舟山岛北部海域虾蟹类物种多样性的季节变化研究
2022-02-24俞存根蒋巧丽牛威震许永久
汪 全,俞存根,郑 基,蒋巧丽,牛威震,许永久
(浙江海洋大学水产学院,浙江舟山 316022)
舟山渔场位于长江口南端和杭州湾口外侧,海域受江浙沿岸水及台湾暖流的季节性交互消长影响,特别是受长江和钱塘江等大陆径流携带而来的大量陆源物质的影响,致使该海域水质肥沃,饵料生物丰富,渔业资源种类繁多。历史上,调查海域东南部的黄大洋渔场,主产银鲳Pampus argenteus、鳓Ilisha elongata、曼氏无针乌贼Sepiella maindroni 等,西北部的灰鳖洋渔场,主产银鲳、鳓、蓝点马鲛Scomberomorus niphonius 及鮸Miichthys miiuy 等,曾经是众多传统经济鱼类的生息区和舟山渔场的主要捕捞作业区[1]。但是,由于受多年来的过度捕捞、环境污染及栖息地破坏等影响,该海域的传统鱼类资源相继衰退,渔业生物群落结构发生明显变化,低质低值的小型鱼类及虾蟹类逐渐替代了原来的传统底层经济鱼类资源等[2]。针对舟山渔场渔业生物群落结构变化,以往有不少专家学者曾对舟山渔场不同区域的渔业资源进行过调查与研究,如俞存根等[1]根据2006—2007 年春夏秋冬四季对舟山渔场大面调查资料,系统阐述了舟山渔场的鱼类、虾蟹类及头足类等生物群落结构演替趋势及生物多样性变化;何贤保等[3]分析了灰鳖洋渔场春秋季渔业生物多样性;倪海儿等[4-5]解析了舟山渔场渔业资源动态及其主要渔业资源利用现状。就虾蟹类而言,也有不少研究报道,如郑献之等[6]报道了舟山渔场及邻近海域蟹类种类组成和时空分布;何贤保等[7]分析了舟山渔场及附近海域虾类群落结构特征;瞿潇南等[8]研究了舟山黄大洋海域春秋季虾类种类组成和数量分布;李德伟等[9]研究了舟山渔场官山附近海域春秋季虾类群落结构特征;张平等[10]分析了舟山近岸海域虾类种类组成与数量分布及其变动趋势;刘惠等[11-12]研究了舟山近岸海域主要甲壳类的生态位及其种间联结性,以及舟山沿岸渔场甲壳类群落结构特征;胡嘉豪等[13]分析了舟山群岛以东沿岸海域虾类资源状况及其多样性;近年,于南京等[14]、牛威震等[15]、刘坤等[16]研究了舟山群岛东侧海域虾蟹类群落结构及其与环境因子的关系、主要甲壳类物种的空间生态位。但是,对于地处舟山岛以北、岱山岛及长涂岛之间的灰鳖洋与黄大洋两大传统渔场的虾蟹类群落结构及其生物多样性却少有报道,特别是缺乏利用周年四季的数据研究虾蟹类生物多样性的季节变化。