五种双壳贝类贝壳微观结构观察与成分分析
2022-03-01莫天宝徐洪强董迎辉林志华
莫天宝, 徐洪强, 何 京, 董迎辉, 林志华,
五种双壳贝类贝壳微观结构观察与成分分析
莫天宝1, 2, 徐洪强3, 何 京3, 董迎辉2, 林志华2, 3
(1. 上海海洋大学 水产科学国家级实验教学示范中心, 上海 201306; 2. 浙江万里学院 生物与环境学院 浙江省水产种质资源高效利用技术研究重点实验室, 浙江 宁波 315100; 3. 浙江万里学院 宁海海洋生物种业研究院, 浙江 宁海 315604)
利用扫描电镜技术、X-射线粉末衍射技术和弱酸去钙法, 对菲律宾蛤仔、文蛤、厚壳贻贝、泥蚶、缢蛏5种习见海洋经济贝类贝壳的断面微观结构、物相组成、有机基质和蛋白质含量等进行了观察和分析。电镜观察结果显示, 5种贝壳的微观结构主要包含棱柱层和珍珠层; 棱柱层晶体结构有斜棱柱层、球棱柱层、棱柱层、均质层和交错板状层5种类型, 其中缢蛏只有斜棱柱层, 而泥蚶除球棱柱层外, 其他4种晶体类型均存在, 此复杂结构可能与其贝壳强度大有关; 珍珠层晶体结构有珍珠层和肌棱柱层2种类型, 其中厚壳贻贝的珍珠层呈典型的“砖-泥”结构, 具有明显的层状结构, 其余4种贝壳珍珠层均由不规则块状结构组成。X-射线衍射结果显示, 菲律宾蛤仔、文蛤、缢蛏和泥蚶4种贝壳都属于文石质壳体, 无机相几乎由文石组成, 而厚壳贻贝属于混合质壳体, 无机相由文石和方解石组成; 贝壳化学成分分析显示, 5种贝壳有机质含量均为3%左右, 而总蛋白含量占有机质的2.98%~7.21%, 其中可溶性蛋白是不可溶蛋白含量的5.55~20.31倍。上述结果为贝壳形成机理的研究积累了基础资料。
贝壳; 微观结构; 有机质; 扫描电镜; X-射线衍射
贝壳是一种天然的复合材料, 其优异的力学性能和良好的生物相容性历来是生物材料和仿生学等领域的研究热点之一[1-2], 而在水产领域中贝类壳色、壳型、壳厚度等性状近年来也引起诸多研究者和育种学家的广泛关注。在壳色方面, 国内学者相继培育出栉孔扇贝()“蓬莱红”[3]、海湾扇贝()“中科红”[4]、长牡蛎()“海大2号”和“海大3号”[5-6]、文蛤()“科浙1号”[7]和“万里红”[8]等优良新品种, 而这些壳色靓丽的新品种均能提高其经济价值; 壳型方面, 培育有“壳宽型”菲律宾蛤仔()[9]、“深凹壳型”香港牡蛎()[10]、“壳宽型”美洲牡蛎()[11]等新品系, 更受产业和市场的青睐; 在壳厚度方面, 开展了三角帆蚌()[12]、马氏珠母贝()[13-14]等贝壳珍珠层厚度和矿化机理相关研究, 而厚重坚硬的贝壳能更好地抵御敌害生物捕食、降低生产管理带来的机械损伤[15-17]。上述性状与贝壳的结构和成分有着密切关系, 但目前研究工作主要集中在腹足纲贝类[18-19]和双壳纲褶纹冠蚌()[20]、三角帆蚌[21]、马氏珠母贝[13]等育珠贝的贝壳微观结构类型、有机质和无机质特征及贝壳蛋白质的分子组成等[1], 其中以珍珠层的相关研究最为经典[22], 而对双壳纲其他贝类的贝壳结构和成分等的系统研究鲜有报道。
