黄羽肉鸡日粮中添加胆汁酸盐对肠道黏膜转运蛋白及消化酶的影响
2022-02-26张振明贺海莲王静鸽李鹏祥刘雄杰郭家卉丁保安
■张振明 贺海莲 王静鸽 李鹏祥 刘雄杰 郭家卉 丁保安
(青海大学农牧学院,青海 西宁 810016)
胆汁酸(BAs)是由肝脏中独有的胆固醇合成的,是胆汁中特异性和定量的重要有机成分[1],有助于促进脂质和脂溶性维生素的吸收和转运。禽类胆汁酸主要由去氧胆酸和胆酸组成[2]。近年来,胆汁酸在畜牧业中的积极作用已有报道,在日粮中添加BAs可以提高肉鸡的生产性能,有效提高饲料养分利用率,胴体品质得到提高[3-4]。胆汁酸作为一种新型的饲料添加剂,被广泛应用于动物饲料中,以促进脂肪的吸收,是哺乳动物胆固醇分解代谢的主要途径。但是禽类有关胆汁酸对脂质代谢的研究报道很少。除了作为促进脂质吸收的乳化剂,还发现胆汁酸作为信号分子在机体的多种代谢过程中发挥着重要的调节作用,而且在糖脂代谢中也同样发挥着不可或缺的作用[5-6]。固醇调控元件结合蛋白1(SREBF1)属于核转录因子家族,是动物体内脂肪合成的一个极其重要的调节因子,在脂肪酸合成过程中发挥着重要的作用,从而控制体内脂肪合成[7]。脂肪酸转运蛋白4(FATP4)存在于肠绒毛细胞中,被认为是调控肠道细胞吸收脂肪酸的重要因子[8]。乙酰辅酶A 羧化酶(ACC)是脂肪酸合成的主要限速酶,对脂肪酸的代谢调控有着重要的生物学作用[9]。脂肪酸合成酶(FAS)对动物体脂肪的沉积性状起到了重要的调节作用,是动物体脂肪酸合成的关键酶之一[10]。本试验以黄羽肉鸡为研究对象,在黄羽肉仔鸡日粮中添加不同水平胆汁酸盐,探究其对肠道黏膜中营养转运蛋白和消化酶的基因表达的影响,以期为青海农业区黄羽肉鸡的养殖提供理论依据。
1 材料与方法
1.1 试验动物与试验设计
选择同批次孵化、体格健康良好的420 只1 日龄黄羽肉雏鸡(红羽王)为试验动物。采用单因素试验设计,将420只雏鸡分为7个组(每组4个重复),每个重复15只鸡,试验1组为阴性对照组,饲喂基础日粮,试验2 组为阳性对照组,在基础日粮上添加0.02%泰乐菌素代替麸皮,试验3组、试验4组、试验5组、试验6 组、试验7 组分别添加0.015%、0.025%、0.035%、0.045%、0.055%的胆汁酸代替麸皮。进行为期28 d的饲养试验。