然而,该海域不仅是舟山海域的传统渔场,也是近年在海洋大开发背景下人类干扰活动频繁、各类海洋工程建设较多,海洋生态受到威胁的区域。本文利用2020 年10 月、2021年1 月、4 月、6 月在舟山岛北部海域开展渔业资源底拖网调查所获得的虾蟹类资料,分析该海域虾蟹类的种类组成、生态类型、生物多样性等群落结构特征,以掌握调查海域虾蟹资源利用、生物多样性及群落结构动态变化状况,为舟山渔场渔业资源可持续利用及各类涉海工程项目建设对海洋生态环境影响评价提供参考依据。
1 材料与方法
1.1 调查海域与采样方法
文章所用数据来自2020 年10 月(秋季)、2021 年1 月(冬季)、4 月(春季)、6 月(夏季)租用“浙普渔32207号”渔船在舟山岛北部海域开展渔业资源底拖网调查,调查船主机功率为330 kW,调查网具规格为860目×80 mm。调查范围为29°59'~30°22'N,121°54'~122°33'E,调查海域中设置站位共20 个(图1)。调查方法按照《海洋渔业资源调查规范》(SC/T 9403-2012)进行。在每个站位拖曳作业保持3.5 kn的拖速,持续时间约1 h。通过调查船上的水深仪测量并记录各站位水深,用实验室样品袋将各站位获得的渔获物全部装取放入冰库保存,并记录站号对应的袋号,带回渔获物后在实验室内进行种类鉴定及生物测定,体长和体质量分别使用量鱼板和电子天平进行测量,电子秤的精度为0.1 g。

图1 调查站位示意图Fig.1 Schematic plot of investigation station
1.2 数据处理与分析
1.2.1 优势种确定
采用相对重要性指数(IRI)确定优势种,计算公式如下[17]:
式中:ni、wi分别为第i 种虾蟹的尾数和生物量,ing.、g;N 为调查所获虾蟹类的总尾数,ind.;W 为总生物量,g;fi为第i 种虾蟹在m 次取样中出现的取样次数;m 为总站位数。
1.2.2 生物多样性指数
采用Shannon-Weiner 指数(H')、Pielou 指数(J')、Margalef 指数(D)分析虾蟹类群落结构的生物多样性,具体公式分别如下[18-19]:
式中:Pi为第i 种虾蟹类的尾数占总尾数的百分比或质量占虾蟹类总生物量的百分比,%;H'为生物多样性指数;J'为均匀度指数;D 为丰富度指数;G 为虾蟹类生物量总和,g;S 为虾蟹类种数总和。
1.2.3 群落结构分析
