海洋渔业资源评估模型的研究进展
2022-02-24丁伟康张亚洲周永东
丁伟康,梁 君,张亚洲,吴 昊,周永东,吴 天
(浙江海洋大学海洋与渔业研究所,浙江省海洋水产研究所,农业农村部重点渔场渔业资源科学观测试验站,浙江省海洋渔业资源可持续利用技术研究重点实验室,浙江舟山 316021)
随着计算机运行的快速提升和模型设计的多样化和复杂化,海洋渔业资源评估由单物种到多物种再逐步发展到空间生态模型的评估[1-2]。渔业资源评估实现了对海洋中鱼虾贝藻类的资源估算与统计,一定程度上保障了海洋科研工作者在实施海洋管理[2]、增殖放流[3-4]、人工鱼礁建设[5-7]等各项工作时能够顺利开展,并对渔业未来健康发展起到了举足轻重的作用。然而,在计算机模型对渔业资源评估不断深化的同时,模型的混乱使用和评估不精确等问题也凸显而出[8]。再者,对现有模型的未来发展方向进行判断、归纳模型中常用的影响因子[9]和区分其使用范畴等亟待解决[10]。因此,海洋渔业资源评估模型是一个值得更加深入研究与进行多角度探讨的课题。
资源评估模型需要大量数据,且对数据质量要求较高,但国内外大部分渔业资源是在数据缺乏的情况下运用模型进行评估的[11]。传统的资源评估多从单物种和单渔具的视角,应用种内关系和单一模型进行简单的定性研究[12],但未对环境影响因子和重要参数加以考虑或先验。因此,研究人员需要取得尽可能全面的数据,并对存在误差的数据进行必要的补充调查,才能避免结果出现较大的偏差。
渔业资源的评估不仅需要结合统计学和数学量化的方法,还需要基于物种的生活史,对检测、监测调查和商业捕捞数据进行整合,再通过模型开展有效、合理、准确的评估[13]。我国为了养护和恢复近海渔业资源,设置了不同类型的海洋自然保护地,其渔业资源保护的好坏亟待实施评估。比如,海洋牧场的建设使资源量呈指数型增加[14],在资源评估中运用合适的模型,促进了模型从单一向多方位的转变。同时,国外许多研究也奠定了一定的历史地位。例如,日本学者土井在资源评估方面的研究(图1),为我们进一步构建资源评估框架起到了重要的启示作用[15]。
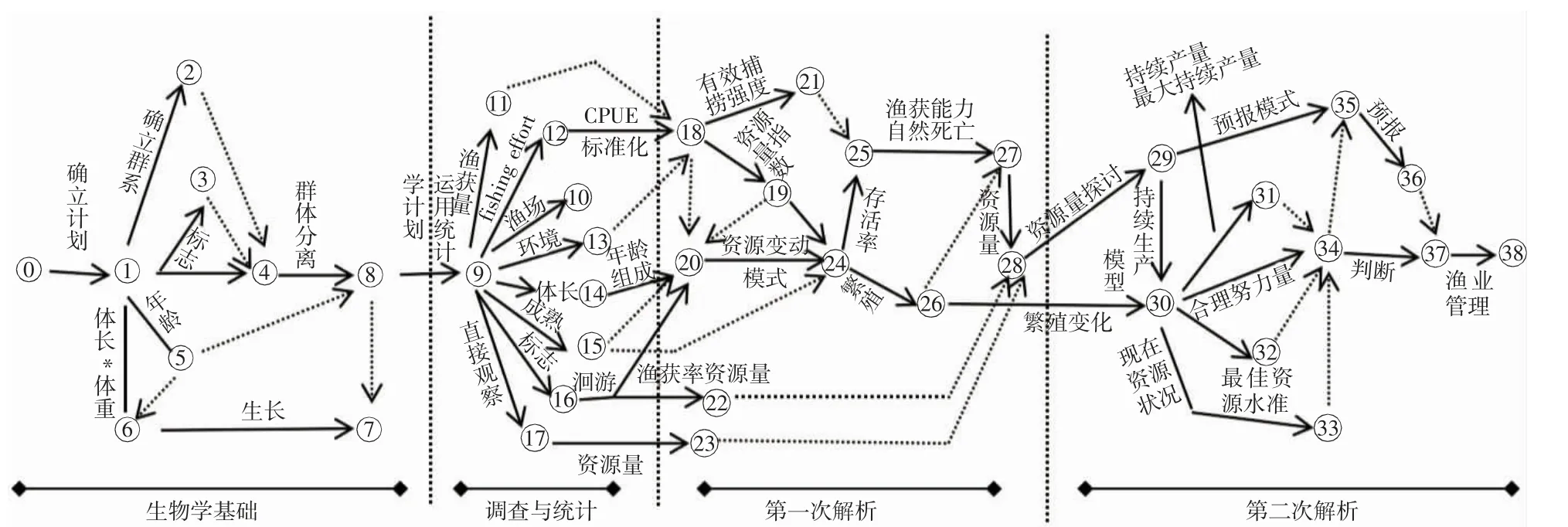
图1 资源评估矢量图(仿自土井[15])Fig.1 Resource evaluation vector diagram
国内外学者开展了大量渔业资源评估方面的研究。在“中国知网”搜索关键词“资源评估模型”检索到学术论文59 篇。其中,“渔业缺乏数据资源评估”的学术论文3 篇,“渔业资源评估研究进展”的学术论文8篇。在“web of science”搜索关键词“resource assessment model”检索到学术论文455 篇,其中,“fisheries lack resource assessments”的学术论文8 篇,“research progress of fisheries resource assessment”的学术论文19 篇。通过对国内外渔业资源评估的文献进行归纳分析可知,有必要对渔业资源评估模型的系统性整理。
1 海洋渔业资源评估模型的逐步建立与发展脉络
20 世纪50 年代的著名鱼类学家GULLAND 曾说“从三大学说开始,关于渔业资源评估的争论就层出不穷,但是对于它的定义却一直未有定论”。詹秉义等[16]在《渔业资源评估》中认为“渔业资源的评估就是利用种种方法对渔业进行评估或估算”。模型是渔业资源评估的载体,一定程度上能保障评估的高效性和科学性。随着计算机运算能力的不断加强,渔业资源评估模型也在不断完善。本研究按照时间顺序(图2),对渔业资源评估模型的发展脉络分为3 个历史阶段,即基础阶段、发展阶段和多样化阶段。