典型的双壳贝类贝壳分为3层, 角质层、棱柱层和珍珠层[23], 不同贝类其贝壳微观结构及层次排列特征不同, Carter等[24]对双壳贝类的贝壳形态、微观结构特征进行观察和分类, 根据其表型形态和晶体结构差异, 分为肌棱柱层(myostracum)、斜棱柱层(fibrous prismatic)、复杂交叉状层(complex crossed lamellar)、交叉板状层 intersected crossed platy)、珍珠质层(nacre)、球棱柱层(spherulitic prismatic)、均质层(homogeneous)等7大类。但由于贝类种类、生长环境的不同, 造成贝壳结构、化学成分和性能上的差异很大[25-27]。贝壳主要成分包括95%的碳酸钙质晶体和少量有机质(约占5%)[28]。碳酸钙晶体构型主要有文石质、方解石和混合质3种形式, 其中文石的力学性能优于方解石, 但方解石的稳定性好, 在混合质壳体贝类(牡蛎、贻贝等)中, 位于外层的方解石能有效防止碳酸钙溶解, 保护贝壳和适应复杂多变的水体环境[29]。另外, 贝壳的形成是一个有机相-无机相分子相互作用的过程[30], 有机质在贝壳的形成以及结构、性能方面起到至关重要的作用, 它通过控制晶体形成、晶体结构、物相转换等影响贝壳的性能[31]。因此充分了解不同贝类贝壳的结构和成分组成能针对性地对贝壳性状进行开发利用。
菲律宾蛤仔、文蛤、厚壳贻贝()、泥蚶()和缢蛏()均是东部沿海习见的海洋贝类, 具有重要的经济价值。本实验用扫描电镜技术、X-射线粉末衍射技术和弱酸去钙分析法对5种贝类贝壳的断面微观结构、物相组成、有机基质和蛋白质含量等进行了观察和比较分析, 以期为深入研究贝壳的形成机理及贝壳性状的开发利用积累基础资料。
1 材料与方法
1.1 贝壳样品处理
实验用菲律宾蛤仔、缢蛏为1龄贝, 文蛤为2龄贝, 泥蚶为3龄贝, 均采自于浙江省宁波市海洋与渔业研究院科技创新基地的养殖池塘, 厚壳贻贝为2龄贝, 采自浙江舟山。各取成贝3粒, 解剖去除软体部, 用5% NaOH溶液去除贝壳上附着的杂质, 并用去离子水清洗贝壳, 晾干备用。
1.2 贝壳结构的扫描电镜观察
选取贝壳中间部位制成0.5 cm×0.5 cm小块(图1), 断面用去离子水清洗后, 置于超声波清洗仪处理10 min以清洁断面。利用日本日立公司SU8230型扫描电镜观察拍照。
1.3 贝壳成分的X射线分析
将贝壳置于105 ℃烘箱中恒温烘干3 h后, 用小型粉碎机粉碎, 过0.15 mm筛, 得到待测样品。采用德国布鲁公司D8 Advance型多晶体X射线衍射仪进行检测, 检测条件为: CuKα辐射、波长0.154 nm、电压40 kV、扫描范围3°~60°、步长0.02°、步速0.2 s。采用绝热法计算各种晶体类型的比例。
W=/[K(1/1+2/2+……I/K)][32]. (1)
用待测物质与纯刚玉(αAl2O3)按重量比1∶1混合均匀, 测量两者的衍射强度之比, 这个比值称为这个物相的值。式中代表样品中任一相,和分别指衍射强度和质量分数。
1.4 有机质提取和蛋白含量测定
将贝壳用小型粉碎机粉碎, 取样品各5 g, 用6.5 mol/L的醋酸于冰浴条件下溶解, 溶解过程尽量避免CO2过快生成和溢出, 反应至完全无气泡后终止。将混合物离心(4 000 r/min, 4 ℃, 10 min)后分别收集上清液和沉淀, 上清液装入截留分子量为1 000 Da的透析袋透析, 每隔半天换水一次, 透析共计3 d, 整个透析过程温度维持在4 ℃。收集已透析的样品溶液, 经 0.22 μm微孔滤膜过滤后于–20 ℃至完全冻结, 冷冻干燥后收集样品, 得到可溶有机质(soluble matrix, SM)。将离心后的沉淀用去离子水冲洗3次, 离心后冷冻干燥, 收集样品得到不可溶有机质(insoluble matrix, IM)。称重并计算可溶性有机质和不可溶性有机质的产率。将冷冻干燥后的上清和沉淀样品溶解, 采用二喹啉甲酸蛋白质法测定蛋白含量。
2 结果与分析
2.1 5种贝壳的微观结构