1.2 试验日粮与营养水平
试验日粮组成与营养水平见表1。胆汁酸盐为混合型饲料添加剂,购自山东某有限公司,有效成分为胆汁酸,含量≥30%。泰乐菌素购自某生物药业有限公司,纯度为10%。
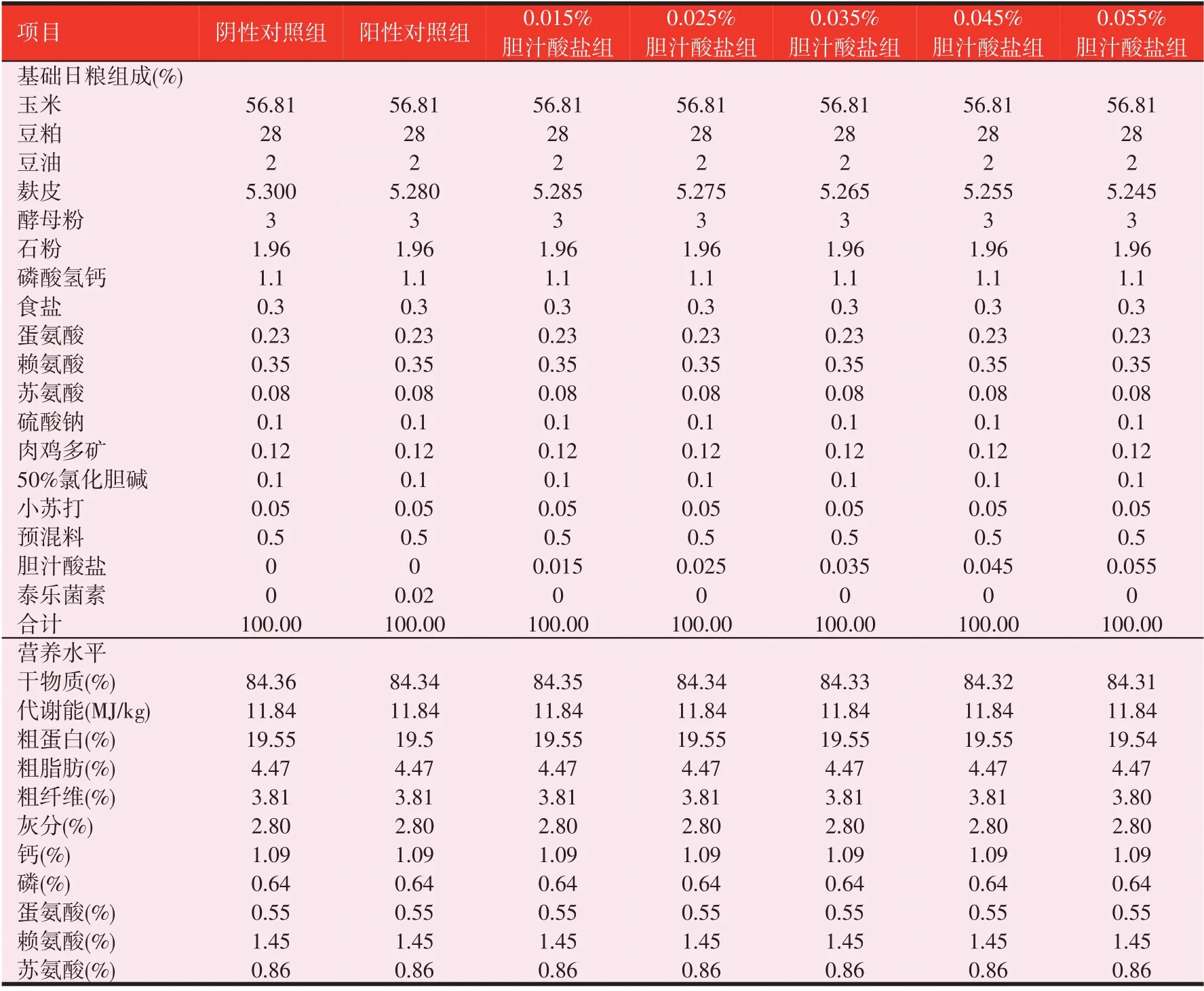
表1 基础日粮组成和营养水平(风干基础)
1.3 饲养管理
饲养期间为人工喂料,自由采食和饮水,第1 周每天24 h 光照,之后以0.5 h/d 递减直至第26 d,随后每天保持17 h 光照。第1~7 d 室温控制在33~35 ℃,第8~14 d室温控制在29.5~32.5 ℃,第15~28 d室温控制在24~29 ℃。湿度为60%~70%,按照常规免疫程序进行定期消毒,打扫鸡舍卫生。饲养试验在青海省湟中县常绿牧业公司养鸡场进行。
1.4 样品采集
饲养至第28 d每个重复随机选取1只鸡,颈部放血致死,宰前12 h不采食。采集十二指肠、空肠、回肠和肝脏。样品都置于2.0 mL effendoff管,然后液氮速冻,6 h内放于-80 ℃冰箱长期冷冻保存。
1.5 荧光定量PCR
1.5.1 样品总RNA提取
采用Trizol 一步法提取总RNA,利用核酸蛋白仪器检测RNA的浓度和纯度,检测是否合格,进行后续试验。
1.5.2 反转录
反转录的体系为20 μL(见表2),整个操作过程在冰盒上完成,样品与试剂混匀后于GenAmp PCR System 9700 仪器37 ℃反应30 min,85 ℃变性15 s,4 ℃保存。反转录产物置于-20 ℃保存使用。

表2 反转录体系
1.5.3 目的基因引物设计及qPCR反应体系
PCR引物来自参考文献[11-12],由生工生物工程(上海)股份有限公司合成,引物序列见表3。

表3 目的基因SREBF1、FATP4、FAS、ACC和内参基因β-actin引物序列
