长度为1 674 bp,设定扩增片段长度为1 810 个碱基,其设计的引物序列范围为13~34,1 799~1 823。对基因进行扩增对其扩增产物进行检验,其产物长度约为1 810 bp(见图1)。
2.4 FmPAL基因中SNPs位点的筛选
利用BioEdit 软件将基因测序结果进行拼接,发生突变位点中,有一些位点的突变频率极低(<3%),为防止测量误差造成的影响,这些位点不算作SNP 位点。筛选出的SNP 位点共有33个,根据比对结果部分序列筛选SNP位点如图2所示,其中C-T突变的位点有12个(90位,174位,306位,334 位,432 位,654 位,718 位,744 位,783 位,837 位,1 167 位,1 182 位);A-C 突变位点1 个(604位);G-A 突变位点12 个(204 位,348 位,435 位,603 位,623 位,657 位,796 位,829 位,1206 位,1 272 位,1 412 位,1 638 位);T-G 突变位点2 个(753 位,1 165 位);C-G 突变位点5 个(91 位,140位,387 位,828 位,1 421 位);A-T 突变位点1 个(589 位)均出现在编码区。基因中属同义突变16 个,属非同义突变17 个。进行SNPs 位点筛选的基因总长度为1 674 bp,故基因内部SNP 位点出现的概率为1/51。
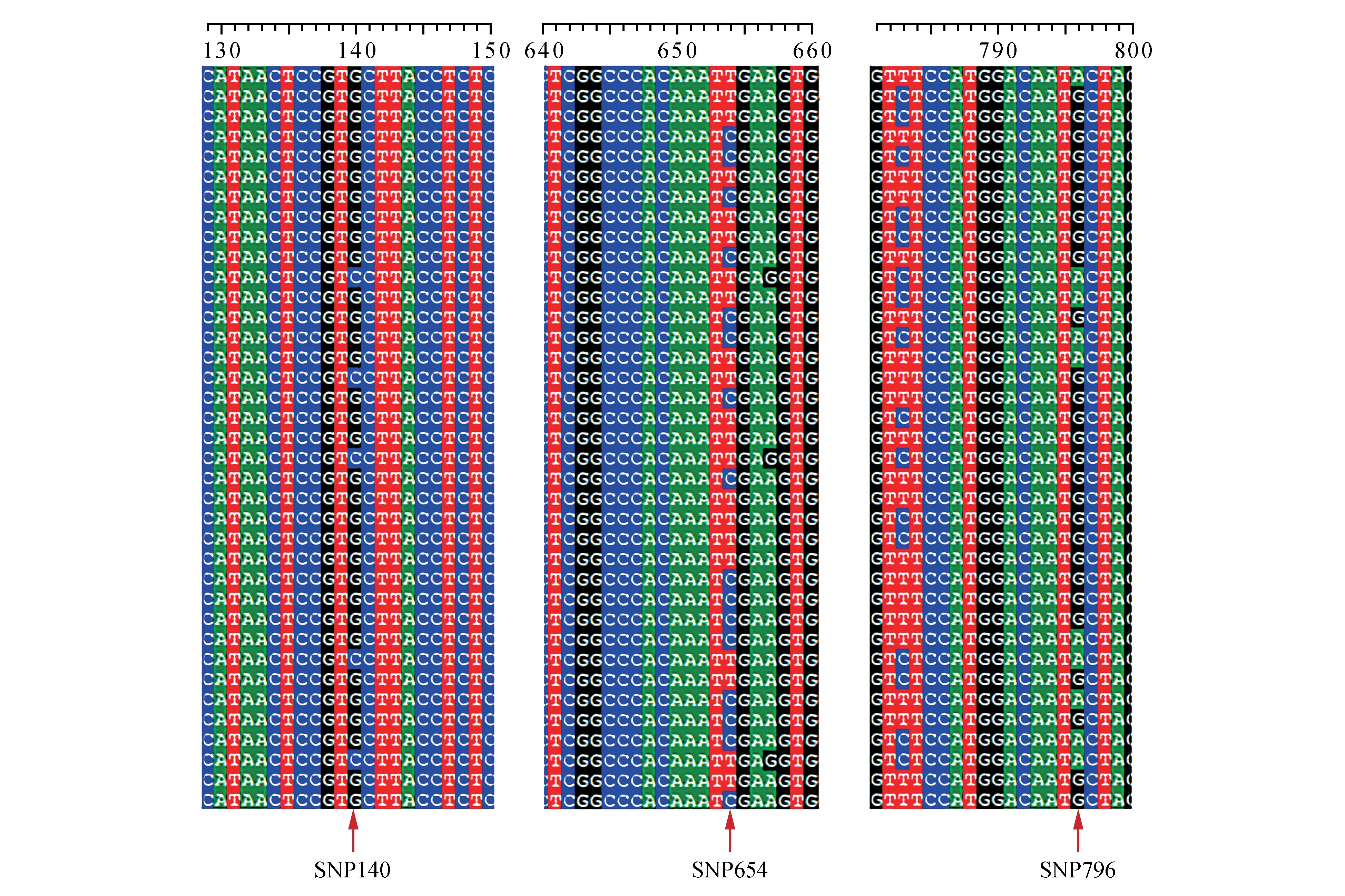
图2 FmPAL基因序列比对结果Fig.2 Comparison of FmPAL gene sequences
2.5 核苷酸多态性分析
SNP 多态性指的是DNA 序列中单个碱基突变而引起的多种排列方式。这种突变的产生,主要由转换(C-T、G-A)和颠换(C-A、C-G、G-T、A-T)引起,本研究筛选出的33个SNP位点中,转换突变位点24 个,颠换突变位点9 个。为了更深刻理解水曲柳群体的遗传多样性,利用DnaSP5.0 软件对于比对序列的核苷酸多样性()进行分析,值反应了基因的相似性及多样性,结果显示所研究的基因中,多态位点数为93,经筛选去除突变频率极低(<3%)位点后的有意义SNP 位点数为33 个,核苷酸多样性值为0.007 45,单倍型数是131,单倍型多样性值为0.999 7,核苷酸平均差异数:12.423。其中,单倍体多样性反应了群体总体的遗传多样性,越高则遗传资源水平越丰富,抽取样本时不同的概率越大。本研究中的水曲柳群体的基因单倍体多样性值达到了0.999 7,属高水平,证实了基因具有较高的变异,遗传资源多样。
2.6 不同无性系中FmPAL基因的遗传距离
遗传距离是用于衡量不同基因序列的综合遗传差异大小的指标。分析基因的遗传多样性,对水曲柳不同无性系的基因测序结果进行比对后,利用MEGA 生物学分析软件,计算不同无性系之间的遗传距离。结果表明基因总体平均遗传距离为0.010,各无性系间遗传距离的分布范围是0.000~0.030。对比发现,无性系Ⅰ-33-3、Ⅱ-3-5-2 和Ⅰ-3-3-9 与其它大部分无性系的基因之间遗传距离都大于0.020 cM,其中最大的遗传距离达到0.030 cM(Ⅱ-3-5-2 与Ⅱ-91-3、Ⅰ-33-3 与Ⅱ-91-3、Ⅱ-3-5-2 与Ⅱ-66-8),其次的遗传距离达到0.029 cM(Ⅱ-3-5-2与Ⅰ-28-1、Ⅱ-3-5-2与Ⅰ-105-2、Ⅰ-73-6 与Ⅰ-33-3、Ⅰ-73-6 与Ⅱ-3-5-2、Ⅰ-73-6 与Ⅰ-3-3-9、Ⅱ-66-8 与Ⅰ-33-3、Ⅱ-66-8 与Ⅱ-3-5-2、Ⅱ-66-8 与Ⅰ-3-3-9)。