图2 海洋渔业资源评估模型的发展脉络Fig.2 The historical development of marine fishery resource effectiveness assessment model
基础阶段(19 世纪90 年代—1970 年):渔业资源评估历史最早可以追溯到19 世纪50 年代,当时模型发展较为缓慢,大多数模型基于生长方程和简单的数量变动进行评估。到了19 世纪90 年代,第一次世界大战后,耿克(HEINCKE)、彼得逊(PETERSEN)、约尔特(HJORT)3 人相继提出的3 大学说:繁殖论,稀疏论和波动论,成为了当时最为鲜明的资源评估的三面旗帜,开启了渔业资源评估的新元年[17]。随后1918 年巴拉诺夫运用数学分析法研究了捕捞对种群数量的影响[18]。在此基础上Russell 于1931 年建立了简单模型来说明捕捞过度的问题。伴随着(非线性)Von Bertalanffy 生长方程的建立,更加奠定了渔业资源评估的模型化。式(1)体长和式(2)体质量的模型有效的描述渔业的生长变化规律,但是对于某些特定鱼种还需再验[19-22]。
式中:分别为生物体的极限体长(理论最大体长)和极限体质量,cm 和kg;K 为生长参数;t0为理论上体长和体质量等于零时所对应的年龄(一般为负值)。
之后诸多学者建立了季节性VBGH 生长方程[23-25],证明了季节性生长方程比一般生长方程估算参数效果更好。Logistic 曲线最初是运用于人口和生物增长[26-27],GRAHAM 首次把它运用到了渔业评估中,SCHAEFER 基于GRAHAM 的鱼类S 型群体增长曲线建立了非平衡剩余产量模型(SCHAEFER),阐明了当资源处于中等水平时,采取适当的捕捞可以加强最大持续产量[17]。
式中:g(B)种群维持在平衡状态下的捕捞量,kg;r 为内在生长率,%;B 为生物量,kg;k 为平均未开发的平衡生物量,kg。
发展阶段(1971—1980 年):渔业资源评估矢量图[15]的建立标志着渔业资源评估的快速发展,为之后模型加入各种参数评估提供了良好的基础。B-H(Beverton-Holt)模型[28]的建立最初在1957 年,CLAYDER 于1972 年在经典动态综合模型上建立了现代综合模型,此模型从种间和种内关系考虑,建立多鱼种渔业的B-H 模型和自食性鱼类的动态模型。GULLAND 在巴拉诺夫和BEVERTON 和HOLT 研究的基础上建立了年龄实际种群分析VPA(virtual population analysis)模型[29]。之后POPE 对VPA 模型进行简化得到年龄结构股分析(cohrt analysis)CA 模型,也有学者称实际年龄结构股分析(TCA)模型[18]。JONES 又在POPE 的年龄结构世代分析模型基础上建立了体长股分析模型(length based cohort analysis,LCA)[30]。