采用SYBR Green 实时荧光定量PCR 方法,以βactin作为内参基因,对SREBF1、FATP4、FAS、ACC 进行相对定量分析。qRT-PCR 反应在BIO-RAD CFX96 Touch上进行。
PCR 反应体系:采用单管法进行PCR 扩增,以混合样(待测样品等比例混合)对PCR反应条件,循环数以及目的基因SREBF1、FATP4、FAS、ACC和内参基因β-actin的引物浓度等进行优化。各目的基因qPCR条件如下:在20 μL的反应体系中含有2 μL反转录产物,10 μL TB Green Premix Ex Taq,6.8 μL ddH2O,上、下游引物各0.6 μL。PCR 反应条件为:95 ℃预变性2 min;95 ℃变性15 s,58 ℃退火30 s,40 个循环。每个待测样品设置3个平行,取3个Ct值的平均值作为后续的公式计算。
1.6 数据统计
数据采用Excel 2019 进行整理,采用2-ΔΔCt方法(Livak 法)计算目的基因的相对表达量,用SPSS 22.0 General Liner Model 的Univariate 方法进行双因素方差分析,对F检验达到显著水平的因子进行LSD法多重比较,分析结果包括胆汁酸盐水平、肠段以及“胆汁酸盐水平×肠段”互作三个影响模型。不同肠段的SREBF1、FATP4、FAS、ACC mRNA表达的发育变化用ANOVA 进行单因子方差分析,用Duncan’s 法进行多重比较检验,以P<0.05 作为显著性差异判断标准,结果均以“平均值±标准差”表示。
2 结果与分析
2.1 胆汁酸盐对黄羽肉鸡SREBF1 mRNA 表达量的影响(见表4)
由表4 可知,随着胆汁酸盐水平的递增,十二指肠和回肠SREBF1 mRNA 相对表达量呈先增后减再增的趋势。在十二指肠中,0.055%胆汁酸盐组SREBF1 mRNA 相对表达量最高,且显著高于其余各组(P<0.05);在空肠中,0.055%胆汁酸盐组表达量显著高于其余各组(P<0.05);在回肠中,0.055%胆汁酸盐组表达量显著大于其余各组(P<0.05)。肝脏中,0.035%和0.055%胆汁酸盐组表达量显著大于其余各组(P<0.05)。