本研究采用聚类分析,比较4 个季节不同站位的群落相似性。将舟山岛北部海域的虾蟹类原始数据中生物量通过四次方根系数转换,目的是减小群落中优势种和稀有种的影响。在秋季站位调查中由于Z1、Z19 号站位没有采样到虾蟹类,因此群落结构组成分析中除去这两站位。虾蟹类群落结构与季节间的关系用到聚类分析(CLUSTER)和非参数多维标度排序分析(NMDS)进行计算。判断NMDS 二维点阵图分布结果可信度通过胁迫性系数(stress)进行检验,一般认定当stress<0.05 时,结果可信度较好;当0.05≤stress<0.1时,可信度一般;当0.1≤stress<0.2 时,结果在一定程度上具有解释意义[20]。
1.3 数据处理方法
为了便于比较,将虾蟹类的渔获数量统一标准化为每小时的渔获量。通过Excel 2019 软件计算相对重要性指数(IRI),绘制调查站位点分布图用到软件Surfer11,用PRIMER 5.0 软件对虾蟹类群落进行等级聚类(CLUSTER)和非参数多维标度排序分析(NMDS),之后进行ANOSIM 检验[21],利用Origin 2021 软件中单因素方差(ANOVA)分析水深对虾蟹类生物多样性的影响显著性,本文中的差异显著性检验是对不同水深组之间的四季变化进行单因素方差分析。
2 结果
2.1 种类组成
根据2020—2021 年春夏秋冬季4 个航次渔业资源底拖网定点调查获得的虾蟹类样品,共鉴定出虾蟹类25 种,隶属于14 科20 属,其中,虾类有16 种,蟹类有9 种如表1 所示。虾类种类数的周年变化趋势为冬季(12 种)>春季(11 种)>夏季(7 种)>秋季(6 种),蟹类种类数的周年变化趋势为秋季(6 种)>冬季(5 种)>夏季(4 种)>春季(3 种)。其中一年四季均有出现的主要有中国毛虾Acetes chinensis、安氏白虾Exopalaemon annandalei、葛氏长臂虾Palaemon gravieri、日本鼓虾Alpheus japonicus 和三疣梭子蟹Portunus trituberculatus 5种,根据各种虾蟹类的生态习性,可将调查海域的虾蟹类划分为2 种生态类型,即广温广盐型,如细螯虾、三疣梭子蟹等;广温低盐型,如安氏白虾、中国毛虾等。

表1 调查海域虾蟹类种类组成的季节变化Tab.1 Seasonal variations in the composition of shrimp and crab species in the surveyed sea