系统化阶段(1981 年—至今):联合国粮农组织(FAO)以GULLAND 为首的专家在1980 年首次来我国对各水产学者进行资源评估培训,这次培训不仅对我国资源评估的普及和推广起到了良好作用,还为国际上资源评估模型的多元化建立和使用开启了新篇章[31]。多物种实际种群分析MSVPA(Multispecies Virtual Population Analysis)模型从建立至今一直在不断更新,是多物种常用的评估模型。Polina 建立了生物量生产和损耗平衡模型。SEAPODY 在ECOPATH 的基础上增加了模拟生态系统在时间上的动态模块,建立了Ecosim 模型,随后2 个模型相结合形成了EWE 三维模型[32],至今经常被使用在生态系统资源评估中。OSMOSE(object-oriented simulator of marine ecosystems exploitation)原本是基于个体的多物种模型,后面不断演化成了End-to-End 模型来模拟整个生态系统的动态变化[33]。延迟差分模型中的(depletion-based stock reduction analysis)DB-SRA 模型结合了(depletion-corrected average catch)DCAC 模型和(stock reduction analysis)SRA 模型,该模型可以从历史的渔获量中得到相关的信息,并且可以缩减关于持续产量的问题的不确定性,及一些参数的不确定性,比如,MSY(最大可持续产量)和K 值[34-35]。
2 海洋渔业资源评估模型的分类与解析
据不完全统计,全世界90%的渔业资源缺乏相关评估数据[36]。2013 年,美国波士顿举办世界渔业资源评估方法大会时,将“数据缺乏的渔业资源评估方法”列为四大主要议题之一[33]。渔业资源评估在近200 年的发展历史中不断演变[17],现阶段单物种模型在运用中占据多数[37]。渔业资源评估模型大多分为4 种:产量模型、延迟差分模型(delay-difference)、年龄-结构模型(age-structured models)、消耗模型(又称“消散模型”,depletion models)。每种模型包括种群动态模型、目标函数、投影或预测模型和观测模型4 个子模型[35]。
本研究按单物种模型、多物种模型和生态系统资源评估模型3 个方面对模型进行划分(图3),加入重要参数的估算与方法以及相关环境因子[17],并进行解析,以供今后参考并运用最适宜模型对资源进行评估。
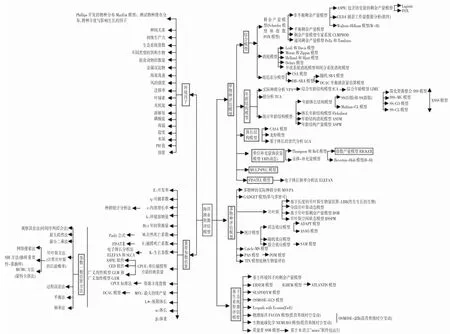
图3 海洋渔业资源评估模型分类Fig.3 The development of marine fishery resource effectiveness assessment model
2.1 单物种评估模型
单物种评估模型主要分为综合模型、年龄结构模型和动态综合模型(单位补充量渔获量模型YPR)三大类[33,35,38]。
2.1.1 综合模型
综合模型包括剩余产量模型、延迟差分模型和消耗模型三大类。综合模型需要统筹考虑生物种群之间的生长、死亡和单位补充量,把着重点放在总产量数据、总数量的动态变化、CPUE 和时间等的关系上[33,39]。