表4 黄羽肉鸡28日龄雏鸡SREBF1 mRNA的相对表达量
2.2 胆汁酸盐对黄羽肉鸡FATP4 mRNA表达量的影响(见表5)
由表5 可知,随着胆汁酸盐水平的递增,回肠和肝脏FATP4 mRNA 相对表达量呈先减后增的趋势。在十二指肠中,0.055%胆汁酸盐组和阳性对照组FATP4 mRNA相对表达量较高,且显著高于其余各组(P<0.05);在空肠中,0.015%、0.045%胆汁酸盐组和阴性对照组的相对表达量高于其余各组(P<0.05);在回肠中,0.055%胆汁酸盐组的相对表达量显著大于其余各组(P<0.05);在肝脏中,0.045%和0.055%胆汁酸组的相对表达量显著大于其余各组(P<0.05)。
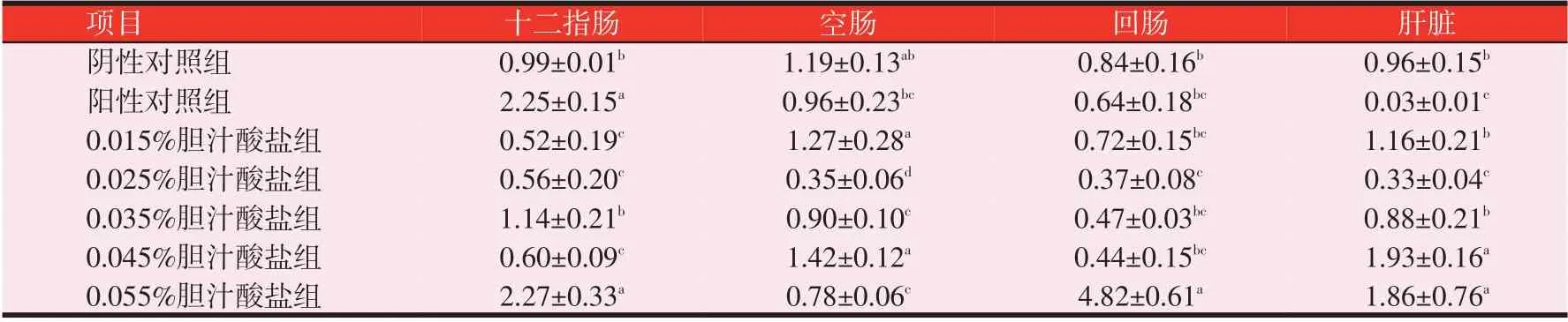
表5 黄羽肉鸡28日龄雏鸡FATP4 mRNA的相对表达量
2.3 胆汁酸盐对黄羽肉鸡FAS mRNA 表达量的影响(见表6)
由表6 可知,随着胆汁酸盐水平的递增,回肠FAS mRNA相对表达量呈逐渐增加的趋势。在十二指肠中,0.035%胆汁酸盐组FASmRNA相对表达量较高,且显著高于其余各组(P<0.05);在空肠中,0.015%胆汁酸盐组相对表达量显著高于其余各组(P<0.05);在回肠中,0.055%胆汁酸盐组相对表达量显著高于其余各组(P<0.05);在肝脏中,0.015%、0.035%和0.055%胆汁酸盐组的相对表达量显著高于其余各组(P<0.05)。
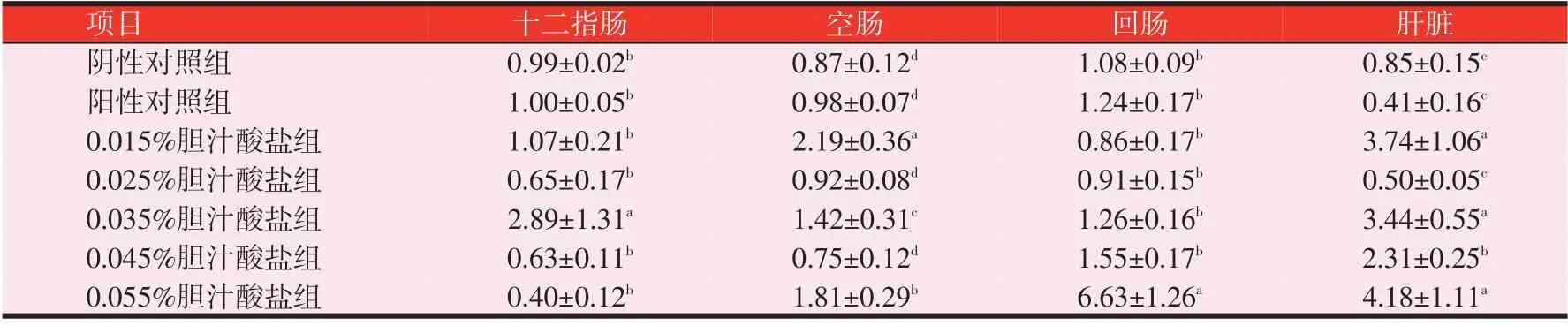
表6 黄羽肉鸡28日龄雏鸡FAS mRNA的相对表达量
2.4 胆汁酸盐对黄羽肉鸡ACC mRNA表达量的影响(见表7)
由表7 可知,随着胆汁酸盐水平的递增,十二指肠和空肠ACC mRNA 相对表达量呈先增后减再增的趋势。在十二指肠中,0.055%胆汁酸盐组ACC mRNA相对表达量较高,且显著高于其余各组(P<0.05);在空肠中,0.055%胆汁酸盐组相对表达量显著高于其余各组(P<0.05);在回肠中,0.055%胆汁酸盐组相对表达量显著高于其余各组(P<0.05);在肝脏中,0.015%胆汁酸盐组相对表达量显著高于其余各组(P<0.05)。