菲律宾蛤仔、文蛤、厚壳贻贝、泥蚶、缢蛏5种贝壳的厚度分别为(1.03±0.08) mm、(1.36±0.14) mm、(1.15±0.09) mm、(2.64±0.18) mm、(0.28±0.04) mm, 均由角质层、棱柱层和珍珠层3层组成。
贝壳断面结构扫描电镜结果显示, 5种贝壳中壳层[33]主要包括斜棱柱层、棱柱层和球棱柱层3类层次结构。斜棱柱层在菲律宾蛤仔、厚壳贻贝、缢蛏中均存在, 菲律宾蛤仔斜棱柱层位于角质层下方, 晶体呈柱状且不同晶体长度有明显的差异, 晶体排列方向朝向贝壳生长方向(图1a); 厚壳贻贝斜棱柱层呈圆柱状, 与菲律宾蛤仔斜棱柱层相比较规律有序, 晶体厚度约为1 μm, 长度不均, 最长从珍珠层到角质层, 随贝壳大小变化, 晶体排列紧密(图1g); 缢蛏斜棱柱层呈“M”字结构从左到右重复排列, 有圆形颗粒晶体依附在柱状晶体上, 排列紧密厚实, 构成了大部分贝壳(图1m)。棱柱层在文蛤、泥蚶中存在, 文蛤棱柱层表现为由垂直于贝壳表面的短柱状晶体组成平行于贝壳表面的方块状晶体, 厚度约为0.3~0.4 μm(图1d); 泥蚶棱柱层则表现为柱状晶体紧密堆叠, 平行于贝壳表面, 晶体排列比文蛤更为紧密(图1j)。球棱柱层只存在于菲律宾蛤仔中, 其结构由大小较均匀的小圆块紧密组合而成, 排列紧密(图1b)。另外, 在泥蚶的棱柱层下方分别存在有均质层和交错板状层。均质层表现为表面光亮并有不规则的晶体突起, 晶体密实, 整体质地均匀, 可观察到晶体表面分布有丝状有机物(图1k); 交错片状层结构是由互相平行的文石晶体聚集而成, 首级片体与次级片体互相部分叠加, 首级片体长而且规则, 内部次级片体较短且不规则, 分布广泛, 为贝壳中间层主要结构, 这种复杂交错片状排列方式可以增加贝壳的强度(图1l)。

图1 贝壳断面微观结构扫描电镜图
注: a—c: 菲律宾蛤仔的斜棱柱层、球棱柱层和珍珠层; d—f: 文蛤的棱柱层、肌棱柱层和珍珠层; g—i: 厚壳贻贝的斜棱柱层、肌棱柱层和珍珠层; j—l, o: 泥蚶的棱柱层、均质层、交错板状层、和珍珠层; m, n: 缢蛏的棱柱层和珍珠层
5种贝壳的珍珠层主要包括不规则状、复杂交叉状、“板砖”状3种类型的层次结构(表1)。菲律宾蛤仔、文蛤、缢蛏珍珠层呈不规则结构, 表现为由密实的块状晶体镶嵌而成, 缝隙有不规则的颗粒晶体填充, 晶体排列较紧密, 排列紧密程度由大到小为: 文蛤、缢蛏、菲律宾蛤仔, 同时观察到块状晶体之间存在丝状有机物连接(图1c、f、n)。泥蚶珍珠层呈复杂交叉状, 首级片体形状不规则, 相邻块体中的板片主要在两个相反的方向排列, 该结构层相对于交错片状结构而言, 有序性较小, 厚度约为22 μm(图1o)。厚壳贻贝的珍珠层呈典型的“砖-泥”结构, 由许多整齐堆叠的片层构成, 具有明显的层状结构, 片层厚度约0.4~0.6 μm, 在片层结构之间存在丝状有机物连接, 与前4种的珍珠层相比, 显得更加有序规则(图1i)。在5种贝类贝壳中, 发现文蛤和厚壳贻贝珍珠层中存在肌棱柱层, 文蛤肌棱柱层呈方柱状结构, 包埋在珍珠层内并垂直贝壳表面, 厚度1~2 μm (图1e)。厚壳贻贝肌棱柱层同样包埋在珍珠层中, 呈垂直于贝壳表面的柱状结构, 紧密排列形成厚度约为1 μm板片并平行于贝壳表面, 整个肌棱柱层结构如文蛤一样, 晶体排列紧密并平行于贝壳表面(图1h)。

表1 5种贝类贝壳断面的微观结构特征
2.2 贝壳的X-射线粉末衍射和物相组成