剩余产量模型因简单实用,所需数据少,因此得到了不少学者的广泛使用,且它又分为平衡与非平衡2 种[40]。平衡剩余产量模型因忽视年龄和体长信息而受到质疑[8,40]。ASPIC(a stock production model incorporating covariates)属于非线性的非平衡剩余产量模型,其计算比平衡剩余产量模型相对复杂[41-44]。4 种非平衡剩余产量模型(W-H 模型、SCHNOUE 模型、D-Fox 差分模型和I-Fox 积分模型)使用较多[45-47]。由于SCHAEFER 模型种群动态与实际不对应,相比之下FOX 模型的评估结果更为稳定[40-41,48-49]。
Pella 与Tomlinson 给出了剩余产量模型的一般形式,也称通用剩余产量模型[40,50]。
式中:Dy为y 年的生物量,kg;My为y 年的剩余产量,kg;n 为形状参数;K 为承载能力;r 为内禀增长率,%;Zy表示y 年的总渔获量,kg。当n 为2 时,公式(4)为Schaefer 模型,当n 为1 时,公式(5)为FOX 模型。
剩余产量模型专家系统是由联合国粮农组织专门开发出来并针对东海鲐鲹类(鲐鱼Pneumatophorus japonicus、蓝圆鲹Decapterus maruadsi 等中上层鱼类)最大持续产量(MSY)作评估分析的[46,51]。该系统内大多包含一个环境,连续12 a 以上的产量和捕捞努力量数据参数。最大的特色是增加了交互式问答方式,提供了一个最佳拟合模型[45]。在产量模型中有2 个特殊的模型,分别是限制产量模型和无限制产量模型,两者皆可同时用于估算渔获量和捕获量等相应参数,处理一些产量模型中难以解决的问题[52]。
延迟差分模型进一步拓展了剩余产量模型,加入了生物参数和生物过程中的时间延迟,但是当数据获取不充足时,无法精确估算模型参数,还会产生错误的估算值,较少用于渔业当中[18]。NEWMAN,et al[53]证明了DCAC 是剩余产量模型的延伸并且同样适用渔业数据缺乏的情况。
我国渔业资源评估常用剩余产量模型和年龄结构模型,消耗模型使用的却很少[47]。经典的消耗模型[39]包括了Leslie and Davis 模型[54]、Delury 模型[55]、Zippin 模型[8]、Moran 模型[56]、Helland 模型和Hjort 模型[18]。消耗模型可分为闭合和开放2 种[57],其中,闭合系统的消耗模型未研究关于随着时间的种群动态变化情况,缺点是无法对生物学参考点进行估算。例如,Delury 模型是典型的闭合系统消耗模型,在数据缺乏的情况下比较适用,但是它也存在诸多不足之处,尤其是无法估计对资源与环境之间相互关系的影响程度,表现在对自然死亡系数(M)的敏感度较强,以及生物学参考点计算困难等,当利用该模型评估小型中上层的鱼类资源时,应特别注意环境的变化可能会造成资源空间分布和资源丰富的误差[58]。
2.1.2 年龄结构模型
实际种群分析VPA 和股分析CA 是相对简单的年龄结构模型。实际种群分析VPA 模型是渔业资源评估的经典模型之一[59-61]。股分析CA 方法又是VPA 模型的简化模型,它是将传统VPA 的非线性计算方法转变为线性方法[62-63]。综合年龄结构模型ICA 是基于VPA 的转变模型,仅需要总渔获量(年度)、各年龄组渔获量、资源量平均体质量、自然死亡系数(M)和年龄成熟度等数据就可评估中上层渔业资源量[37]。若渔获量和体长2 种数据都具备,则可用LIME(length-based integrated mixed effects)模型进行评估。如果只有体长数据时,此模型只适用时间序列超过1 a 的渔业资源评估[64]。