调查海域主要虾蟹类不同季节的优势度(IRI)如表2 所示。优势种划分归类为IRI 值>1 000 的所有种;常见种划分定为IRI 值介于100~1 000 之间的所有种,主要虾蟹类包括优势种及常见种。调查海域春季优势种依次有细螯虾、三疣梭子蟹、中国毛虾、安氏白虾4 种;夏季有中国毛虾、三疣梭子蟹2 种;秋季和冬季分别有2 种,均为安氏白虾和中国毛虾。
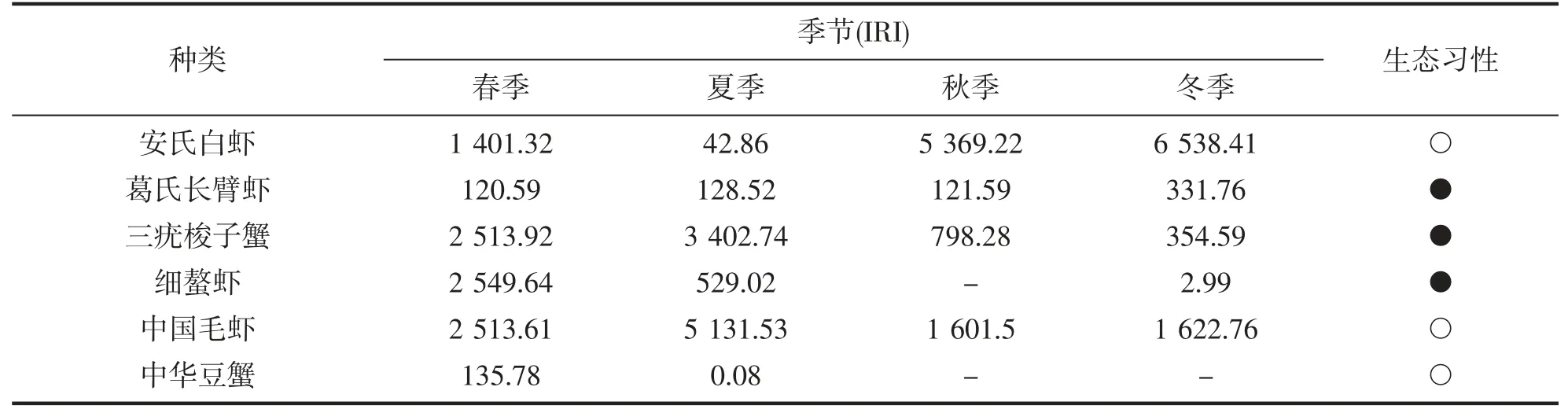
表2 不同季节调查海域主要虾蟹类的优势种及重要种组成特征Tab.2 Changes in the importance relative index(IRI)of shrimp and crabs in the surveyed areas between different seasons
2.2 物种多样性
调查海域虾蟹类的丰富度指数(D)、均匀度指数(J')、生物多样性指数(H')的季节变化如表3 所示,结果显示调查海域虾蟹类丰富度指数(D)值以春季最高,以夏季最低;均匀度指数(J')以春季最高,以冬季最低;多样性指数(H')以春季最高,以秋季最低。夏季丰富度指数(D)平均值全年最低为0.55;秋季(H')平均值全年最低为1.01,与秋季个别站位未捕获虾蟹类有关;春季、冬季(J')平均值相差较大,而夏季、秋季(J')平均值相差较小。
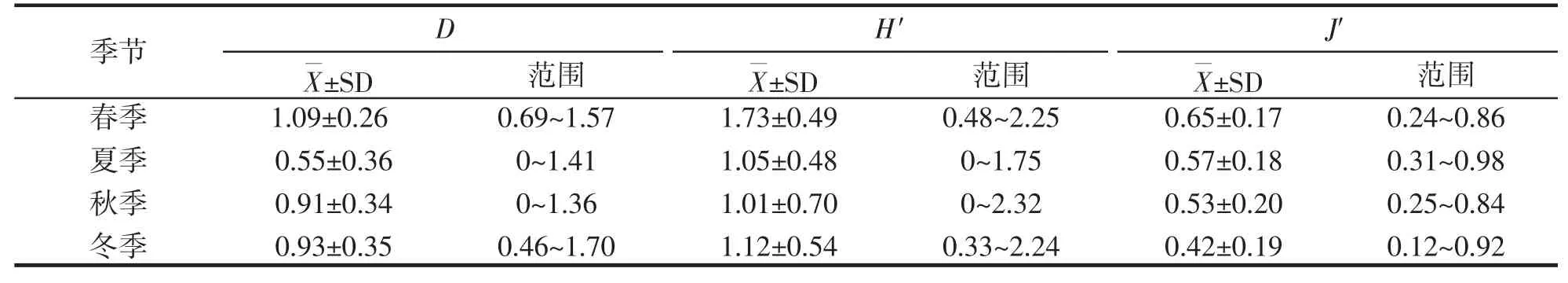
表3 舟山岛近岸北部海域虾蟹类群落的生物多样性的季节变化Tab.3 Seasonal variation of biodiversity of shrimp and crab communities in the northern coastal waters of Zhoushan Island
不同水深带对虾蟹类物种多样性的季节变化如表4 所示。春季,虾蟹类丰富度指数(D)在水深>40 m处最高,在20~40 m 水深处最低;多样性指数(H')在水深>40 m 水深处最高,在20~40 m 水深处最低;均匀度指数(J')在20~40 m 水深处最高,在水深<20 m 处最低。夏季,虾蟹类丰富度指数(D)在水深>40 m 处最高,在水深<20 m 处最低;多样性指数(H')在水深>40 m 水深处最高,在水深<20 m 处最低;均匀度指数(J')在20~40 m 水深处最高,在水深<20 m 处最低。秋季,虾蟹类丰富度指数(D)在20~40 m 水深处最高,在水深<20 m 处最低;多样性指数(H')在水深>40 m 处最高,在水深<20 m 处最低;均匀度指数(J')在水深>40 m处最高,在20~40 m 水深处最低。冬季,虾蟹类丰富度指数(D)在水深<20 m 处最高,在水深>40 m 处最低;多样性指数(H')在20~40 m 水深处最高,在水深>40 m 处最低;均匀度指数(J')在20~40 m 水深处最高。
