家禽肠道健康关键营养技术
2022-02-26呙于明
■呙于明 刘 丹
(动物营养学国家重点实验室,中国农业大学动物科学技术学院,北京 100193)
维持家禽肠道健康对家禽发挥最大的生长性能具有重要作用。家禽养殖场每年因肠道健康问题遭受巨大的经济损失。在很长一段时间内,在饲料中添加抗生素是防止这一问题发生的主要预防策略。但是因细菌耐药和抗生素残留等公共卫生问题,欧盟已于2006年广泛禁止抗生素作为饲料添加剂在动物饲料中使用,我国农业农村部发布194 号公告宣告“自2020 年1 月1 日起,退出除中药外的所有促生长类药物饲料添加剂品种”。自此,我国饲料生产进入“后抗生素”时代。在安全高效的饲用抗生素替代品尚未出现的情况下,研究建立符合绿色生态畜牧业发展需求的综合营养技术体系来调控家禽肠道健康势在必行且完全可能。
1 家禽肠道疾病病因及其危害
多种病原体(细菌和寄生虫)被认为是引起家禽肠道疾病的主要原因。这些病原体既可以单独或与其他微生物一起导致肠道疾病,也可以通过与饲料或管理等非传染性因素协同作用。近年来,全世界主要养禽国家和地区都普遍存在由产气荚膜梭菌(Clostridium perfringens,Cp)过度增殖引起的坏死性肠炎(Necrotic enteritis,NE)。该肠道疾病在各日龄段的肉鸡均有发生,呈急性死亡,死亡率可高达50%。随着抗生素促生长剂的禁用,该病发生越来越严重,致使家禽生产性能和饲料报酬降低,鸡群死亡率陡增。而家禽生长早期的主要肠道疾病与沙门氏菌、大肠杆菌等的垂直传播感染和孵育管理不当有关(Hafez,2005)。此外,鸡空肠弯曲杆菌病时常会与其他疾病发生混合感染,如大肠杆菌病、沙门氏菌病、支原体病等,导致发病鸡群的死亡率明显升高。作为由寄生虫引起的鸡类疫病,鸡球虫病广泛存在于肉鸡、蛋鸡养殖过程当中,发生率和病死率较高。鸡患球虫病后,病鸡不仅会出现肠道上皮细胞坏死,而且肠黏膜有明显的出血症状,严重影响鸡只的生产性能和健康。霉菌毒素作为广泛存在于饲料原料中的真菌有毒次生代谢产物,可通过破坏肠上皮的形态、组织完整性和肠道菌群平衡来破坏肠黏膜的屏障功能,从而造成肠黏膜损伤,降低畜禽的生产性能。在畜禽饲料中常见且危害较大的霉菌毒素主要有黄曲霉毒素B1(Aflatoxin B1,AFB1)、玉米赤霉烯酮(Zearalenone,ZEA)、呕吐毒素(Deoxynivalenol,DON),而且三者共存的概率较高,具有叠加毒性,其危害也更大。此外,饲料成分如饲料原料中的不饱和脂肪酸和外源添加的脂肪氧化,生成自由基或活性氧代谢物,能使畜禽机体氧化与抗氧化系统失衡,消耗内源抗氧化剂,导致细胞凋亡和局部组织坏死,造成肠道组织氧化损伤(Dibner 等,1996;Haddad,2004)。诺伟司国际公司对2000~2005 年所收集的不同种类的脂肪进行了调查,发现40%~50%用于动物日粮的脂肪都不稳定,这一比例在炎热的夏季则会更高。