PUNT,et al[65]等研究发现剩余产量模型在加入年龄结构种群特征后,该模型更具可信度和灵活性,并取名为“年龄结构产量模型”。XIAO Yongshun,et al[66]报道了年龄结构的产量模型ASPM(age-structured production models)和年龄结构的消耗模型ASDM(age-structured depletion models)。
最初SS 模型(stock synthesis,最新版本为SS3)的评估特点是任一年龄的某一体长组的消失,都不会影响该年龄下一时刻的体长分布概率[67]。简化资源整合模型SSS(simple stock synthesis)是基于体长的综合混合效应模型[68]。而最新SSS 模型是基于DB-SRA 模型和SS 模型调整所得。SSS-MC(simple stock synthesismonte carlo)模型对资源损耗状况特别敏感,所以研究者所提出的XSSS(extended simple stock synthesis)模型运用了贝叶斯参数估算,它更接近标准的年龄结构资源评估模型,但是也只适用于有丰度指数的渔业资源评估[69]。
体长结构模型是未来资源评估的开发方向,所以在往后的渔业资源评估中应记录好体长结构数据。此模型的特点是在资源评估的全过程不仅可以优化年龄体长数据分布,且所有过程都可在体长数据上进行。而对于基于体长的年龄结构模型来说由于年龄数据较难获取,大部分数据存在一定误差,所以并不适于评估寿命较长的甲壳类水生生物资源[38]。不乏有学者将莱斯利矩阵的年龄结构模型转化为体长结构模型[39,65],如现今广泛被采用的CASA(Carnegie-Ames-Stanford Approach)模型和加入了分季节性和性别特定的体长结构模型-龙虾模型[8,38]。
2.1.3 动态综合模型
此类模型可以根据已知的生长率和死亡率,计算出首次捕捞年龄和捕捞死亡率。其中,B-H 模型和RICKER 模型在国内使用较早,运用较多[70]。B-H 模型是基于一个稳定状态的模型,它是在捕捞强度和开捕年龄不变的情况下,分析对资源和渔获量所产生的影响。而且,B-H 模型主要适用于单物种渔业资源评估,不考虑其它鱼种,这样评估结果往往受限。特别是南海区,以拖网作业为主,一次捕捞会有多个鱼种,有些开捕年龄适合,而有些不适合,且捕捞渔具所引起的死亡系数也大不相同[28]。综上,动态综合模型的计算方法是在一种理想状态下进行,如果出现过度捕捞,资源量里的补充量就会下降,生长率就会增长。
2.2 多物种评估模型
传统的渔业资源评估一般基于单物种模型,单物种模型的建立让我们认识到资源评估模型是对资源评估有利又有效的工具。单物种模型在实际评估中不是很适用,因为存在兼捕效应,也就是在实际捕捞中,我们不可避免地捕获到非目标种类或不符合要求的目标种类。所以出现了多物种评估模型,多物种的资源评估模型是在单物种资源评估模型的基础上发展而来。例如,多物种实际种群分析MSVPA 是基于单物种的VPA 所建立的评估模型,也是最常见的多物种渔业资源评估模型[8,71]。在传统的单物种模型中自然死亡系数(M)一般都设为常数,而在实际水生生物活动中,死亡系数是个变量,所以在多物种的资源评估中我们不仅要考虑M,还要考虑种间关系、捕食与竞争关系等各参数之间的关系,才能更加准确地估计M,提高渔业资源评估的准确性[39,62]。因此,GADGET 模型、Catch-MSY 模型、PSA 模型和POM 模型等兼容度较高(既适用于单物种,也适用于多物种)的模型应运而生。由于生态系统的复杂性,缺少必要的数据支持,多物种的评估模型至今尚不能成为渔业资源评估的主流模型。
