野生二粒小麦PYL基因家族的鉴定及其在逆境胁迫下的表达特性分析
2022-03-01王振宇潘文秋宋卫宁聂小军
王振宇,杨 光,赛 娜,潘文秋,宋卫宁,2,聂小军,2
(1.西北农林科技大学农学院,陕西杨凌 712100; 2.旱区作物逆境生物学国家重点实验室,陕西杨凌 712100)
脱落酸(ABA)在植物生长发育中发挥着关键作用,参与细胞伸长和分裂、胚成熟、种子脱水耐性、种子休眠和萌发、叶片衰老以及诱导根生长和果实成熟等过程。此外,ABA还可以调节气孔孔径,在调控植物响应各种生物和非生物胁迫中具有不可或缺的重要作用。[()-]是目前已知的最大的植物激素受体基因家族。Park等利用种子萌发抑制剂Pyrabactin处理拟南芥,首次筛选出了抗Pyrabactin的突变株,并从突变株中分离得到可特异结合ABA的基因及其他13个的同源基因,将其分别命名为~。Ma等研究发现,PYL家族成员可以结合ABA,抑制A类PP2C蛋白磷酸酶ABI1或者ABI2的活性。PYL蛋白作为ABA的受体,是ABA信号通路最上游的调控因子,承担着识别ABA信号和启动信号转导的作用。
鉴于基因家族的重要功能,前人对拟南芥()、水稻(L.)、玉米(L.)、大豆()、甘蓝型油菜(L.)、烟草(L.)和陆地棉(spp.)中的基因家族已经进行了全基因组鉴定,部分基因的功能也已经被验证。如在拟南芥中,、、、、、和可促进ABA诱导的种子萌发、气孔关闭和根系生长,而和则抑制种子萌发。另外,前人还发现和与拟南芥的耐旱性相关;过表达可以增强拟南芥的耐旱性。在水稻中,过表达可促进种子萌发、幼苗生长,并增强水稻对干旱和盐胁迫耐受性;和在水稻胚乳中特异性表达,并在水稻种子萌发时促进其对ABA的敏感性。在小麦中,基因在ABA和盐胁迫下上调表达。这些结果都表明,基因能够广泛参与植物生长发育的调控以及逆境胁迫的响应。
野生二粒小麦(,2=4=28,AABB)是普通小麦(2=6=42,AABBDD)和栽培四倍体小麦(硬粒小麦)(2=4=28,AABB)A、B染色体组的祖先供体种,具有遗传资源丰富、粒大、蛋白质含量高、抗病、耐逆、耐贫瘠等多种优良特性,一直是小麦遗传改良的重要基因库。但截至目前,有关野生二粒小麦基因的研究还未见报道,这限制了野生二粒小麦基因生物学功能的深入研究。野生二粒小麦参考基因组的破译,使得在全基因组水平鉴定和分析其基因家族的组成和特征成为可能。鉴于此,本研究利用最新的野生二粒小麦参考基因组信息,基于全基因组搜索的方法,对野生二粒小麦基因家族在全基因组水平上进行了鉴定和分析,并进一步对其基因结构、保守结构域、启动子顺式作用元件以及不同组织和逆境胁迫下的表达特性进行系统分析,以期为研究野生二粒小麦基因的功能以及了解ABA信号转导通路提供有益信息。
1 材料与方法
1.1 野生小麦PYL基因家族成员的鉴定
首先,从Ensembl Plants数据库(http://plants.ensembl.org/index.html)中下载野生二粒小麦参考基因组注释信息及其所有蛋白序列,将下载到的蛋白序列构建本地蛋白库,从拟南芥基因组数据库(TAIR, https://www.arabidopsis.org/browse/genefamily/index.jsp)下载得到的拟南芥14条PYL蛋白序列作为query序列,采用BLASTP工具进行blast比对,参数设置为1e;同时,从PFAM数据库(http://pfam.xfam.org/)下载基因家族的profile模型(PF10604)作为搜索模型,利用HMM 3.0软件筛选含有该结构域的蛋白序列;然后,将上述两种方法筛选到的候选蛋白合并,用DNAMAN 5.0软件进行多序列比对,手工去除不完整读码框序列和冗余序列;最后,将得到的蛋白序列提交到PFAM及NCBI-CDD在线工具检测其保守蛋白结构域,只保留含有完整PYR-PYL-RCAR-like功能域的序列作为最终的野生二粒小麦基因;利用ExPASy(https://www.expasy.org/)在线软件对野生二粒小麦PYL蛋白的分子量、氨基酸长度和等电点进行预测,利用CELLO软件进行亚细胞定位。
1.2 系统发育树构建、基因结构和蛋白结构域 分析