病原体相关分子模式(PAMPs)是存在于低等微生物或其细胞壁上的一类保守成分,广泛存在于病原体细胞表面, 如内毒素、肽聚糖、鞭毛蛋白、双链RNA以及酵母细胞壁上的甘露糖等。损伤相关分子模式(DAMPs)是从组织或细胞损伤中释放出来的内源性应激分子。PAMPs 和DAMPs 的主要结合受体是Toll样受体,不同的PAMPs 和DAMPs 可以被不同的Toll样受体所识别或组合识别,然后通过一系列蛋白质级联反应激活转录因子NF-κB 和Jun/Fos,从而导致编码促炎细胞因子、趋化因子和共刺激分子的基因激活,从而触发先天免疫反应(Piccinini 等,2010)。生长过程中的热应激、饲料中存在的霉菌毒素、氧化的油脂以及原料中的某些致敏蛋白也会引起肠道组织或细胞损伤进而释放DAMPs,通过被模式识别受体(PRR)识别后引起肠道炎症反应。同时,卫生条件差的鸡舍和污染的饲料中存在大量的病原菌,他们细胞表面的内毒素、肽聚糖、鞭毛蛋白等PAMPs 在进入肠道后会通过激活免疫细胞表面的PRR,进而引起Jak/Stat、TLRs/NF-κb 等信号通路激活,诱导IL-1β、IL-8等细胞因子的产生(Truong 等,2017)。大量的细胞因子同时又可以吸引巨噬细胞、中性粒细胞等聚集到感染部位,通过释放组胺、TNF-α、一氧化氮等消除有害物质,但当局部出现过多的白细胞聚集和过高浓度细胞因子就会引起紧密连接蛋白表达下调,肠上皮的完整性受损,严重时也可能激活全身炎症反应(Kogut 等,2018;Sato 等,2011)。免疫系统产生的IL-1β、IL-6、TNF-α等炎性细胞因子通过对靶组织的直接作用或通过作用于神经内分泌系统,从而改变机体激素水平如GH、IGF-1 和糖皮质激素(Häussinger 等,1992),影响机体蛋白质、脂肪和矿物质等营养物质的代谢(Andus 等,1991;Klasing 等,1991),造成生产性能如产蛋率、体增重、饲料转化率等降低(Quinteiro-Filho 等,2010;Quinteiro-Filho 等,2012;Zhang等,2020)。
2 肠道疾病防控与肠道营养技术
防控肠道疾病的关键在于消除肠道疾病病因或病原。供给清洁饲料和卫生饮水、抑杀或阻断病原微生物、坚固肠道屏障和增强免疫机能的综合多维的技术方案切实可行,可以保障“后抗生素”饲料时代养殖业提质增效。
2.1 清洁饲料
全面维护肠道健康当从清洁饲料开始。清洁饲料是指饲料内病原微生物、霉菌毒素、抗营养因子、有毒有害重金属、免疫应激原性物质都控制在零或接近于零的极低水平,因此不会释放PAMPs 和DAMPs 来激活肠道及全身炎症反应。