GADGET(globally applicable area-disaggregated general ecosystem toolbox)模型具有与SS3 类似的功能,它不仅可用作单物种渔业资源评估,也可用作多物种渔业资源评估[72]。Catch-MSY(catch-maximum sustainable yield)模型在评估精度上有时无法取代标准资源评估模型。但我国近海大部分渔业都是处于数据缺乏的状态,所以此模型在多物种渔业资源评估中应用前景良好[73-74]。使用过程中需要注意的是参数的错误对评估结果很敏感,在渔获量波动大的渔业中需谨慎使用。PSA(productivity susceptibility analyses)模型多用于计算多种群的相对易损性[75]。与其相似的是POM(pattern oriented modeling)模型,此模型仅在美国淡水湖泊中使用过,评估时需要多个鱼种的不同生长阶段的生物学参数,由于参数获取困难,大部分鱼种的资源评估不满足这个条件[76]。
夏日苍白的光照下更显翠绿。铁手里又落下一段灰白的骨头,在红褐色的土壤映衬下,非常耀眼,看起来就像浸在血液中。汤翠吐了一阵,又开始哆嗦。不是乱坟场吗,怎么就这几根骨头?
2.3 基于生态系统渔业资源评估模型
FAO 公布的世界渔业统计数据显示,在全球大面积捕捞和海洋工程建设下,捕捞网具的网目逐渐减小,包括不正确的捕捞方式,如底拖网捕捞[31]。不合理的捕捞方式导致了海洋生态系统遭受到了严重破坏,海洋内渔业资源加速枯竭[77]。
虽然如今的单物种估算模型在更新发展中也加入了物种间的捕食关系和一些简单的环境因子,但研究重点还在单物种目标上,与生态型模型仍有所差距[77-78]。生态系统模型构建的意义不仅在于它可以描绘生态循环模式,能够分析解释生态过程和机理,更能让我们认识到海洋生态系统的结构动态[12]。它包含了从初级生产者到顶级捕食者在内的全部营养级,生物所需的栖息环境和营养化学元素循环动态,以及气候变化和人类活动的影响[79]。EwE 模型、ATLANTIS 模型和OSMOSE 模型等是基于生态系统渔业资源评估模型的典型代表。
2.3.1 EwE 模型
EwE 主要由Ecopath、Ecosim 和Ecospace 三大模块构成[11,80]。Ecopath 是一个静态模型,并且它的结果可用于驱动Ecosim 和Ecospace 进行评估,再通过简化参数得出在生态系统大背景下种群的数量[81]。
Ecopath 模型可以描述生态系统食物网关系,是最常用的生态系统模型。常常估算水生生态系统生物量和食物消耗[73]。Ecosim 模型动态模拟了捕捞产量和资源量在既定时间内随着环境与人为的影响所造成的变化,说明了生态系统营养交互过程。Ecospace 模型是由用户提供相关功能组的栖息地、捕捞和保护区域的信息,再进行空间分析[82]。
2.3.2 ATLANTIS 模型
ATLANTIS 模型属于生物地球化学系统模型,它依然是属于生态系统模型的延伸,是在IGBEM 模型基础上建立的,而IGBEM 模型又是整合了ERSEM 模型,加入了物理过程。由于IGBEM 本身就可以模拟底栖生物、浮游动植物、微生物和碎屑等群落中的各元素循环过程[83]。ATLANTIS 就更不同于一般模型,此模型结构复杂、运算量较大、参数设置繁琐、需要大量数据支撑且估算时间较长,对于计算结果验证也比较困难,所以对相应群体补充过程敏感性强[33]。