从拟南芥基因组数据库(https://www.arabidopsis.org/browse/genefamily/index.jsp )和水稻基因组数据库(http://rice.plantbiology.msu.edu/)中检索并下载拟南芥和水稻基因序列,利用Clustal_W工具将野生二粒小麦、水稻和拟南芥基因序列进行多序列比对,利用MEGA 8.0软件的邻接法(NJ)构建系统发育树,Bootstrap参数设置为1 000;根据野生二粒小麦基因组gtf注释信息,获得各个野生二粒小麦候选基因的外显子-内含子结构信息,并利用在线软件GSDS 2.0(http://gsds.cbi.pku.edu.cn/)绘制示意图;采用MEME工具(http://meme-suite.org/)预测各野生二粒小麦PYL蛋白的保守结构域,并利用DNAMAN 5.0软件进行结构域序列多重比对,比对参数设为默认值。
1.3 顺式作用元件分析
利用TBtools工具(https://github.com/CJ-Chen/TBtools/)截取各个野生二粒小麦基因上游2 kb的基因组序列,然后递交到启动子预测数据库PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)进行启动子顺式作用元件预测并手工整理,并对预测到的顺式作用元件进行图形化展示。
1.4 野生二粒小麦PYL基因的组织表达模式及和逆境胁迫表达分析
从NCBI SRA数据库(https://www.ncbi.nlm.nih.gov/sra)下载野生二粒小麦不同组织以及干旱胁迫下野生二粒小麦品系TR39477和TTD22的RNA-seq数据,同时利用本实验室保留的耐盐品种A5盐胁迫下不同时间点的RNA-seq数据;采用Hisat 2和StringTie软件将RNA-seq原始reads与参考基因组进行映射(mapping),并计算各个基因的FPKM值,然后提取所有24个基因在不同样本中的FPKM值,并利用Deseq2鉴定差异表达基因,利用R软件绘制表达谱热图。
1.5 PYL基因在四倍体小麦中的单倍型分析
从GSA数据库中下载所有四倍体小麦重测序的原始测序数据(https://bigd.big.ac.cn/gsa)(CRA001951)和SNP遗传变异的VCF文件(https://bigd.big.ac.cn/gvm)(GVM000082)。重测序样本共包含有28份野生二粒小麦、29份栽培二粒小麦和13份硬粒小麦材料;根据野生二粒小麦基因的染色体位置,从VCF文件中提取每个基因的SNP,并计算群体中每个基因的单倍型及其数量,将群体中50%以上样品具有的单倍型定义为主要单倍型,并比较三个群体中主要单倍型的频率和比例。
2 结果与分析
2.1 野生二粒小麦PYL基因家族鉴定结果
利用BLASTP比对和HMM比对两种方法,从野生二粒小麦基因组中共鉴定到26个候选基因,进一步对保守结构域和基因结构验证发现,有2个基因的保守结构域不完整,剔除后最终得到24个包含有完整特征结构域的野生二粒小麦基因。这24个基因不均匀分布在野生二粒小麦1A、1B、2A、2B、3A、3B、4A、4B、7A 和7B染色体上,其中3号染色体组最多,3A和3B染色体上均具有5个,其次是1B染色体,有4个基因;5号和6号染色体组上没有鉴定到基因(表1)。共发现有10组同源基因,即A和B同源拷贝均存在,而剩下的4个基因发生了同源基因的丢失,同时还发现基因在3号染色体上发生了明显的片段复制,这可能是其扩张的潜在原因。

表1 野生二粒小麦PYL家族成员基因信息及其编码蛋白特征Table 1 Genomic information and characterization of protein encoded by PYL gene family members in wild emmer wheat
序列特征分析发现,野生二粒小麦基因的蛋白序列长度为123~253 aa,相对分子量为13.14~26.99 kDa,等电点为4.47~8.99,表明基因存在较大的组成变异。同时,亚细胞定位发现,有7个野生二粒小麦基因定位在叶绿体,1个定位在线粒体,其他16个均位于细胞核中。
2.2 野生二粒小麦PYL基因的系统进化、保守结构域和基因结构分析
为明确野生二粒小麦基因的系统进化关系,将鉴定到的24个野生二粒小麦基因与已报道的水稻和拟南芥基因进行系统进化分析。结果(图1)发现,所有基因聚为3大类,其中Class I中只包含有野生二粒小麦和水稻的基因,而Class II和Class III中包含有野生二粒小麦、水稻和拟南芥的基因,说明Class I的基因可能是单子叶植物特异的。根据系统进化树及水稻和拟南芥基因家族的分类信息,可将24个野生二粒小麦基因分为三大类,其中Class I亚家族包含9个成员,Class II亚家族包含11个成员,Class III亚家族只有4个成员(图2a)。