饲料加工企业里饲料原料采购的品质管控非常关键,好原料才能喂出好肠道。适当的加工技术也可提升饲料的清洁度。生物发酵饲料不仅可降低饲料中的抗营养因子和有害成分,且提高营养价值,主要表现在:改善消化道微生物菌群,防止腹泻量(Sugiharto 等,2020),提高养分消化吸收可利用率(Jazi 等,2019;Olukomaiya 等,2019),增强动物机体免疫抗病力(Lin等,2020),改善适口性、促进采食和生产(Gungor 等,2019),降解抗营养因子(Tang 等,2012),改善畜产品品质(Ahmed 等,2014),减少臭味氨味物质形成和释放、改善畜舍环境(Ahmed等,2014)等。
2.2 抑杀或阻断病原微生物
植物提取物、酸化剂、益生菌、酶制剂、寡糖等饲料添加剂均在一定程度上可直接抑杀肠道内微生物或阻断病原微生物在肠道黏膜上定植与增殖。
2.2.1 植物精油
植物精油(essential oils, EO)是植物的次级代谢产物,通过蒸馏法从植物中提取出来。常见的EO 来自牛至、百里香、迷迭香、丁香等芳香类植物,组成成分繁多,包括酚、醛、酮、醇、醚、烃类等。大量体外试验证实EO及其组成成分能够抑制微生物,包括细菌、真菌、病毒、原虫等,且对细菌的抑制作用具有广谱性,这些微生物绝大部分是动物或食源性的致病微生物(Brenes 等,2010;Chen 等,2021b;Lin 等,2016)。近几年一些植物EO 成分作为抗菌剂在家禽上的应用研究很丰富(见表1)。例如,肉仔鸡日粮中添加500 mg/kg 的牛至油或40 mg/kg 的牛至、月桂叶和薰衣草的混合精油还能降低粪便中的球虫卵囊排出量,减轻球虫感染所造成的肠黏膜损伤,其作用效果等同或优于60 mg/kg 的抗球虫药盐霉素(Achahbar 等,2012;Remmal 等,2011)。在感染坏死性肠炎的肉鸡上,牛至油可在科和属水平增加回肠乳杆菌丰度,降低编码微生物毒力因子ClpE(VF0073)、LPS(VF0124)、BSH(VF0350)和Capsule(VF0274)基因的相对丰度,下调TLR2和TNF-α基因的表达,从而降低坏死性肠炎肉鸡的肠道损伤评分和死亡率(Du 等,2016a)。可见,日粮中添加EO 可以缓解病原微生物的感染,改善感染家禽的肠道健康,因此能够作为抗生素的替代品应用于动物生产。
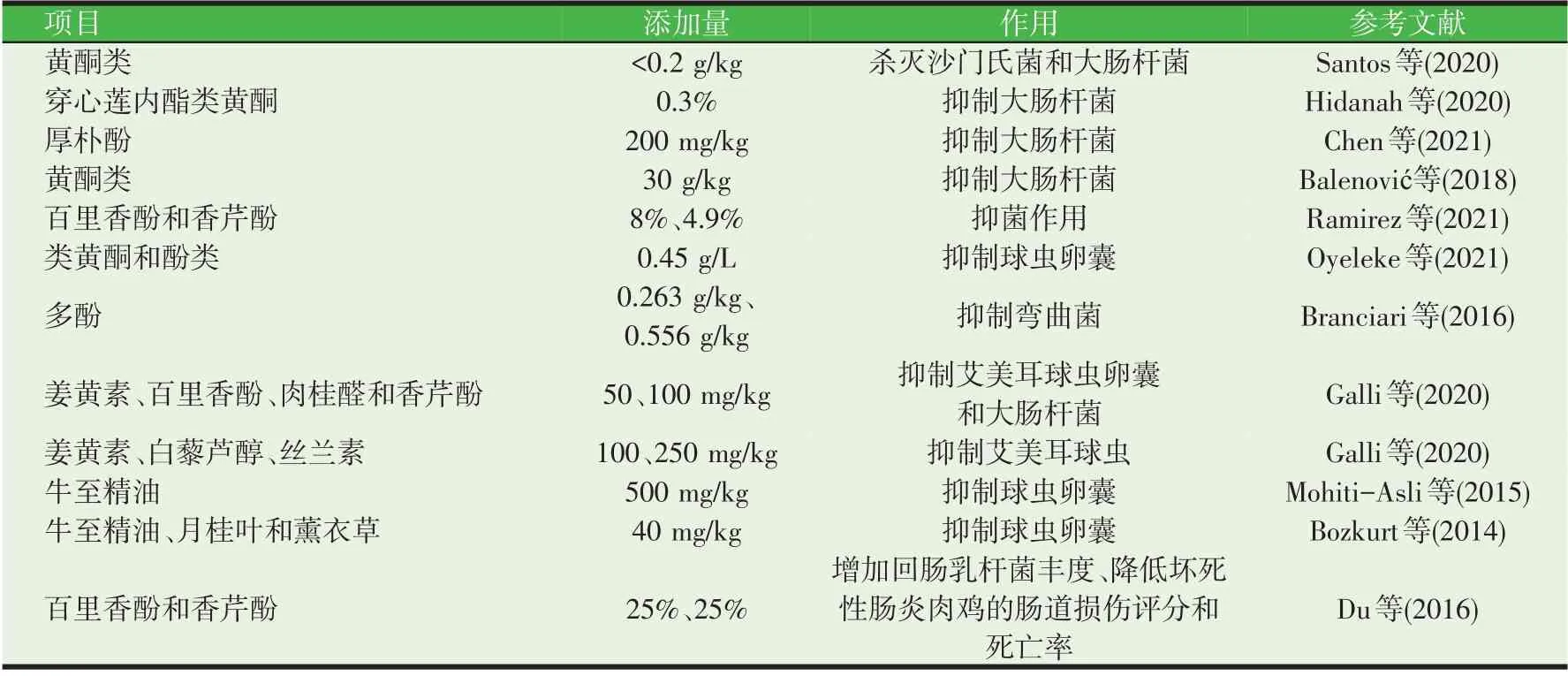
表1 植物精油成分作为抑菌剂在家禽上的应用研究
目前已有不少研究探索了EO 抑菌作用的机制。由于EO的化学成分非常复杂,其发挥抑菌作用的作用位点和作用机制不会局限于几种,很可能是对多个位点共同作用的结果。EO组成成分的抑菌活力大小为酚类>醛类>酮类>醇类>醚类>烃类。酚类物质含量高的EO,其抑菌能力更强。对于EO抗菌作用的机制可能与EO或其组成成分具有明显的亲脂性相关。当EO进入细胞膜时,可引起磷脂双分子层结构紊乱,破坏膜蛋白,造成细胞膜的渗透性增大,细胞内容物(包括代谢产物、离子等)漏出,质子动势缺失,ATP合成受阻,从而造成细胞死亡(Burt,2004;Langeveld等,2014)。