2.3.3 OSMOSE 模型
OSMOSE 能够模拟从低营养级到高营养级物种的完整生态系统动态,主要构建种群食物网以模拟摄食动态变化和营养级相互作用,对观测数据的依赖较低,适于大尺度和多物种的生态系统研究[84]。
OSMOSE-JZB 模型也是一个端到端模型,并耦合了FVCOM(finite volume ocean model)模型和NEMURO(North pacific ecosystem model for understanding regional oceanography)模型,它囊括了自浮游生物到顶端捕食者在内的所有生物。FVCOM-NEMURO 用来描绘低营养生物时空动态变化,OSMOSE 用来描绘物种的生活史和鱼群生物学进程。此模型与其他生态模型一样,要对参数进行优化估算,避免模型不当使用[85]。
虽说基于生态系统的模型经过重要参数和环境因子的加入,可以缩小模型对资源评估的误差范围,但是参数的计算量的增加不仅相对繁琐,很可能伴随数据的不准确性,导致结果的高估和低估[86-87]。
2.4 各模型的适用性与优缺点
在众多的模型中,每种模型对应的物种不同,所适用的评估模型也会不同。就模型本身而言并没有好坏之分,是因各模型所需假设和数据需求不同而所选的适配模型不同[8,88],所以模型在评估使用时还需先验考虑。在不同区域、不同物种和不同环境条件下,模型的运用大不相同[89-90]。本研究总结了前人的研究成果和对模型的运用,阐述了各个模型的类型、所需数据、应用物种和海域、优缺点,详见附录表https://docs.qq.com/doc/p/8c0f911ee2f935c3f18ac30ccfc3bad47fae4d70。
根据附录表分析可知,对于单物种资源评估模型在不易测定年龄、不易区分渔获物组成的种群和小型中上层鱼类资源评估时使用平衡剩余产量模型,特别是热带渔业资源评估可使用非平衡的剩余产量模型[37]。剩余产量模型尽管因忽略信息(如,种群的年龄或体长结构等)而受到质疑,但因其具有数据与参数需求少、便于理解与执行、模型直观等优势,还能提供MSY 等渔业管理所需的生物参考点,在渔业资源评估中一直被广泛应用,但建立过程中应该根据不同阶段情况进行不同分析。延迟差分模型是剩余产量模型的补充,在应用延迟差分模型时需要通过耳石等间接获取年龄,但耳石鉴定又容易产生误差,势必会增加渔业资源评估的不确定性[8,48],模型中若增加生长、自然死亡、补充量等重要参数,估算结果将会更为精确[1]。年龄结构模型一般只在数据丰富的前提下使用,若用于评估短生命周期水生生物尚有局限性[1]。消耗模型使用场景有局限性,推荐在对甲壳类生物资源进行评估时使用。
对于多物种资源评估的模型中MSVPA 也是比较常用的,但是它本身还是基于单物种的演变,如果物种间复杂关系可能会导致结果会有一定偏差。而Catch-MSY 模型与其他方法相比需要较少的数据,并且Catch-MSY 模型对最大可持续产量(MSY)的估计较为稳定,与完整的资源评估模型的评估结果一致,在数据缺乏下的中国近海渔业中运用更广泛,所以相对来说Catch-MSY 在多物种模型中更为适用[34]。