这说明Ⅰ-33-3、Ⅱ-3-5-2 和Ⅰ-3-3-9 号无性系的变异程度高,遗传差异最明显。而遗传距离最小的为0.000 cM,多对序列间的距离呈现最小值,包括Ⅱ-72-9 与Ⅱ-38-2、Ⅰ-44-8 与Ⅱ-72-2、Ⅱ-38-2 与Ⅱ-70-1。参考BioEdit 软件的比对结果,这些遗传距离小的无性系之间基因序列基本是一致的,而遗传距离较大的两个无性系之间,基因序列上的碱基差异较大,故遗传距离的计算对于不同无性系基因变异及差异性的描述具有显著意义,为表型数据的分析提供重要参考。
2.7 中性检验及连锁不平衡分析
通过对所研究的DNA 序列进行基础检测分析,判定此物种群体的DNA 是否可以遵循中性进化模式。我们利用DnaSP5.0 软件对基因在进化过程中的自然选择的作用情况进行中性检验,结果显示,基因的Tajimas D 值为-0.957 39,Fu 和Li值为-1.563 74,均为负值,推测群体序列的进化方式为负向选择。关联作图,也称为LD 作图或全基因组关联研究(GWAS),是连锁分析的一种映射选择,应用DnaSP 5.0 软件中的LD 程序进行对176 个不同水曲柳无性系基因连锁不平衡进行估算。由图3 可见随着基因中核苷酸序列长度增加,基因无明显衰退趋势,其不平衡水平缓慢上升。值由0.1 缓慢上升至0.2。说明基因中SNP 位点的连锁不平衡水平无明显衰退。

图3 FmPAL基因的LD连锁不平衡图谱R2值是衡量两个位点连锁不平衡程度的重要指标;R2表示两个位点在等位基因中出现的频率;当R2=1,表示连锁完全不平衡,没有重组;当R2=0,表示连锁完全平衡,随机组合Fig.3 LD linkage disequilibrium map of FmPAL geneR2 value is an important index to measure the degree of linkage dis‐equilibrium between two loci;R2 represents the frequency of the two loci appearing in the alleles;R2=1,indicates complete linkage disequi‐librium,no recombination;R2=0,indicates complete linkage balance,random combination
2.8 单个SNP关联分析
结合水曲柳无性系的木质素含量,对所有33个位于基因编码区的单个SNP位点与木质素含量进行关联分析。关联分析中具有显著差异的 位 点 分 别 是SNP140(G-C)、SNP796(G-A)、SNP654(T-C)。在SNP140 和SNP796 中突变型纯合体木质素含量显著高于野生型纯合体木质素含量,但在SNP654 中突变型纯合体木质素含量显著低于野生型纯合体木质素含量。SNP140的CC突变纯合基因型水曲柳木质素含量为(29.03±3.53)%,GG 野生纯合型木质素含量为(26.86±5.39)%,两种基因型之间具有显著差异(=0.044);SNP654的野生纯合TT 基因型水曲柳木质素含量为(28.49±4.72)%,CC 突变纯合型木质素含量为(25.67±5.95)%,两种基因型之间具有显著差异(=0.003);SNP796的GG野生纯合基因型水曲柳木质素含量为(26.91±5.47)%,AA 突变纯合基因型木质素含量为(29.03±4.74)%,两种基因型之间差异显著(=0.039)。其余30 个SNP 位点其突变纯合基因型水曲柳木质素含量与野生纯合型基因木质素含量两种基因型之间差异不显著,说明这30 个SNP位点单一突变对木质素含量影响不明显。