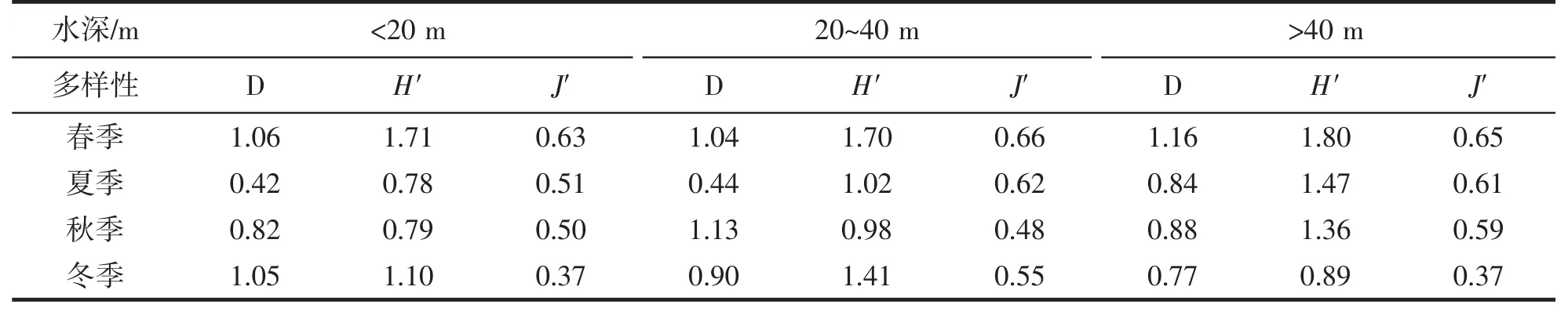
表4 舟山岛近岸北部海域不同水深带虾蟹类生物多样性的季节变化Tab.4 Seasonal changes in shrimp and crab biodiversity in different water depths in the northern coastal waters of Zhoushan Island
虾蟹类生物多样性指数(H')的平面分布如图2 所示。春季,H'值以舟山岛与秀山岛之间海域为较高;以调查海域的西部区域为较低。夏季,H'值也以舟山岛与秀山岛之间海域为较高;以调查海域东部区域为较低。秋季,H'值仍以舟山岛与秀山岛之间海域为较高,以调查海域东北部区域为较低。冬季,H'值以舟山岛东部和西部附近海域为较高,以调查海域北部区域为较低。除冬季外,其他3 个季节的生物多样性指数H'均较集中在秀山岛附近海域。通过ANOVA 检验结果表明,四季H'值平面分布存在极显著差异P=0.000(P<0.01)。不同水深带虾蟹类生物多样性指数H'的季节变化如图3 所示,单因素方差检验表明,虾蟹类生物多样性指数H'在水深<20 m 差异显著(F=3.99,P=0.016<0.05),水深在20~40 m 差异不显著(F=2.15,P=0.134),水深>40 m 差异极显著(F=5.34,P=0.007<0.01)。单一季节不同水深带T 检验分析结果显示,春秋冬季各水深带虾蟹类生物多样性生物指数H'差异不显著(P>0.05),仅夏季存在显著性差异(F=4.83,P=0.02<0.05)。

图2 舟山岛北部海域虾蟹类(H')值空间分布(a.春季;b.夏季;c.秋季;d.冬季)Fig.2 Spatial distribution of shrimp and crab(H')values in northern Zhoushan Island(a.spring;b.summer;c.autumn;d.winter )

图3 不同水深带虾蟹类生物多样性指数(H')的季节变化Fig.3 Seasonal variation of the biodiversity index(H')of shrimp and crabs in different water depth zones
2.3 群落结构特征