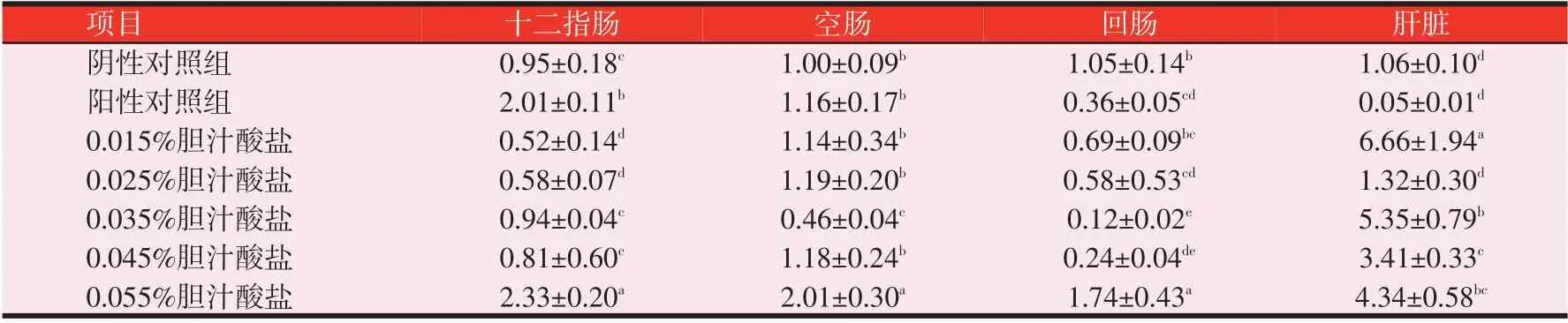
表7 黄羽肉鸡28日龄雏鸡ACC mRNA的相对表达量
2.5 肠段与不同水平胆汁酸盐对SREBF1、FATP4、FAS、ACC mRNA相对表达量的影响(见表8)
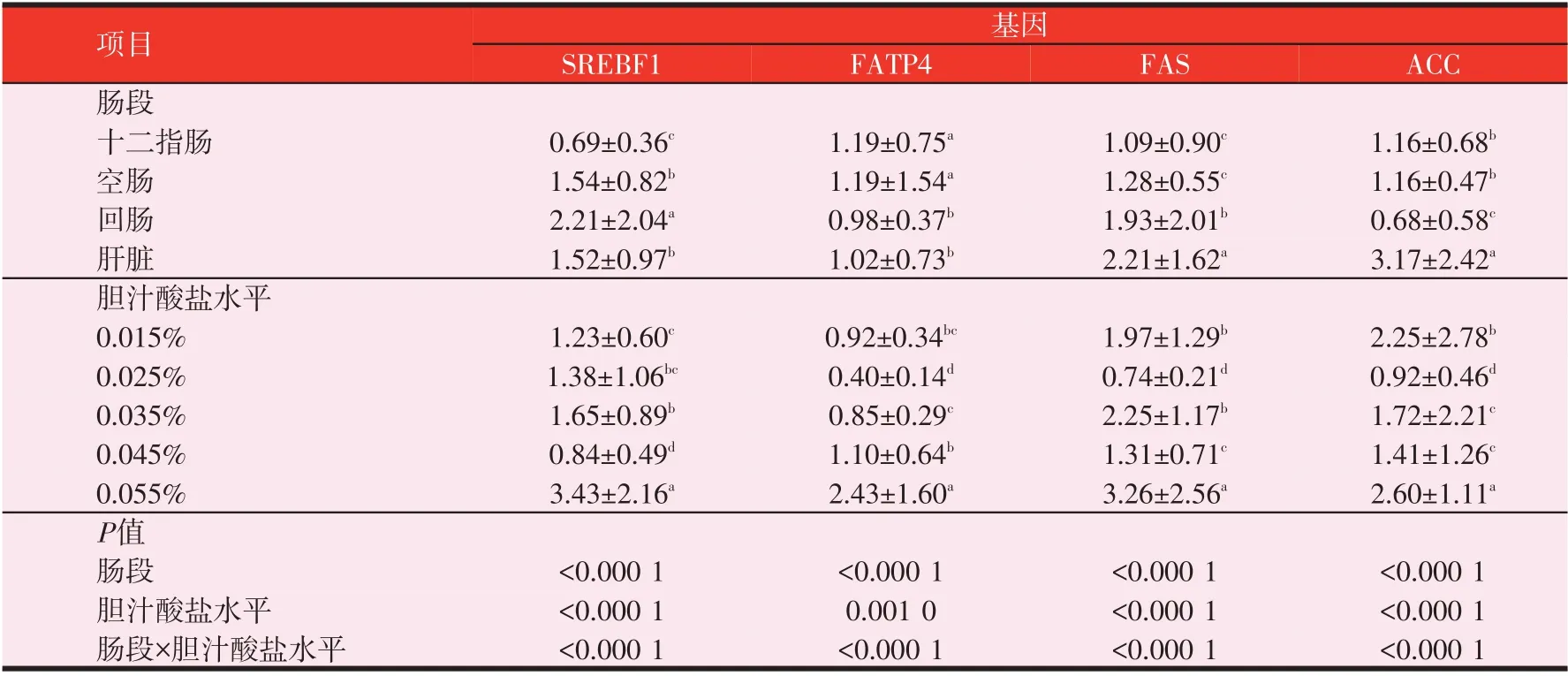
表8 肠段、胆汁酸盐水平对SREBF1、FATP4、FAS、ACC mRNA的相对表达量的影响
由表8可知,SREBF1 mRNA的相对表达量回肠较高,其次是空肠和肝脏,十二指肠最低;SREBF1 mRNA的相对表达量回肠显著高于十二指肠、空肠和肝脏(P<0.05),空肠和肝脏显著高于十二指肠(P<0.05)。FATP4 mRNA 的相对表达量十二指肠、空肠较高,其次是肝脏和回肠;十二指肠和空肠显著高于回肠和肝脏(P<0.05),回肠和肝脏之间差异不显著(P>0.05)。FAS mRNA 的相对表达量肝脏较高,其次是回肠,之后是空肠,十二指肠最低;肝脏、回肠都显著高于十二指肠、空肠(P<0.05),十二指肠与空肠之间差异不显著(P>0.05)。ACC mRNA的相对表达量肝脏较高,十二指肠、空肠次之,回肠最低;肝脏显著高于十二指肠、空肠和回肠(P<0.05),但十二指肠和空肠之间差异不显著(P>0.05)。
黄羽肉鸡十二指肠、空肠、回肠和肝脏SREBF1、FATP4、FAS、ACC mRNA的相对表达量均存在极显著差异(P<0.01);不同胆汁酸盐添加水平黄羽肉鸡肠道SREBF1、FATP4、FAS、ACC mRNA相对表达量存在极显著差异(P<0.01);SREBF1、FATP4、FAS、ACC mRNA的相对表达量均存在“肠段×胆汁酸盐水平”的互作效应(P<0.01)。
3 讨论
3.1 雏鸡肠道SREBF1 mRNA 的表达差异与不同水平胆汁酸盐下的变化