3 讨论
目前,水曲柳作为珍贵阔叶用材树种,种质资源选择和杂交育种是其遗传改良的重要手段。林木遗传变异是选择的前提,前期研究发现白蜡属的各杂交组合水曲柳×大叶白蜡组、水曲柳×小叶白蜡和水曲柳×绒毛白蜡组与母本自由授粉子代对照组相比其树高、胸径均存在显著差异。本研究也发现各杂交组合的木质素含量与母本也存在显著差异,表明生长性状和木材性状存在丰富的变异基础,这为优良杂交种的选择奠定了基础。表型是在遗传和环境的共同作用下形成的。李昌荣等研究9.5年生的大花序桉()胸径、树高、单株材积、生材密度等,明确了大花序桉生长性状对材性性状间接选择效果不好,基本密度对其他材性性状的间接选择效果较好。多性状综合指数法可以选择出部分生长和材质兼优的家系。本研究通过对白蜡属种间杂交组合无性系木质素含量进行研究,有利于筛选各杂交组合中优于母本自由授粉子代的组合及无性系。通过分析木质素含量水平的高低,有利于种间杂交子代的木材性状评价及实际生产应用的深入研究。
1989 年日本学者Fumio Tajima 首次建立了中性检测(Tajima Test),现在广泛的应用于关联性检测前的基础检验。通过对所研究的DNA序列进行基础检测分析,判定此物种群体的DNA 是否可以遵循中性进化模式。通过中性检测推测群体序列的进化方式为负向选择。连锁不平衡(LD)为基础的关联映射已替代传统的QTL 作图,用于解析性状的复杂性。这种新颖的方法易应用于自然或育种种群,以检查自然等位基因变异与感兴趣的性状之间的关联,并提高标记/性状关联的分辨率。LD 的估计值从一个染色体区域到另一个染色体区域变化,并且在周围区域中更高。通过分析发现基因中SNP位点的连锁不平衡水平无明显衰退。
SNP 由于其分子标记在基因组内数量丰富,在编码区和非编码区皆广泛分布,已成为鉴定树木品种身份的辅助方法之一。在油橄榄()研究中,通过11 个核心SNP 位点对57 种油橄榄品种进行区分。SNP 位点的筛选分析还有助于对植物表型相关基因的研究。例如在水稻()气孔性状相关基因的研究中,分析EPF-TMM-SDD 途径相关基因在粳稻(var.)和籼稻(subsp.)的SNP 位点的不同,为其性状差异的机制解析奠定基础。本研究通过对176 个水曲柳无性系基因进行测序,从总长度为1 674 bp 的序列中筛选出共33个SNP位点,其基因内部SNP位点出现的概率为1/51,包含同义突变18 个,非同义突变15个。本文明确了等位基因变异与水曲柳的木质素含量密切相关。单倍型多样性值为0.999 7,核苷酸平均差异数:12.423,说明基因具有较高的变异,遗传资源丰富多样。关联分析确定了与木质素含量显着相关的3个SNP,分别为SNP140、SNP654、SNP796,而其余SNP位点并没有对木质素含量产生明显影响,其原因可能为这些单一的非同义突变位点可能与其它SNP 位点连锁共同发挥作用,而单一的SNP位点分析不能揭示其与木质素合成的关系,所以具体作用机制有待于进一步的分析。这些发现表明了参与木质素合成,并提供了与木质素性状相关的分子标记,在标记辅助育种中的有较好的应用潜力。由于大多数SNP 所解释的性状变异百分比很小,因此可能需要将这些重要标记组合在一起。通过结合水曲柳中高度保守的木质部转录组来解析这种复杂的关系,这将有助于构建次生生长和木材形成的调控模型,并设计出改善木材产量和品质性状的策略,包括缩短育种周期和增加早期选择准确性。