图2是5种贝类贝壳粉末的XRD图谱, 其中菲律宾蛤仔、文蛤、泥蚶、缢蛏的图谱类似, 峰形尖锐、结晶度良好, 均在(111)、(021)、(012)、(200)、(112)、(221)、(202)等晶面出现了与标准文石碳酸钙晶体(PDF#41-1475)基本吻合的特征衍射峰, 表明这4种贝壳的碳酸钙晶体基本为文石晶体。厚壳贻贝图谱在(104)、(110)、(113)、(202)等晶面与标准方解石碳酸钙晶体(PDF#47-1743)的特征衍射峰基本吻合, 表明其贝壳碳酸钙晶体主要以方解石晶型为主。
根据公式(1)计算得出, 菲律宾蛤仔贝壳无机相由98.9%文石和1.1%方解石组成, 文蛤、泥蚶贝壳无机相由100%文石组成, 缢蛏由98.6%文石和1.4%方解石组成, 而厚壳贻贝则由30.1%文石和69.9%的方解石组成(图2, 表2)。
2.3 贝壳有机质及蛋白含量的比较
用酸作为脱钙溶液来分离可溶有机质和不可溶有机质, 5种贝类贝壳样品中提取的有机质产率见表3。菲律宾蛤仔、文蛤、厚壳贻贝、泥蚶、缢蛏的有机质含量分别为3.283%、3.429%、3.057%、2.461%、3.345%, 不可溶有机质含量较高。

图2 五种贝壳的X射线粉末衍射图
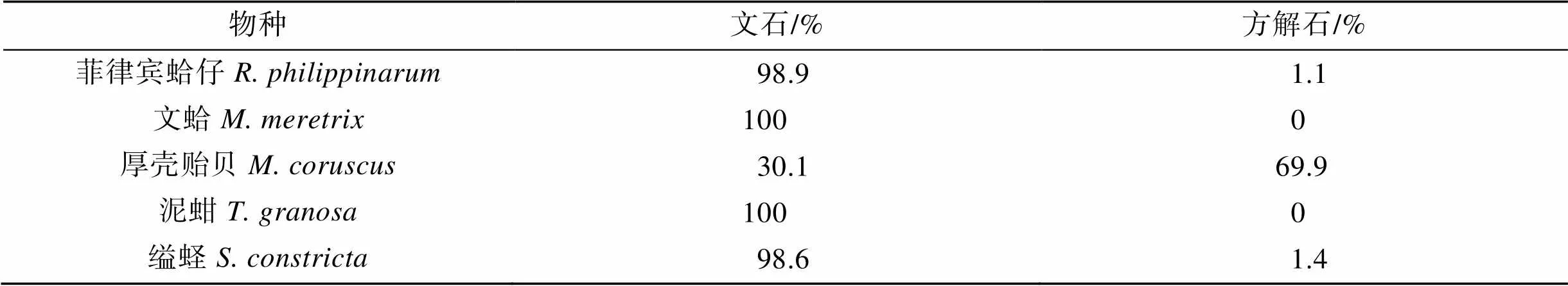
表2 五种贝类贝壳物相组成
通过二喹啉甲酸蛋白质法测定有机质中的蛋白质含量, 结果显示, 菲律宾蛤仔、文蛤、厚壳贻贝、泥蚶、缢蛏贝壳有机质中蛋白质的相对含量分别为2.98%、3.543%、4.954%、7.21%、5.086%, 可溶性蛋白含量是不可溶蛋白含量的5.55~20.31倍(表3)。
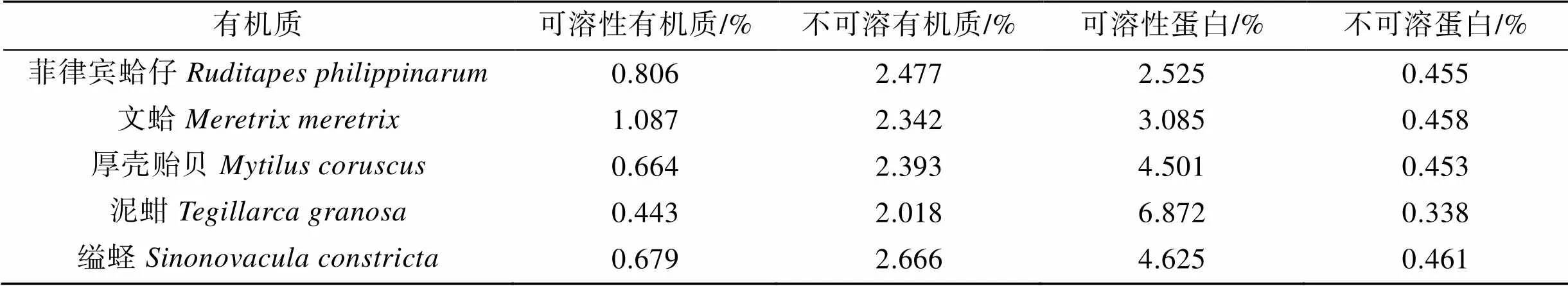
表3 五种贝类贝壳有机质含量比较
3 讨论
贝壳的微观结构有着明显的多样性, 如同为帘蛤科(Veneridae)的菲律宾蛤仔和文蛤却在贝壳中间层组成上存在较大差异, 菲律宾蛤仔的中间层为短柱状的斜棱柱层和颗粒状的球棱柱层, 斜棱柱层晶体排列朝向贝壳生长方向, 球棱柱平行于贝壳开口方向, 而文蛤的中间层为平行于开口方向的短柱状棱柱层。这些差异主要表现在晶体的组成和排列结构两个方面, 这与陈道海等[34]关于13种双壳类贝壳的扫描电镜观察结果一致, 不同种双壳贝类贝壳横切面微观结构均有一定的差异。厚壳贻贝[28]和绿贻贝()[2]均属于贻贝科, 但二者贝壳断面结构也存在明显的差异, 厚壳贻贝断面结构主要包含珍珠质层、肌棱柱层和斜棱柱层3种层次结构, 而绿贻贝仅包含珍珠质层和肌棱柱层2种, 不存在斜棱柱层。在池蝶蚌()[35]中, 贝壳及其珍珠的珍珠层表面出现螺旋状纹理, 这在其他的淡水珍珠贝(如三角帆蚌)的贝壳及珍珠中极为少见。另外, 研究发现泥蚶贝壳断面结构比菲律宾蛤仔、文蛤、缢蛏贝壳更为复杂, 包含棱柱层、均质层、交错板状层、斜棱柱层和珍珠层5种类型的层次结构, 与虾夷扇贝()贝壳层次结构相似[36], 其复杂的层次结构可能赋予了贝壳更强的抗击能力和抗磨损能力。
贝壳的无机相组成与生活环境及生活方式有关。文蛤、缢蛏、泥蚶、菲律宾蛤仔属于埋栖型贝类, 生活在潮间带滩涂或池塘的沙泥底质中, 壳体被底质覆盖, 可以做短距离运动, 如文蛤具有随着生长由中潮区向低潮区或潮下带迁移的习性, 缢蛏可在洞穴内做升降运动等, 需要坚硬的贝壳来抵抗底质和周围环境的摩擦力。相对坚硬的贝壳更利于抵抗周围环境的摩檫力。Fan[29]指出, 处在泥沙之中, 周围介质(主要指孔隙水)的理化变化相对平稳, 对壳体稳定性的要求相对降低, 从文石、方解石性质差别上分析, 文石更适合这样的环境。且文石的硬度比方解石大, 所以4种埋栖贝类贝壳以文石质壳体为主。而厚壳贻贝是附着型贝类, 与扇贝的生活方式相同, 贝壳属于混合质壳体。Fan[29]认为混合质壳既能节省壳体需要付出的生物能量, 又具有方解石质壳体的性能, 适应复杂的水体变化。另有研究表明[37], 柳珊瑚全轴亚目(Holaxonia)的基部和轴, 如果温度在15 ℃以上全为方解石, 20 ℃以上则全是文石质, 在15~20 ℃之间为文石和方解石组成, 这也解释了贝壳物相的组成与其生活环境、生活方式有关。