动物采食后,胆汁酸进入小肠,进行乳化脂肪。乳化后的脂肪被水解为甘油和脂肪酸,胆汁酸和脂肪酸形成的复合物被小肠黏膜所吸收,之后与脂肪酸分离,经门静脉进入肝脏。之后游离型胆汁酸再转变为结合型胆汁酸,与胆汁一起重新进入肠道,这一过程称为胆汁酸的肝肠循环[13]。固醇调控元件结合蛋白(SREBP)是一种属于基本螺旋-环-螺旋亮氨酸链家族的转录因子,编码SREBP-1a 和SREBP-1c,两者都可以调节与脂质代谢相关的基因表达,尤其是脂肪生成基因[14]。有报道说,一旦SREBP-1a被激活,SREBP-1b会增强胆固醇和脂肪酸合成相关基因的表达,从而达到预防脂肪肝的发生[15-16]。Yin等[17]研究了在日粮中添加胆汁酸补充剂来改善热应激肉鸡的生长性能和脂质代谢,得出了日粮中添加胆汁酸降低了三酰甘油和肝脏SREBP-1c mRNA的表达结论。Kamisako等[18]研究在小鼠饲料中添加胆固醇、胆汁酸盐或消胆胺对与胆固醇和胆汁酸代谢相关的肠道mRNA 调节的影响中得出胆固醇组和胆汁酸盐组肠道SREBP-1c mRNA表达显著增加。消胆胺组肠道SREBP-1c mRNA 表达明显降低。而本试验中得出胆汁酸盐组肠道SREBF1 mRNA 表达显著增加。这与Kamisako 等的结果有相似之处。Ge 等[11]研究发现在肉鸡日粮中补充胆汁酸可有效改善SREBP-1 mRNA 的表达,从而降低脂肪酸的合成。陆江等[19]研究发现在产蛋后期日粮中添加胆汁酸可以调节SREBP1c mRNA 的表达,抑制脂肪酸的从头合成,促进脂肪酸β-氧化,从而降低蛋鸡血液和肝中的TG,以0.06 g/kg 添加剂量为宜。SREBF1 mRNA的相对表达量随着胆汁酸盐水平的递增,呈现一种较为规律的变化,且最适添加水平范围为0.055%。根据这一结果可以在分子水平方面推测出,对家禽胆汁酸盐添加水平的调节,满足其对营养物质的调控具有一定价值。
3.2 雏鸡肠道FATP4 mRNA的表达差异与不同水平胆汁酸盐下的变化