2.2.2 有机酸等酸化剂
酸化剂具有抑菌效果,可通过提高消化酶的活性有效降低饲料在消化道中的pH,从而改善畜禽肠道健康和营养物质的消化利用,促进动物生长。酸化剂可分为无机酸化剂和有机酸化剂,有机酸化剂因有良好的适口性,且可在体内循环发挥良好作用。在家禽上,近几年补充酸化剂对病原体感染后鸡生长有积极作用的研究报道见表2。例如,300 mg/kg单月桂酸甘油酯可影响大肠杆菌的丰度和艾美尔球虫卵囊的数量与增殖,并发挥了类似于抗生素的促生长作用(Fortuoso 等,2019)。日粮添加0.2%的有机酸混合物(包含11 种有机酸短链)在试验期前几天就能观察到明显的抗菌效果;相比于中链脂肪酸,有机酸更能降低肠球菌的丰度,这种菌一日龄禽类就会感染;有机酸在减少成年鸡的肠球菌丰度上同样比中链脂肪酸更有效;同时,有机酸混合物的有益菌乳酸菌和双歧杆菌属更丰富,其中卷曲乳杆菌和唾液乳杆菌数量最多,他们具有不同的益生作用(抗菌、生长促进剂和促健康)(Dauksiene等,2021)。
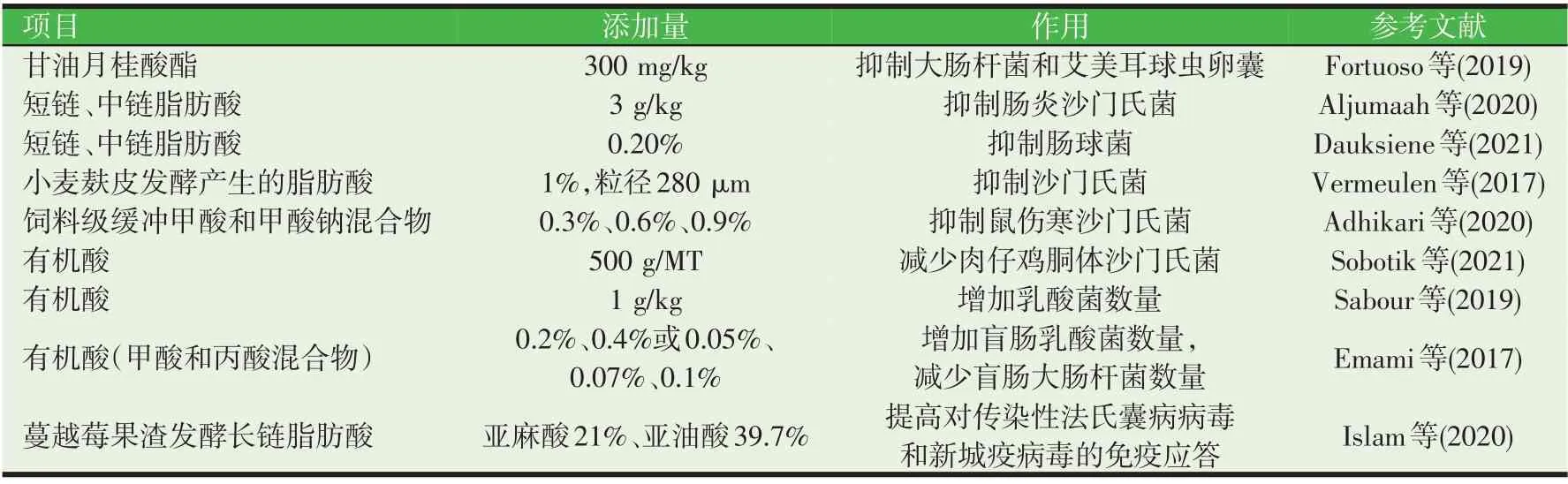
表2 有机酸在病原体感染家禽上的应用研究
2.2.3 酶制剂
非淀粉多糖酶不但可以通过消除非淀粉多糖(NSP)的“笼子”效应,释放更多可供动物利用的营养素,还可以减少可供病原菌利用的养分,生成更多可供肠道有益菌利用的益生元、阻断病原菌的寡糖,并降解免疫应激源(多糖),消除肠道组织炎症反应,从而来改善畜禽肠道健康。目前,饲用酶制剂种类丰富,且相当精细化和有针对性。不同的酶制剂因其底物种类和数量的差异及与其他酶的配伍,发挥着程度不同的维护肠道健康的作用。例如,木聚糖酶可以将麦类饲料中的主要抗营养因子木聚糖水解成木寡糖和木糖,不仅能成为可利用的物质为动物提供能量,还具有高效益生作用,促进肠道内有益菌的增殖,抑制肠道内腐败菌的生长并减少有毒发酵产物的形成(Lei等,2016)。在病原菌感染的情况下,木聚糖酶可改善坏死性肠炎肉鸡肠道的屏障功能,减少产气荚膜梭菌在肠道内的定植,降低死淘率(Liu等,2012)。葡萄糖氧化酶(Glucose oxidase,GOD)能消耗肠道中大量的氧气并特异地将β-D-葡萄糖氧化为葡萄糖酸和过氧化氢,从而使肠道内形成厌氧酸性环境而有利于厌氧耐酸菌的增殖。肉鸡日粮中添加GOD可降低回肠pH,提高了绒毛高度和绒毛高度/隐窝深度,改善坏死性肠炎肉鸡的肠道屏障的完整性和回肠菌群的平衡(Zhao等,2021)。
2.2.4 益生菌