贝壳的形成是一个有机相-无机相分子相互作用的过程[30], 其中有机质在贝壳形成以及结构、性能方面起到至关重要的作用。有机质通过控制晶体形成、晶体结构、物相转换等影响贝壳的性能。研究发现, 有机质在贝壳中的百分含量随贝壳种类不同而不同, 扇贝和珠母贝贝壳的有机质含量分别为4.36%和5.24%[38]; 三角帆蚌与褶纹冠蚌贝壳粉平均有机质含量为0.903%, 蛋白质含量为1.64%[39]; 而香螺()[40]贝壳中的蛋白含量约为0.943%。本实验中, 5种贝类贝壳有机质含量均为3%左右, 文蛤有机质含量最高, 占壳重的3.429%, 泥蚶有机质含量最低, 占壳重的2.461%; 其中蛋白质的相对含量在2.98%~7.21%, 泥蚶蛋白质含量最高, 占有机质的7.21%, 菲律宾蛤仔含量最低, 占2.98%。不同贝类贝壳或者同一贝类在不同环境、不同生长期、不同壳层, 其有机质的含量均有差异[41]。
4 结论
贝壳是绝大多数贝类防御保护的特殊结构, 而目前有关贝壳结构和成分的系统研究较为欠缺。本研究综合应用扫描电镜技术、X-射线粉末衍射技术和弱酸去钙分析法对中国5种习见经济贝类贝壳的断面微观结构、物相组成、有机基质和蛋白质含量进行了系统观察和比较分析, 主要结论如下:
1) 它们的微观结构不尽相同, 差异主要表现在晶体的组成和排列结构;
2) 它们的物相组成均由约97%的碳酸钙和约3%的有机质组成;
3) 菲律宾蛤仔、文蛤、泥蚶和缢蛏4种贝壳无机相几乎由文石组成, 而厚壳贻贝贝壳由文石和方解石组成;
4) 不同种类贝壳中总蛋白质含量差异较大, 可溶性蛋白含量显著高于不可溶蛋白。
[1] 廖智, 孙琦, 姜雨婷. 贝类闭壳肌-贝壳连接界面的分子组成及连接机制[J]. 浙江海洋大学学报(自然科学版), 2018, 37(4): 313-319.
LIAO Zhi, SUN Qi, JIANG Yuting. Molecular composition and mechanism of muscle-shell attachment of shellfish[J]. Journal of Zhejiang Ocean University (Natural Science), 2018, 37(4): 313-319.
[2] 鲍林飞, 高鹏, 赵鲁苹, 等. 绿贻贝贝壳的微结构特征及光谱分析[J]. 浙江海洋学院学报(自然科学版), 2014, 33(4): 347-353.
BAO Linfei, GAO Peng, ZHAO Luping, et al. Microstructural characteristics and FTIR analysis of the shell from green mussel ()[J]. Journal of Zhejiang Ocean University (Natural Science), 2014, 33(4): 347-353.
[3] 王伟, 唐玉卿. 杂交栉孔扇贝试养技术[J]. 水产科学, 2003, 22(2): 37.
WANG Wei, TANG Yuqing. Trial on hybridizedculture[J]. Fisheries Science, 2003, 22(2): 37.