脂肪酸是动物体生命活动所必需的能量物质,在动物体内的吸收、转运和代谢是一个比较复杂的过程。需要通过肠道上皮细胞来完成[20]。FATPs对长链脂肪酸的吸收和代谢发挥着重要的作用,FATP4是动物肠道中唯一存在的FATPs家族成员,存在于肠绒毛细胞中,是调控肠道细胞吸收脂肪酸的重要调控因子[21-22]。王彦[23]研究了FATP4基因在鸡上的表达模式并进行了单核苷酸多态性(SNP)分析,发现FATP4基因在不同鸡的品种间表达差异不显著,FATP4基因型与鸡的腹脂率、腿肌率、净膛重等屠宰性状具有显著的正相关。罗涓[24]试验发现FATP4 基因在不同组织中的表达丰度从高到低依次为:腿肌、肝脏、皮脂、肠道、腹脂。FATP4在腿肌中的表达量极显著高于其他各组织。FATP4 基因在各个组织中的表达量基本上呈现出肌肉、胸肌和腿肌中最多,在脂肪组织腹脂和皮脂中最少的规律,在肌肉组织和心脏中的基因表达量差异不显著。肝、肠、腹脂、皮脂中的基因表达量差异不显著。不同品种间的表达差异不显著。
本试验结果表明FATP4 mRNA 表达量在空肠和十二指肠中最高,其次是回肠和肝脏。这与张爱华[12]研究结果一致,表明雏鸡肠道吸收转运脂肪酸的主要部位是在小肠前端。在不同水平胆汁酸盐水平递增的情况下,FATP4 mRNA的相对表达量在0.055%水平最高,且显著高于0.015%、0.025%、0.035%、0.045%水平。在本试验中发现添加不同水平胆汁酸盐可以调剂小肠和肝脏中FATP4 mRNA的表达量,这可能与胆汁酸盐的作用有关。
3.3 雏鸡肠道FAS和ACC mRNA的表达差异与不同水平胆汁酸盐下的变化
FAS和ACC是脂肪酸合成途径中的关键限速酶,在脂肪合成代谢过程中发挥着不可或缺的作用,其活性和表达可以调节脂肪酸的合成,从而影响脂肪代谢及体脂沉积[25]。其中乙酰辅酶A羧化酶(ACC)是催化脂肪酸合成的第一步,脂肪酸合成酶(FAS)是参与合成三酰甘油的关键酶[26]。日粮中的营养成分可以影响编码这些酶的基因的表达,从而调节脂肪沉积。陆江等[19]研究发现,在产蛋后期日粮中添加合适的胆汁酸,可以调控FAS mRNA 的表达,从而降低脂肪酸的合成,减少脂肪沉积。田卫华等[25]研究表明在产蛋期母鸡饲粮中添加8%水平的大豆油能够明显提高FAS、ACC mRNA 表达水平,在肝脏组织中表达量最高。