舟山岛北部海域虾蟹类群落的等级聚类如图4 所示,非参数多维标度排序分析如图5 所示。聚类分析中胁迫系数如下:春季、冬季分别为0.1、0.12,说明其结果具有一定解释意义;夏季、秋季分别为0.07、0.08,说明其结果基本可信。由于秋季的Z1 和Z19 站位点没有捕到虾蟹类,因此秋季数据只选取18 个站位的数据。春季,在60%相似性水平上,可划分2 个组群,分别为I 组群(Z17)、II 组群(Z1~Z16、Z18~Z20);夏季,在60%相似性水平以上,可划分3 个组群,分别为I 组群(Z19、Z20)、II 组群(Z1~Z18);秋季,在30%相似性水平以上,可划分3 个组群,分别为I 组群(Z9)、II 组群(Z14、Z18)、III 组群((Z1~Z8、Z10~Z13、Z15~Z17、Z19、Z20);冬季,在50%以上相似性水平以上,可划分3 个组群,分别为I 组群(Z19、Z20)、II 组群(Z9、Z17)和III组群(Z1~Z8、Z10~Z16、Z18)。春季I-II 组群相异性最低为69.36%,夏季I-II 组群相异性为93.18%,秋季III、I-III、II-III 组群相异性均在90%以上,冬季I-II、I-III、II-III 组群相异性均在70%以上(表5)。
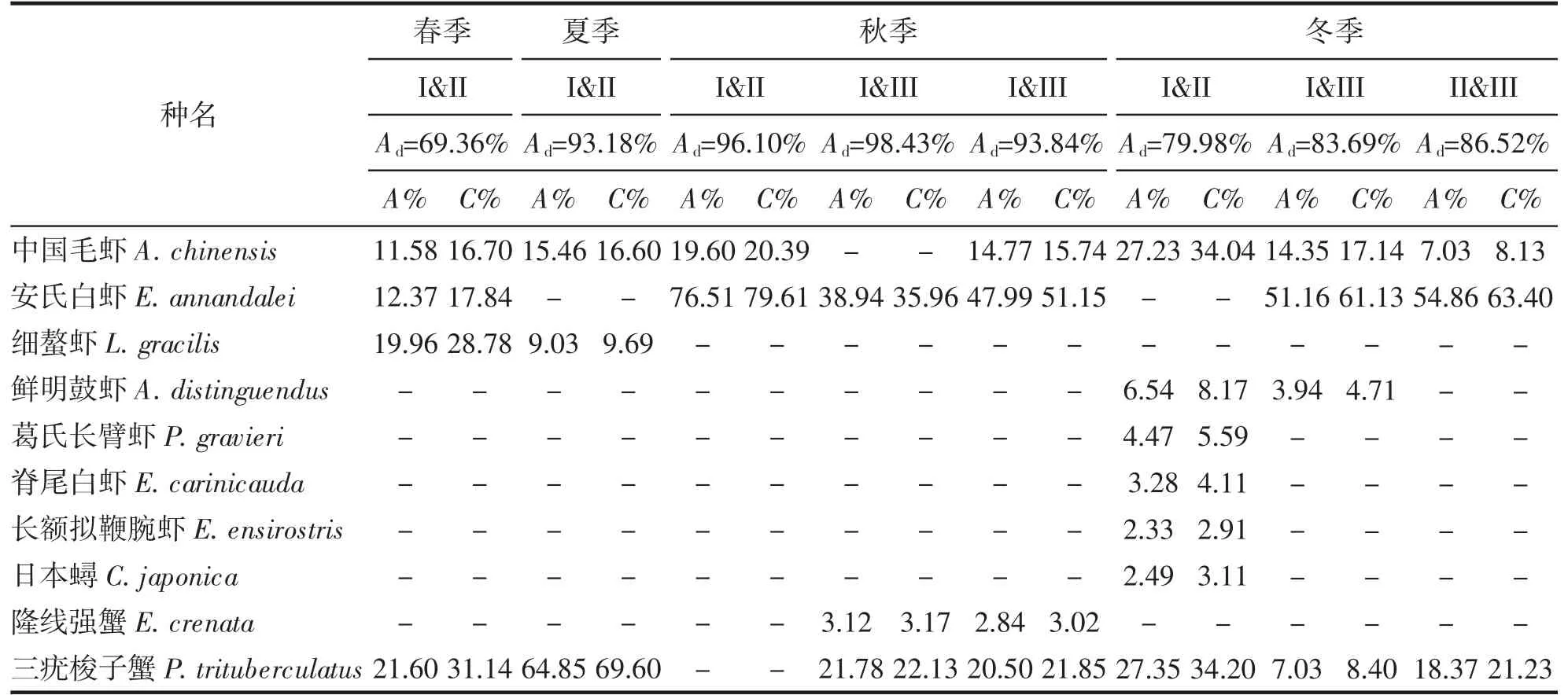
表5 舟山岛近岸北部海域虾蟹类群落内平均相异性贡献率之和占90%以上的特征种Tab.5 Characteristic species with a total contribution rate of mean heterogeneity in shrimp and crab communities in the northern coastal waters of Zhoushan Island accounted for more than 90%
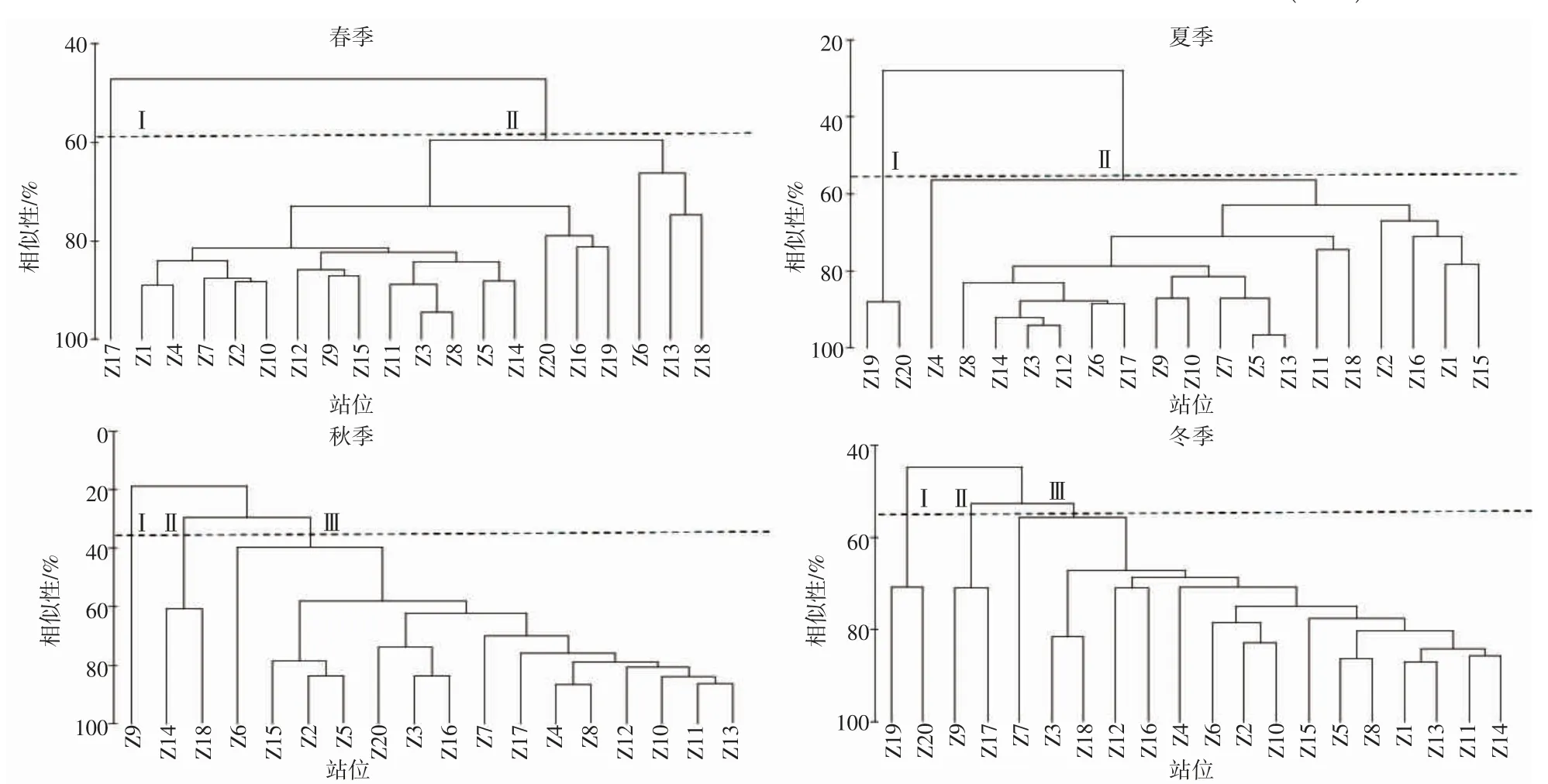
图4 舟山岛北部海域虾蟹类群落的聚类图Fig.4 Bray-Curtis cluster diagram of shrimp and crab communities in northern Zhoushan Island
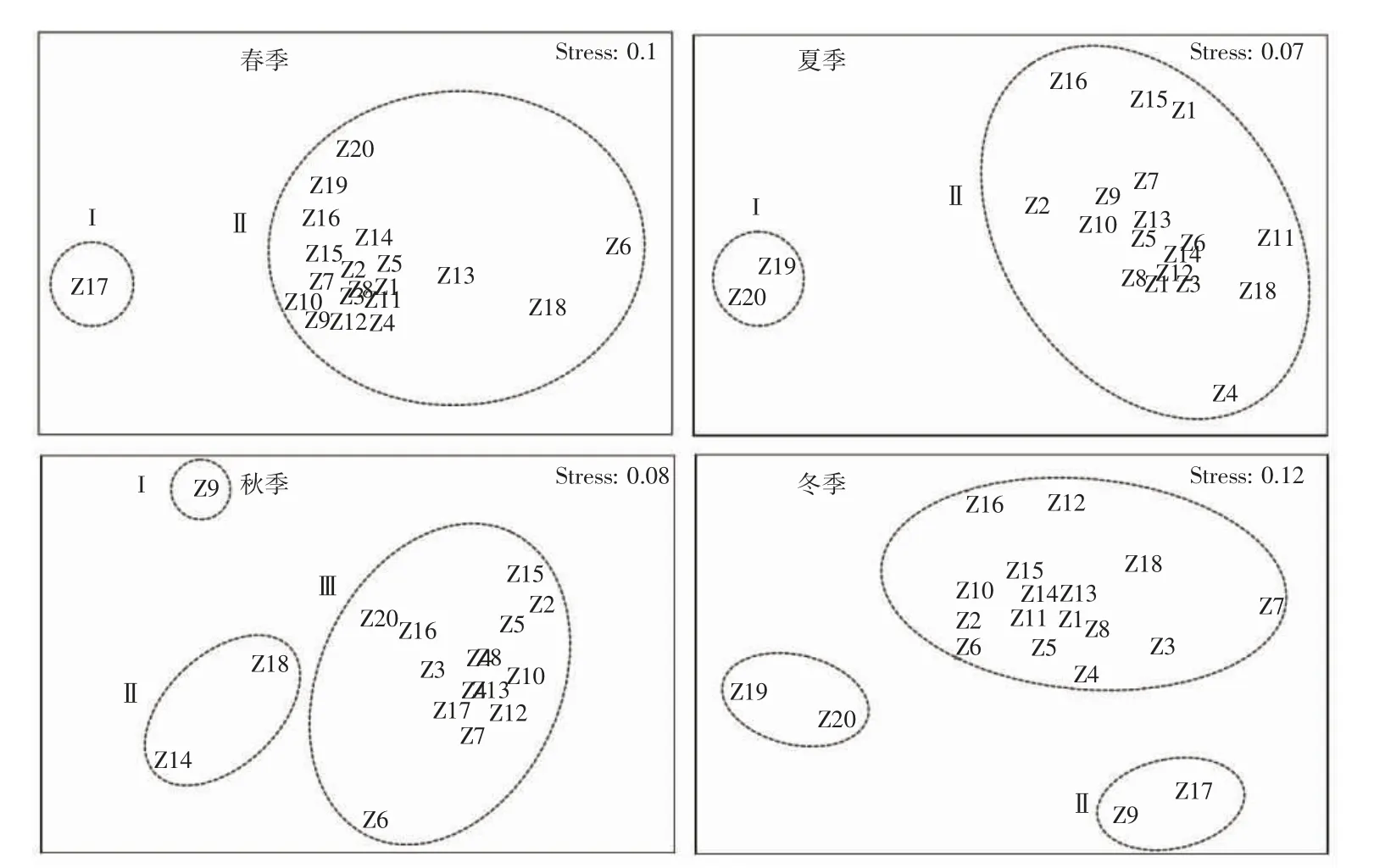