[4] 许飞, 郑怀平, 张海滨, 等. 海湾扇贝“中科红”品种与普通养殖群体不同温度下早期性状的比较[J]. 水产学报, 2008, 32(6): 876-883.
XU Fei, ZHENG Huaiping, ZHANG Haibin, et al. Comparison on the larval traits between strain “Zhongkehong” and common cultured population of bay scallopunder different temperature[J]. Journal of Fisheries of China, 2008, 32(6): 876-883.
[5] 李琪, 于瑞海, 孔令锋, 等. 长牡蛎“海大2号”[J]. 中国水产, 2018(6): 84-92.
LI Qi, YU Ruihai, KONG Lingfeng, et al.“Haida No. 2”[J]. China Fisheries, 2018(6): 84-92.
[6] 李琪, 于瑞海, 孔令锋, 等. 长牡蛎“海大3号”[J]. 中国水产, 2020(4): 98-105.
LI Qi, YU Ruihai, KONG Lingfeng, et al.“Haida No. 3”[J]. China Fisheries, 2020(4): 98- 105.
[7] 刘保忠, 柴雪良. 文蛤“科浙1号”[J]. 中国水产, 2014(10): 48-50.
LIU Baozhong, CHAI Xueliang.“Kezhe No. 1”[J]. China Fisheries, 2014(10): 48-50.
[8] 林志华, 董迎辉. 文蛤“万里红”[J]. 中国水产, 2015(10): 72-74.
LIN Zhihua, DONG Yinghui.“Wanlihong”[J]. China Fisheries, 2015(10): 72-74.
[9] 张兴志, 张继彪, 刘辉, 等. 菲律宾蛤仔獐子岛群体两种壳型表型性状比较[J]. 水产科学, 2015, 34(10): 616-620.
ZHANG Xingzhi, ZHANG Jibiao, LIU Hui, et al. Comparisons of phenotypic traits in two shell type strains of manila clam in Zhangzi Island[J]. Fisheries Science, 2015, 34(10): 616-620.
[10] 莫日馆, 肖述, 秦艳平, 等. 深凹壳型香港牡蛎家系生长与存活性状比较[J]. 中国水产科学, 2019, 26(5): 869-882.
MO Riguan, XIAO Shu, QIN Yanping, et al. Estimates of genetic parameters of growth-related traits for the selective breeding of deep-cupped shell[J].Journal of Fishery Sciences of China, 2019, 26(5): 869-882.
[11] ALLEN S K, GAFFNEY P M, EWART J W. Genetic improvement of the eastern oyster for growth and disease resistance in the northeast[R]. NRAC fact sheet No. 210, College Park, MD: Norrheastern Regional Aquaculture Center, 1993: 2.
[12] 马玉菲, 乔莉, 冯庆玲. 淡水珍珠的生物矿化机理研究进展[J]. 无机材料学报, 2013, 28(1): 109-116.
MA Yufei, QIAO Li, FENG Qingling. Research progress on biomineralization mechanism of freshwater pearl[J]. Journal of Inorganic Materials, 2013, 28(1): 109-116.
[13] 战欣, 陈琼, 顾志峰, 等. 马氏珠母贝贝壳珍珠质层厚度性状与微卫星标记的相关性分析[J]. 热带生物学报, 2016, 7(3): 290-295.
ZHAN Xin, CHEN Qiong, GU Zhifeng, et al. Analysis of correlationship between the thickness of shell inner nacreous layer and microsatellite markers in[J]. Journal of Tropical Biology, 2016, 7(3): 290-295.
[14] 张伟钢, 汪港, 唐建, 等. 贝壳珍珠层动态的文石板片厚度及其新结构模型[J]. 广西大学学报(自然科学版), 2009, 34(4): 509-512.
ZHANG Weigang, WANG Gang, TANG Jian, et al. Dynamic aragonite sheets thickness and new structural model of shell nacre[J]. Journal of Guangxi University (Natural Science Edition), 2009, 34(4): 509-512.
[15] GUIÑEZ R, TORO J E, KRAPIVKA S, et al. Heritabilities and genetic correlation of shell thickness and shell length growth in a mussel,(Bivalvia: Mytilidae)[J]. Aquaculture Research, 2017, 48(4): 1-8.
[16] GREFSRUD E S, STRAND I. Comparison of shell strength in wild and cultured scallops ()[J]. Aquaculture, 2006, 251(2/4): 306-313.
[17] NAGARAJAN R, LEA S E G, GOSS-CUSTARD J D. Seasonal variations in mussel,L. shell thickness and strength and their ecological implications[J]. Journal of Experimental Marine Biology & Ecology, 2006, 339(2): 241-250.
[18] 谢忠东, 丁晓非, 李凤敏, 等. 3种海产螺壳体相组成及结构特征分析[J]. 水产科学, 2006, 25(5): 253-255.