大量的体外体内试验已证实益生菌能够通过多种机制对肠道屏障进行调节以抑制病原微生物在肠道内定植和生长,有的还可以通过分泌抗菌物质杀灭有害微生物(刘欢欢等,2020;De Keersmaecker 等,2006;El-Sharkawy 等,2020;Fernandez 等,2019;Lone等,2021;Pridmore 等,2008),因此对病原体引起的肠道疾病能起到良好的预防和治疗作用。芽孢杆菌、乳酸菌和丁酸梭菌是三类广泛应用于禽类的益生菌。例如,有研究报道芽孢杆菌制剂可降低雏鸡沙门氏菌的发病率和肠道肠炎链球菌数量(Adhikari等,2019);添加枯草芽孢杆菌B21 或地衣芽孢杆菌B26,可改善肠道形态,增加短链脂肪酸浓度,部分减轻肉鸡坏死性肠炎亚临床症状(Musa 等,2019);补充枯草芽孢杆菌可提高盲肠中与碳水化合物代谢和丁酸、丁酸盐产生有关的黄杆菌属、劳特氏菌属、粪杆菌属的相对丰度(张雪等,2020);丁酸梭菌添加可增加ETEC 攻毒条件下肉鸡回肠的抗菌肽AvBD10 的基因表达量(申露露等,2020)。乳杆菌KL1 和植物乳杆菌Zhang-LL混合可降低雏鸡白痢的死亡率,调节肠道菌群平衡(Chen等,2020)。
2.2.5 益生元
益生元可由特定的肠道微生物菌群代谢为宿主提供能量,并通过调节肠道微生物菌群防止病原体黏附宿主细胞来影响肠道健康。肠道微生物菌群通过发酵益生元生成短链脂肪酸(主要是乙酸、丙酸和丁酸),降低肠腔pH,可为上皮细胞提供能源,也可促进有益菌的生长,抑制有害菌的增殖,并对炎症反应和代谢调节有影响。有些低聚糖和单糖可减少病原体在上皮细胞表面位点的定植(杜晨红等,2019;Townsend,2019)。在家禽上,酵母培养过程中产生的功能性碳水化合物可促进肉鸡生长,减少病原菌定植(Froebel等,2019);菊粉能影响肉鸡盲肠菌群组成、动态及生长相关参数(Xia等,2019);甘露寡糖和β-葡聚糖合用可缓解肉鸡感染大肠杆菌造成的发育迟缓和异常情况,减少大肠杆菌感染和定植(Fadl 等,2020)。小球藻可改善肉鸡肠道微生物群和肠道形态(Alfaia等,2020)。
2.3 坚固肠道屏障
受环境应激等因素的影响,生长早期的家禽肠道屏障很容易受损伤以及免疫机能发育不良,肠道菌群失衡紊乱进而引发各种疾病和生产性能降低。肠道屏障是由物理屏障、化学屏障、微生物屏障和免疫屏障组成的,其中物理屏障尤为重要。物理屏障是由肠黏膜上皮细胞及其紧密连接等组成的完整的彼此紧密连接的肠道上皮结构。紧密连接的作用为封闭细胞间的间隙,阻止肠腔有毒物质进入周围组织,其通透性决定着整个肠上皮细胞的屏障功能(Turner,2006)。紧密连接由不同的跨膜蛋白包括Occludin、Claudin 和JAMs 组成,这些跨膜蛋白通过脚手架蛋白如ZO蛋白与细胞骨架相连。通过营养手段可促进有利于家禽肠道屏障的紧密连接蛋白的表达,改善物理屏障的完整性,维护肠道健康。