对于发展生态模型的意义主要有三部分:其一,解读生态模式;其二,对生态过程和机理的进一步明析;其三,预测系统动态[83]。EwE 是当前国内外使用最广泛和最典型的生态评估模型,它不仅可以简化参数估算过程,还能将时间与空间较好地结合起来,准确评估资源数量。所以对于基于生态系统的模型评估EwE 模型比较适用。
3 结论
根据以上对模型的分类总结和解析,我们不难得到结论,国内外对于评估模型的运用各不相同,不管是中上层渔业[1]、甲壳类[38]、长寿命[83]、热带鱼类和短寿命鱼类[8],还是从浮游生物到顶级生物[83]、底层鱼类[91]等水生生物都各有不同的模型进行评估。因为每种模型所需的输入数据和应用物种与海域都大有区别,所以大部分鱼类的评估都存在数据缺乏的不足之处,而造成这种数据缺乏的原因也有多种,本研究具体归纳为3点:(1)渔业的相关基础统计数据缺乏;(2)记录数据失真;(3)只有较少海域有部分资源数据[68]。所以在模型使用上,不断趋向于在数据缺乏下的非线性模型运用。模型在使用时进行先验和后验也是很重要的,不仅可以减少回顾性问题RP(retrospective problem)的出现,还能精确估算资源量[92]。
如今模型在渔业资源评估中仍存在着一些较为复杂的问题,本文将其大致归纳为2 类:(1)RP 问题。RP 问题的产生是因为输入数据错误和模型假设错误,在资源评估中所运用大部分的模型都存在着RP 问题,因为渔业数据的缺失是常见的现象,所以在实际运用中要多加思考数据量和模型的匹配度。(2)重要参数估算。单物种评估模型也在不断地加入一些常见的参数,多物种与生态系统评估模型更是如此。例如,自然死亡率参数的设置,如是设置为常数,就可能会造成评估结果的不准确,且也要注意个人在填入参数时的主观性。所以在进行参数估算时不仅要运用正确的方法还要避免过度参数化,防止增加模型复杂性和影响模型的灵活性[92]。
4 展望
从历史发展脉络来看,虽说模型的发展不断在进步和填充,各项因素的把控和精确化的演算减少了资源的估算差幅,但是我们依然都还是在前人的基础上进行革新。根据本文研究总结,我国海洋渔业资源的评估今后很长一段时间内还是以单物种模型评估为主,同时采用多物种模型,并不断发展生态系统模型。在底层渔业资源不断衰退的大背景下,国内大部分渔业资源评估都集中在中上层海域,所以未来模型的整体发展还是会基于生态系统模型的建立与完善。未来评估模型对不同海域和不同生物的测定估算会加入更多环境因子和重要参数,对相应的重要渔业对象和重点渔业区域还需要加入贝叶斯方法以提高评估的准确性。
总体而言,尽管生态模型发展是趋势,但近年来的生态模型大多仍是基于前人的模型与理论建立与应用,且它还不能够完全真实的反映生态系统的全部过程。不同模型往往对生态系统的不同组分有所侧重和偏向,模拟和解答生态问题也有所差异,在实际中需针对科学问题,选择合适生态模型。LEVINS 对未来模型发展和选择给出了较好的解释框架:体现生态真实性和精确性的经验模型,体现生态精确性和普适性的分析模型和体现生态普适性和真实性的机制模型[93](图4)。在未来,模型将更好地将社会、经济和生态情况结合起来开展,所以此结构图能对未来模型发展和资源评估起到很好的理论支撑作用。
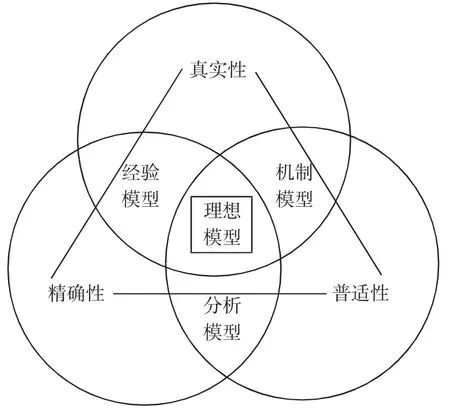
图4 三分法模型选择框架Fig.4 Trichotomous model classification scheme