XIE Zhongdong, DING Xiaofei, LI Fengmin, et al. The structure characteristics of the shell of the conch[J]. Fisheries Science, 2006, 25(5): 253-255.
[19] 陈静, 黄根哲. 鲍鱼壳、香螺壳的结构及力学性能[J]. 机械工程师, 2016, 7(3): 39-41.
CHEN Jing, HUANG Genzhe. The structure and mechanical properties of abalone shell andshell[J]. Mechanical Engineer, 2016, 7(3): 39-41.
[20] FENG Q L, CUI F Z, PU Get al. Crystal orientation, toughening mechanisms and a mimic of nacre[J]. Materials Ence and Engineering: C, 2000, 11(1): 19-25.
[21] MURR L E, RAMIREZ D A. The microstructure of the cultured freshwater pearl[J]. Jom: The Journal of the Minerals, Metals & Materials Society, 2012, 64(4): 469-474.
[22] 严俊, 方飚, 张姗姗, 等. 双壳纲三角帆蚌贝壳的三维微结构及其化学组成研究[J]. 材料导报, 2013, 27(16): 108-112.
YAN Jun, FANG Biao, ZHANG Shanshan, et al. Study on three-dimensional microstructure and chemical composition of bivalve shells of(Lea)[J]. Materials Reports, 2013, 27(16): 108-112.
[23] 吴小平, 欧阳珊, 梁彦龄, 等. 蚌科贝壳的扫描电镜观察[J]. 南昌大学学报(理科版), 1999, 23(1): 58-61.
WU Xiaoping, OU Yangshan, LIANG Yanling, et al. A scanning electron microscopic study of the shells of unionidae[J]. Journal of Nanchang University (Natural Science), 1999, 23(1): 58-61.
[24] CARTER J G. Shell microstructural data for the bivalvia[M]. New York: Van Nostrand Reinhold and Co, 1990.
[25] 唐亚丽, 张恩. 粉红贝贝壳的结构特征及其颜色[J]. 江苏农业科学, 2017, 45(24): 156-161.
TANG Yali, ZHANG En. Structural property and coloring mechanism of pink shellfish[J]. Jiangsu Agricultural Sciences, 2017, 45(24): 156-161.
[26] LOWENSTAM H A. Factors affecting the aragonite: calcite ratios in carbonate-secreting marine organisms[J]. Journal of Geology, 1954, 62(3): 284-322.
[27] WALTER L M, MORSE J W. Reactive surface area of skeletal carbonates during dissolution: effect of grain size[J]. Journal of Sedimentary Research, 1984, 54: 1081-1090.
[28] 赵鲁苹, 徐焕志, 陈东, 等. 厚壳贻贝贝壳的微结构及光谱分析[J]. 浙江大学学报(理学版), 2015, 42(3): 339-346.
ZHAO Luping, XU Huanzhi, CHEN Dong, et al. Microstructure and spectral analysis ofshell[J]. Journal of Zhejiang University (Science Edition), 2015, 42(3): 339-346.
[29] FAN D J. Comparison study on the shell mineralogy of 9 species of modern bivalves living in yellow sea[J]. Acta Sedimentologica Sinica, 2005, 23(3): 475-482.
[30] MANN S. Molecular recognition in biomineralization[J]. Nature, 1988, 332(6160): 119-124.
[31] 张文兵, 姚春凤, 麦康森. 贝壳生物矿化的研究进展[J]. 海洋科学, 2008, 32(2): 74-79.
ZHANG Wenbing, YAO Chunfeng, MAI Kangsen. Progress in the studies on shell biomineralization[J]. Marine Sciences, 2008, 32(2): 74-79.
[32] CHUNG F H. Quantitative interpretation of X-ray diffraction patterns of mixtures. II. Adiabatic principle of X- ray diffraction analysis of mixtures[J]. Journal of Applied Crystallography, 1974, 7(6): 526-531.
[33] 庞云龙, 丁君, 田莹, 等. 不同生长时期虾夷扇贝壳质的超微结构观察及表面5种元素组成分析[J]. 海洋科学, 2015, 39(8): 28-34.
PANG Yunlong, DING Jun, TIAN Ying, et al. Analysis of shell microstructure and five surface elements ofat different ages[J]. Marine Sciences, 2015, 39(8): 28-34.
[34] 陈道海, 霍颖娴. 13种双壳类贝壳的扫描电镜观察[J]. 动物学杂志, 2015, 50(1): 122-130.
CHEN Daohai, HUO Yingxian. Shell microstructure in thirteen species of bivalves[J]. Chinese Journal of Zoology, 2015, 50(1): 122-130.
[35] 罗琴凤, 亓利剑, 尹作为. 池蝶蚌贝壳及其珍珠的微结构表征[J]. 宝石和宝石学杂志, 2008, 10(2): 17-20.
LUO Qinfeng, QI Lijian, YIN Zuowei. Microstructure ofshells and its pearls[J]. Journal of Gems and Gemmology, 2008, 10(2): 17-20.