营养手段包括在家禽日粮添加微量元素(Sun 等,2020)、维生素(VD3等)(Wang,2021)、功能性氨基酸[精氨酸、谷氨酰胺、色氨酸等(Barekatain等,2019;Wu等,2018a;Yue等,2017;Zhang等,2017)]等营养性添加剂以及合理使用植物提取物(He等,2020;Liu等,2020)、多聚糖(β-1,3/1,6-葡聚糖、车前草非淀粉多糖)(Parsons 等,2014;Shao 等,2013)、益生菌(Wu 等,2018b)、酸化剂(Ma 等,2021)等非营养性添加剂。例如β-1,3/1,6-葡聚糖可上调鼠伤寒沙门氏菌肉仔鸡Claudin-1 和Occludin 的mRNA表达,降低盲肠沙门氏菌数量和肝脏中沙门氏菌的数量(Shao 等,2013)。添加车前草非淀粉多糖50 mg/d 减少沙门氏菌对肉仔鸡脾脏的侵袭,经回肠黏膜尤斯灌流室(Ussing chamber)离体试验证实这种作用可能与增加的黏膜短路电流而跨上皮电阻不变相关(Parsons等,2014)。
2.4 增强免疫机能
通过增强机体的免疫机能可有效防控家禽肠道疾病,但需要从呼吸道、胃肠道、生殖道等的黏膜免疫和全身性免疫两个层面来着手,利用疫苗和抗体等针对性地激活特异性免疫和利用营养技术措施适度调控机体非特异免疫功能。可以根据幼龄畜禽、高温高湿高密度集约化养殖等环境应激、引起免疫抑制的病毒疫苗、病原体的变异和疫病流行地域等特殊状况可通过多种营养措施来有针对性地强健免疫系统。目前,广泛应用于增强家禽免疫机能的营养措施同样包括添加微量元素(El-Katcha 等,2017;Habibian 等,2015)、维生素(Lin 等,2002)、功能性氨基酸(Jian 等,2021;Murakami 等,2014)、脂肪酸(Guo等,2004)等营养素,和合理使用活性寡糖(Jahanian 等,2019)、多糖(β-1,3/1,6-葡聚糖)(Shao 等,2013)、益生菌(Fathi等,2018)、黄酮类(Fan 等,2018b)等非营养性添加剂。例如,添加甘露寡糖可提高肉鸡抗传染性支气管炎、法氏囊和新城疫病毒的抗体含量,提高淋巴细胞比例和肝脏IL-2 基因表达(Jahanian 等,2019)。β-1,3/1,6-葡聚糖可增加感染鼠伤寒沙门氏菌肉仔鸡肠道sIgA阳性细胞数和sIgA含量,降低盲肠沙门氏菌数量和肝脏中沙门氏菌的数量,减缓由感染鼠伤寒沙门氏菌引起的肉仔鸡肠道屏障功能的损失(Shao等,2013)。肉鸡饲粮中添加500 mg/kg山花黄芩提取物(绿原酸2.2 mg/g、黄芩苷22 mg/g)可通过提高肉仔鸡的法氏囊指数和抗炎因子TGF-β的表达量,降低促炎因子IL-6 等的表达量来缓解LPS 引起的免疫炎症反应。
2.5 优化早期营养