图5 舟山岛北部海域虾蟹类群落的二维点阵图排序分析图Fig.5 Sorting and analysis of two-dimensional lattice plot of shrimp and crab communities in the northern sea of Zhoushan Island
3 讨论
3.1 种类组成和优势种分析
本次在调查海域共鉴定出虾蟹类25 种。从虾蟹类的生态习性上可以将其划分为广温广盐型、广温低盐型两种生态类型。这与郑献之[6]、瞿潇南[8]等研究结果一致。一方面是与舟山岛北部海域的地理位置、底质类型以及水文环境等有关。调查海域的西北部靠近长江口和杭州湾河口区,受大陆径流入海的影响,其水文环境复杂多变[22],特别是温度、盐度年间变化较大,适合广温广盐型或广温低盐型虾蟹类生存繁殖;另一方面也能反映一年内该海域主要优势种虾蟹类的种类组成及适温盐性。从虾蟹类优势种组成及季节变化中分析,该海域的蟹类种类共有9 种,但是,优势种只有三疣梭子蟹1 种,本文推测可能原因为多数蟹类喜栖息于沙质底或者岛礁附近(如日本蟳)水深相对较浅的区域,而本次海域海底底质以泥沙质等细颗粒沉积混合物为主,取样站位又考虑到拖网作业安全,故均离岛礁相对较远。中国毛虾作为一年四季的绝对优势种,不同季节的IRI 值均超过1 000,以夏季的IRI 值(5 131.53)最高。主要原因是中国毛虾一般不作长距离徊游,仅随潮汐涨落与所栖海域水温变化作近岸与远岸的往复移动[23],故调查站位全年均能捕获中国毛虾。春夏季以三疣梭子蟹为优势种,而秋冬季更替为常见种,这可能是三疣梭子蟹在春夏季洄游到水温适宜的近岸海域繁衍生息,而秋冬季随着其生理生长需求以及水温、盐度等环境因子的变化,三疣梭子蟹向其他更适合其生息的海域移动,如秋末冬初,随着近岸水温下降,三疣梭子蟹开始往较深的海域作越冬洄游[24]。安氏白虾除夏季为常见种(IRI<1 000)外,其他3 个季节均为该海域的优势种,这说明该调查海域是安氏白虾的主要生息区域。
3.2 生物多样性分析
虾蟹类的生物多样性分析结果呈现出春季的丰富度指数(D)、均匀度指数(J')、生物多样性指数(H')均高于其他季节。一方面由于春季伴随降雨量的增多,大陆入海径流量增加,海域营养盐逐渐增多,导致调查海域浮游生物大量繁殖生长,同时,随着水温上升,虾蟹类性腺发育成熟,很多种类要开展生殖洄游;另一方面,可能是调查海域为各种虾蟹类繁衍生息的区域,不同种类的虾蟹类洄游或移动到该海域进行繁殖生长,使得春季生物多样性指数均处较高水平。