这与本试验结果有相似之处,FAS mRNA的表达量在肝脏中最高,回肠次之,十二指肠最低和空肠。Ge 等[11]研究发现在肉鸡日粮中补充胆汁酸可以改善FAS和ACC mRNA的表达,从而起到促进脂肪分解代谢和抑制脂肪合成的作用。Piekarski等[3]研究发现在日粮中添加0.5%纯化的胆汁酸可以有效调节肉鸡组织中ACC-1c 和FAS mRNA 的表达水平,从而抑制了采食量,并且降低肉鸡体重。葛晓可等[27]发现在肉鸡日粮中添加胆汁酸可以使FAS mRNA 的表达量得到调控,得出胆汁酸可以通过抑制脂肪酸的合成及转运,并促进脂肪酸分解,降低肉鸡腹脂率及肝脏的TG水平,缓解肝脏损伤并维持其脂代谢的稳定性。
随着胆汁酸盐水平的递增,FASmRNA 的相对表达量在0.055%水平下极显著高于其余水平。0.015%水平与0.035%水平显著高于另两个水平,另两个水平差异不显著。而ACC mRNA 的相对表达量则在0.015%、0.055%水平下显著高于其他水平,0.055%水平表达量最高且大于0.015%水平。这一情况与陆江等[19]得出的最适胆汁酸盐水平为0.06%相比较,可以推测出日粮0.055%水平范围的胆汁酸对于雏鸡脂肪酸吸收转运、生长发育有较好影响。
4 结论
①28 日龄雏鸡SREBF1 mRNA 的相对表达量回肠较高,其次是肝脏和空肠,十二指肠最低;FATP4 mRNA 的相对表达量十二指肠、空肠较高,其次是肝脏和回肠;FAS mRNA 的相对表达量肝脏较高,其次回肠,之后空肠,十二指肠最低;ACC mRNA的相对表达量肝脏较高,十二指肠、空肠次之,回肠最低。肠道和肝脏中mRNA 的表达,不仅与胆汁酸代谢有关,而且与脂质代谢有关。
②28日龄雏鸡SREBF1、FATP4、FAS、ACC mRNA的相对表达量存在“肠段×胆汁酸盐水平”互作效应。
③本试验得出,黄羽肉鸡雏鸡日粮添加0.055%左右水平的胆汁酸盐对于雏鸡肠道整体营养转运蛋白及消化酶的基因表达影响最佳,因此推测在日粮中添加不同水平胆汁酸可以调节脂肪代谢相关基因的表达,以促进脂肪分解代谢和抑制脂肪合成,这在改善肉鸡脂肪沉积方面发挥关键作用。