[36] LIN A G, DING X F, XIE Z D, et al. Microstructure of(scallop) shell and correlations with functions[J]. Journal of the Chinese Ceramic Society, 2010, 38(3): 504-509.
[37] 梁艳. 几种腹足纲贝壳的结构和性能[D]. 大连: 大连理工大学, 2008.
LIANG Yan. The microstructure and properties of several Gastropoda shells[D]. Dalian: Dalian University of Technology, 2008.
[38] 何朋, 陈建新, 苏敏, 等. 贝壳的化学成分及其结构特征[J]. 化工学报, 2015, 66(S2): 450-454.
HE Peng, CHEN Jianxin, SU Min, et al. Analysis of chemical composition and structure characteristics of shells[J]. CIESC Journal, 2015, 66(S2): 450-454.
[39] 夏静芬, 钱国英, 陈亮, 等. 珍珠粉和贝壳粉的化学成分和结构特征分析[J]. 化学研究与应用, 2010, 22(11): 1467-1471.
XIA Jingfen, QIAN Guoying, CHEN Liang, et al. Analysis of chemical composition and structure characteristics of pearl and conch powders[J]. Chemical Research and Application, 2010, 22(11): 1467-1471.
[40] 梁艳, 赵杰, 王来, 等. 贝壳的有机质及生物矿化机制分析[J]. 生物学杂志, 2006, 23(6): 19-23.
LIANG Yan, ZHAO Jie, WANG Lai, et al. The organic matrix and analysis of biomineralization mechanism of mollusk shell[J]. Journal of Biology, 2006, 23(6): 19-23.
[41] 姜国良, 陈丽, 刘云. 贝壳有机基质与生物矿化[J]. 海洋科学, 2002, 26(2): 17-19.
JIANG Guoliang, CHEN Li, LIU Yun. The organic matrix of molluscan shell and its relevance[J]. Marine Sciences, 2002, 26(2): 17-19.
Microstructure and composition analysis of five species of economic bivalves
MO Tian-bao1, 2, XU Hong-qiang3, HE Jing3, DONG Ying-hui2, LIN Zhi-hua2, 3
(1. National Demonstration Center for Experimental Fisheries Science Education, Shanghai Ocean University, Shanghai 201306, China; 2. Zhejiang Key Laboratory of Aquatic Germplasm Resources, College of Biological & Environmental Sciences, Zhejiang Wanli University, Ningbo 315100, China; 3. Ninghai Marine Biological Seed Industry Research Institute, Zhejiang Wanli University, Ninghai 315604, China)
Five popular species of marine bivalves,,,,, and, living off the east coast of China, were selected for shell structure characterization and chemical composition analyses using scanning electron microscopy, X-ray diffraction, and decalcifying in a weak acid. Scanning electron microscopy results showed that the shells comprised a prismatic layer and nacre layers. Additionally, the prismatic crystal structure included five types: fibrous prismatic, spherical prismatic, prismatic, homogeneous, and crossed lamellar layers.had only a fibrous prismatic layer, whilehad all types of layers except a spherical prismatic layer. These differences might be related to shell-crushing resistance. The nacre crystal structure contains nacreous and myostracum layers. The nacreous layer ofshowed an obvious layered structure that was a typical brick-mud structure, while the other four shells presented an irregular blocky structure. The X-ray powder diffractometry results showed two types of shells in the five mollusks, namely aragonite shells and calcite shells. In four species,,,, and, the inorganic phases were almost entirely composed of aragonite, while theinorganic phase was composed of aragonite and calcite. The organic matrix content in the five bivalves was approximately 3%, and the total protein content accounted for 2.98%–7.21% of the organic matrix, among which the content of soluble protein was 5.55–20.31 times that of insoluble protein. The information obtained from this study will provide a reliable scientific basis for biomineralization.
bivalves; shell; organic matter; SEM; XRD
Dec. 17, 2020
S91
A
1000-3096(2022)12-0041-09
10.11759/hykx20201217003
2020-12-17;
2021-03-03
国家重点研发计划“蓝色粮仓科技创新”专项课题(2018YFD0901405); 浙江省农业新品种选育重大科技专项课题(2021C02069-7); 宁波市重大科技攻关暨“揭榜挂帅”项目(2021Z114); 国家海洋水产种质资源库项目
[National Key Research and Development Program of China, No. 2018YFD0901405; Zhejiang Major Program of Science and Technology, No. 2021C02069-7; Ningbo Major Project of Science and Technology, No. 2021Z114; National Marine Genetic Resource Center Program]
莫天宝(1995—), 男, 广西玉林人, 硕士研究生, 主要从事贝类遗传育种研究, E-mail: 191950530@qq.com; 林志华(1965—),通信作者, 研究员, E-mail: zhihua9988@126.com; 董迎辉(1980—), 通信作者, 教授, E-mail: dongyinghui118@126.com
(本文编辑: 杨 悦)