生物多样性指数(H')季节变化为春>冬>夏>秋,而虾蟹类种类数季节变化为冬>春>秋>夏,两者之间并非同步变化。这与徐开达等[25]对中街山列岛甲壳类研究结果有所区别,具体原因尚有待今后进一步研究。调查海域的虾蟹类生物多样性指数H'平面分布存在较为明显的区域特征,冬季H'值分布呈现两头高、中间低,除冬季外,其它季节H'值主要集中在中部海域附近。T 检验分析得出仅夏季不同水深带生物多样性指数H'存在显著性差异(F=4.83,P=0.02<0.05),这很可能是因为夏季受台湾暖流影响,长江口径流量增大[26],不同水深的温度及盐度变化波动较大,随之水深带越深生物多样性指数越高,这与刘惠等[11]在舟山沿岸渔场海域甲壳类生物多样性随水深增加而增加的结论一致。
3.3 群落结构特征分析
通常从多个特征指标分析群落的稳定性,也能体现出群落内各种群之间保持长期联系的适应能力[27],根据等级聚类、非参数多维标度排序分析结果看,调查海域虾蟹类群落结构一年四季变化明显。春夏季划分为2 个不同组群,秋冬季划分为3 个不同组群。ANOSIM 检验表明春季R=0.889,P=0.005(P<0.01);夏季R=0.88,P=0.005(P<0.01);秋季R=0.916,P=0.007(P<0.01);冬季R=0.867,P=0.001(P<0.01)。检验结果得出不同季节的群组结构之间差异明显,且分组具有一定意义,造成夏季虾蟹类群落I-II 组群差异性较大的主要贡献种为三疣梭子蟹,I 组群主要分布调查海域东侧,II 组群分布在调查海域中部及西北侧。可能是三疣梭子蟹一般在夏季会更靠近河口及岛礁近岸附近的水域进行繁殖[28],伏季休渔期间近岸海域会进行三疣梭子蟹增殖放流活动[29],同时受人类捕捞活动的影响较少,当年生的幼蟹在该海域索饵生长。秋冬季种群之间差异性也较大,主要贡献种为安氏白虾、中国毛虾、三疣梭子蟹,其中虾类远高于蟹类贡献率,可能是10 月后气温随之降低,台湾暖流及长江口冲淡水削弱,外海高温盐水推进等影响三疣梭子蟹往更深处的海域进行越冬洄游[30]。安氏白虾及中国毛虾受水温下降影响从水深更浅的杭州湾河口区移动到近岸附近海域越冬[23],因此虾蟹类群落结构差异性较为显著。
4 结论
舟山岛北部近岸海域虾蟹类生物多样性指数(H')具有极显著的季节性差异(P<0.01);水深是影响该海域虾蟹类生物多样性指数(H')季节性变化的重要因素:水深<20 m 生物多样性(H')差异呈显著性(P<0.05),水深在20~40 m 差异呈不显著性(P>0.05),水深>40 m 差异呈极显著性(P<0.01);调查海域的虾蟹类群落结构可划分为2~3 个组群,以生态类型以广温低盐型和广温广盐型为主。