图1 野生二粒小麦、拟南芥和水稻PYL基因家族的系统进化分析
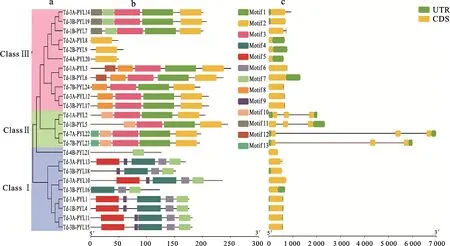
图2 野生二粒小麦PYL基因家族的系统进化关系(a)、保守基序(b)和基因结构(c)
利用MEME工具对野生二粒小麦基因保守结构域的组成和数目进行预测,结果共鉴定到13个保守的蛋白基序(图2b),其中,Class I亚家族均含有基序4、6和7,Class II亚家族均含有基序1、2、3和10,Class III亚家族除了、和只含有基序2外,其余基因均含有基序1、2和3。不同亚家族成员间具有各自特异的保守结构域,但同一亚家族成员具有相似的保守结构域组成,这可能与其具体的生物学功能相关,同一亚家族具有相似的生物学功能。
基因的结构是决定基因表达和功能的重要因素。对野生二粒小麦基因家族的外显子和内含子结构进行分析,结果(图2c)发现,野生二粒小麦家族的基因结构相对简单,含有的内含子很少,其中17个基因只含有1个外显子,没有内含子。从亚家族来看,每个亚家族成员间的基因结构基本一致,各个亚家族之间存在一定差异,其中Class II亚家族基因与其他亚家族基因相比,内含子和外显子数目较多,基因结构也较为复杂。
2.3 野生二粒小麦PYL基因启动子顺式作用元件分析
顺式作用元件在调节基因转录和表达方面发挥重要作用。对鉴定到的24个野生二粒小麦基因的启动子顺式作用元件进行预测,共鉴定到56个顺式作用元件,主要与ABA代谢、响应逆境和光调节相关(图3)。ABA响应元件ABRE在22个野生二粒小麦基因中被检测到,平均每个基因中含有3.4个ABRE元件,特别是、和分别有9、7和9个ABRE元件,表明其可能参与了野生二粒小麦ABA代谢调控过程。基因还含有丰富的光调节因子元件,如G-box和Sp1等。G-box在21个基因中被检测到,Sp1在15个基因中被检测到,暗示这些野生二粒小麦基因的表达可能受光调节。发现5个基因有MBS元件,表明这些基因可能还参与逆境胁迫的响应。

图3 野生二粒小麦PYL基因启动子的顺式作用元件
2.4 野生二粒小麦PYL基因在不同组织中的表达模式
为探究基因在野生二粒小麦生长发育过程中的潜在功能,利用RNA-Seq数据对野生二粒小麦基因在不同组织(根、叶、穗、颖片、外稃、花和种子)的表达模式进行研究。结果(图4)发现,野生二粒小麦基因在不同组织的表达模式有明显差异。、、和在出穗 1 cm时的表达量明显高于其他组织,而所有的基因在出穗3.5 cm时的表达量都比较低,表明这些基因可能在野生二粒小麦穗部的形态建成过程中发挥重要作用。在根中的表达量高于其他基因,而在叶中表达上调,、和在播种后112 d的颖片中表达量较高,在播种后112 d的外稃中表达量较高;、、、和在播种后105 d的花组织中表达量较高;、、和在播种后123 d的籽粒中表达较高;、和在播种后134 d的籽粒中表达量较高。

DS1和DS3.5分别代表出穗1 cm和3.5 cm;Flower_105、Flower_110、Flower_112分别代表105 d、110 d和112 d(均为播种后的天数,下同)的花组织;Glume_112代表112 d的颖片组织;Grain_123和Grain_134分别代表123 d和134 d的籽粒;Leaf_54和Leaf_57分别代表54 d和57 d的叶片组织;Lemma_112代表112 d的外稃组织;Root_20代表20 d的根组织。
2.5 野生二粒小麦PYL基因在干旱和盐胁迫下的表达模式
为挖掘与逆境胁迫相关的野生二粒小麦基因,以野生二粒小麦的两个品系TR9477和TTD22为材料,利用RNA-seq数据分析基因在干旱胁迫下的表达模式。结果(图5a)发现,在TR9477品系中,、和基因在干旱胁迫下的表达量明显上升;而在TTD22品系,这3个基因在干旱胁迫下表达量明显下降,暗示这3个基因可能与野生二粒小麦的抗旱性相关,参与了干旱胁迫的响应。

a:干旱胁迫;b:盐胁迫。图a中,TR39477和TTD22分别代表野生二粒小麦的两个品系;CK代表对照组;Drought代表干旱处理组;1~5代表生物学重复。