初生雏鸡消化道系统、免疫系统、肠道菌群、骨骼发育和肌肉生长均尚未发育成熟和稳定。14 日龄之前是肉鸡消化道系统和免疫系统发育的黄金时期(栾维民,2008;Lammers 等,2010),也是肠道内消化酶合成量最大的时期(Noy 等,2010)。此时,外源性早期饲料营养则是促进雏鸡各系统生长发育的营养来源,供给不足则会影响各系统的发育和功能的完善。早期的日粮能量和蛋白质(Ullah 等,2012;Wijtten 等,2004;郑爱武等,2014)等营养水平和营养素组成(Kidd,2004),如氨基酸(Konashi 等,2000)、微量元素(万妍等,2020)和非营养素如益生菌(Meijerink 等,2020)都能直接或间接地影响家禽各系统的早期发育。万妍等(2020)认为,在肉仔鸡早期基础日粮中额外添加120 mg/kg 硫酸锌有利于肉鸡生长性能的提高,促进免疫力的提升。
动物发育早期的基因表达容易受到表观遗传调控的影响(Sandovici等,2011),种鸡的激素、免疫分子等活性物质或其代谢物,以及日粮中的营养成分能够通过种蛋沉积到子代,在子代发育早期通过表观遗传修饰引起代谢程序化,调控基因表达,留下“代谢印记”,对子代的新陈代谢产生长期的、终身的影响,这种影响称为种鸡营养的印记效应,或营养表观遗传效应。合理配置种鸡的营养可能对于子代的早期营养及后续的健康和生长是至关重要的。新孵化鸟类的适应性免疫系统功能不成熟,依赖来自蛋黄转移的母体抗体维持机体免疫(Al-Natour等,2004;Garnier等,2012)。在肉种鸡日粮补充锌尤其是有机锌能够提高子代的肠道屏障功能(李昌武,2015),其机制包括有机锌能够通过表观遗传修饰提高子代肉鸡A20 基因表达,抑制NF-κB信号通路介导的炎症反应,上调肠上皮细胞紧密连接蛋白的表达,提高肠上皮完整性,降低通透性(Li 等,2015)。在肉种鸡日粮中添加20 mg/kg 大豆黄酮(Daidzein,DA)通过活化一种新型的lncRNA-XLOC_098131来间接调控FOS基因表达,调控Toll样受体等免疫相关通路,提高淋巴细胞活力以及免疫球蛋白含量等,增强产蛋后期肉种鸡免疫抗病能力;并能够提高子代胚胎期的发育和肉仔鸡生长期的肌肉发育与免疫能力,这种调控效应可能是通过影响DNA 甲基化、组蛋白修饰以及lncRNA 调控来实现的(Fan等,2018a;Fan等,2019)。
3 未来展望
大量研究表明肠道微生态是决定肠道健康的关键。宿主及其相关微生物群落一起,形成了一种协同关系,称为全生物体。将动物营养的研究深化到全生物体精准营养,或基于微生物组学的肠道微生物精准营养,会成为肠道健康营养技术的突破点。例如,视黄醇或视黄酸可能通过调节特定的微生物群如乳酸菌和上调IFN-β介导的抗病毒免疫应答来实现抑制小鼠诺如病毒的复制(Lee 等,2016)。肠道共生细菌中多形拟杆菌的生长可能通过消耗维生素B12来影响肠道中维生素B12的浓度,从而抑制食源性病原体肠出血性大肠杆菌毒素STX2 的合成,从而调节肠道健康(Cordonnier 等,2016)。全生物精准营养研究需要包括以下几方面:准确的饲料养分信息即含量与效价;精细的饲料加工工艺即设备与参数;精准的动物营养需求和精准的肠道微生物营养需求等等。