进一步对这些基因在野生二粒小麦耐盐品系A5盐胁迫下5个时间点(0 h、0.5 h、3 h、8 h和27 h)的表达模式进行分析。结果(图5b)表明,在0.5 h时,同源基因和明显上调表达;在3 h时,、、表达量均高于其他时间点,其中和是同源基因,表达模式相同;在8 h时,、、上调表达,其表达量高于其他基因;在27 h时,同源基因和的表达模式相同,均为上调表达。这些结果表明,多个基因参与了盐胁迫的响应,而且同源基因通常具有相似的表达模式,没有发生功能分化,这为进一步的生物学功能研究提供了候选基因。
2.6 野生二粒小麦及其他四倍体小麦PYL基因的单倍型
为了进一步探讨基因家族在四倍体小麦中的遗传分化,利用重测序数据分析野生二粒小麦、栽培二粒小麦和硬粒小麦基因的遗传变异和单倍型组成。结果发现,基因在这三个群体中具有明显的遗传差异,发生了显著的遗传分化。整体上,野生二粒小麦的遗传多样性较高,而硬粒小麦的遗传多样性较低。15个基因均有主要单倍型,而且其主要单倍型在野生二粒小麦、栽培二粒小麦和硬粒小麦群体中的分布具有明显的差异(表2)。其中野生二粒小麦中有6个基因的主要单倍型比例低于栽培二粒小麦,有14个基因的主要单倍型低于硬粒小麦,比如的主要单倍型TTCCGG,其在野生二粒小麦的比例为 42.86%,而在栽培二粒小麦和硬粒小麦的比例分别为 75.86%和84.62%;的主要单倍型为AAGGGGCCGGGG,其在野生二粒小麦的比例为35.71%,而在栽培二粒小麦和硬粒小麦的比例分别为55.17%和84.62%,这说明在野生二粒小麦驯化成栽培类型过程中,野生二粒小麦的基因选择了一些特定的单倍型作为主要单倍型被固定下来。此外,还发现的主要单倍型为CCCCAACCAA,其在野生二粒小麦和栽培二粒小麦的比例为78.57%和75.86%,而硬粒小麦的比例只有38.46%,这可能是在后期育种改良过程中,部分硬粒小麦中引入了其他来源的变异,降低了该基因的主要单倍型频率。

表2 PYL基因在四倍体小麦中的主要单倍型组成Table 2 Main haplotype of PYL genes in tetraploid wheat
3 讨 论
基因家族在许多植物中已被鉴定分离出来,本研究首次鉴定到野生二粒小麦中的基因,在全基因组范围中共搜索到24个基因,而在拟南芥和水稻基因组中基因家族的成员分别为13个和12个,均少于野生二粒小麦基因家族的成员数量。基因序列特征分析发现,基因的蛋白序列长度、分子量和等电点均存在较大的差异,亚细胞定位在叶绿体、线粒体和细胞核中,表明它们可能在不同的微环境中发挥着不同的功能。
野生二粒小麦基因聚为3个大类,其中Class I中只包含有野生二粒小麦和水稻基因,而Class II和Class III中包含有野生二粒小麦、水稻和拟南芥基因,说明Class I的基因可能是单子叶植物特有的。同源性较高的基因可能具有相似的功能,如拟南芥、和野生二粒小麦和具有较高的同源性,拟南芥、已被证明能促进ABA诱导的种子萌发,推测、也可能参与ABA诱导种子萌发的调控。
顺式作用元件通过响应不同外界环境信号来调节基因转录过程,进而影响植物的生长发育,本研究发现,野生二粒小麦基因启动子区域共有56个不同顺式作用元件,主要包括ABA响应元件ABRE和光响应元件Spl和G-box等,暗示基因在调节组织生长和发育过程中可能受到ABA代谢和光调节的影响,这与Kong等的研究结果一致,此外,本研究还检测到与MBS等逆境相关的顺式元件,表明野生二粒小麦基因的表达可由不同的胁迫诱导。
基因家族作为ABA的受体,参与多种ABA有关的生理反应,其中、、、、、、等已经被成功验证。在本研究中,大部分野生二粒小麦基因在根、叶、穗、颖片、外稃、花和籽粒等不同组织中广泛表达,表明它们可能参与多种生理反应。另外,野生二粒小麦TR39477品系中、和在干旱胁迫条件下上调表达,表明野生二粒小麦基因可能参与干旱胁迫响应。本研究还发现,在盐胁迫的各个时间点(0.5 h、3 h、8 h、27 h)都有不同的基因特异表达,表明野生二粒小麦基因可能参与盐胁迫的响应过程。总之,野生二粒小麦基因家族作为ABA受体,在野生二粒小麦抵抗逆境胁迫过程中发挥了作用,但各成员的具体功能需要后续试验进一步验证。对野生二粒小麦、栽培二粒小麦和硬粒小麦基因的遗传变异和单倍型组成的比较分析发现,基因在这三个群体中具有明显的遗传差异,发生了显著的遗传分化。
