利用红外相机技术对贺兰山岩羊集群行为的研究
2022-02-24丁乙轩李宗智孟德怀刘振生滕丽微
丁乙轩 刘 鹏 李宗智 孟德怀 刘振生,4 滕丽微,4*
(1.东北林业大学野生动物与自然保护地学院,哈尔滨,150040;2.成都大熊猫繁育研究基地,四川省濒危野生动物保护生物学重点实验室,成都,610086;3.四川省大熊猫科学研究院,成都,610086;4.国家林业和草原局野生动物保护学重点实验室,哈尔滨,150040)
集群是野生动物对特定时间与特定环境的适应行为[1],是有蹄类动物重要习性特征之一,也是有蹄类生态学研究中的重要内容[2-3]。综合被捕食风险、栖息地特征、食物丰富度、生存繁殖需求等多方面因素可知,有蹄类动物选择不同的集群行为使其更易发现并逃避捕食者,获得配偶和更多资源,提高采食效率,有利于其生存繁殖[4-6]。然而,当野生动物集群规模过大时,个体间资源的竞争加剧,疾病传播概率也会增加[7-9]。集群类型与集群规模是有蹄类集群特征最直接的反映,因此,了解有蹄类集群类型与规模及其变化规律有助于了解其种群特征及动态变化[10],为野生有蹄类动物的科学管护提供理论依据[11]。
岩羊(Pseudoisnayaur)为偶蹄目(Artiodactyla)牛科(Bovidae)动物[12],是国家二级保护野生动物[13],生态价值及经济价值尤为显著,主要分布在我国青藏高原及其周边地域[14-15]。岩羊善于在岩石上奔跑跳跃,常以高山岩石、悬崖为栖息地营群居性生活,具有显著的集群特征[16]。目前,对于岩羊集群行为的研究多采用直接观察、GPS跟踪等方法,相比之下,红外相机技术能昼夜连续工作,人力成本低,不会对动物造成损伤,人为干扰较小,受环境限制小[17],十分适合岩羊集群行为的野外调查研究。
本研究利用红外相机于2014年10月开始对贺兰山国家级自然保护区岩羊集群行为进行为期1年的监测,对红外相机监测数据进行处理,分析岩羊集群行为特征及冷暖季间变化,了解岩羊对贺兰山生存环境的适应性,为贺兰山岩羊种群的保护管理提供科学依据。
1 研究区概况
贺兰山国家级自然保护区地处宁夏平原和内蒙古阿拉善高原交界位置(38°21′—39°22′N,105°49′—106°42′E)[18]。贺兰山呈西南—东北的山势走向,南北长约250 km,东西宽20~40 km,海拔2 000~3 000 m[19]。贺兰山地处内陆腹地,具有典型的大陆性气候,冬长夏短,春寒秋凉。贺兰山年平均气温-0.9 ℃,年较差26.0 ℃[5],年均降水量418.1 mm,全年62%的降水量集中在6—9月,期间平均降水量为260.2 mm,全年日照时间为3 022.8 h[18,20]。该地区森林类型为典型温带山地森林,植被具有显著的垂直分布特点,可划分为4部分:海拔1 400~1 600 m为山地草原带,1 600~2 000 m为山地疏林草原带,1 900~3 100 m为山地针叶林带,3 100~3 556 m为亚高山灌丛和草甸带[18,21]。
2 研究方法
2.1 红外相机布设
2014年10月—2015年10月,利用红外相机(猎科Ltl Acorn-6210MC)对贺兰山国家级自然保护区岩羊行为活动进行监测。相机布设在兽道、水域或具有岩羊活动痕迹处,距地面0.5~0.8 m,固定在树干或人为设置的固定物上。利用地理信息系统在贺兰山地区叠加1 km×1 km的公里网格,每个网格架设1台红外相机,相机间距不低于300 m。相机镜头基本与地面平行,避免阳光直射。相机设置24 h连续工作,每拍摄2张照片后录制15 s的视频,灵敏度调节为中档。相机布设完成后,对相机编号、GPS位点信息(纬度、经度、海拔)、其他环境信息(如动物痕迹、周围植物种类等)、日期、人员相关信息等内容进行记录。定期收集照片,更换电池和储存卡,以确保相机能持续工作,相机布设位点见图1。
2.2 数据处理与分析
以研究区域气候特征、岩羊不同季节食物资源差异及其生理周期为依据,将研究时段划分为冷季(10月11日—次年4月10日)和暖季(4月11日—10月10日)。仔细查验红外相机监测到的岩羊照片,根据岩羊角的形状及大小、体型大小、毛色等区分岩羊的雌雄及成幼,并以此来划分岩羊集群类型与集群规模。
岩羊雌雄以及成幼划分标准如下:(1)成年雄性:体型较大,角长且粗大(40~80 cm),两角基仅具一狭窄缝隙,末端明显向后外侧弯曲生长。(2)成年雌性:与成年雄性个体相比体型略小,角基本无弯曲且短(仅10 cm左右),多向上生长,基部扁平且逐渐变细。(3)亚成体:体型明显小于成年个体,体长约为成年个体的2/3,角短且小,仅根据照片不能区分雌雄。(4)幼体:体型很小,几乎没有角(仅少数个体头顶有角的突出部分),需要雌性岩羊哺乳喂养,仅根据照片不能区分雌雄。
依据红外相机监测到的岩羊照片与视频数据,将岩羊集群行为划分为6种集群类型,即:(1)单个雄性,在研究区域内,被监测到单个雄性个体单独活动的情况。(2)单个雌性,在研究区域内,被监测到单个雌性个体单独活动的情况。(3)母仔群,在研究区域内,被监测到成年雌性个体带着幼仔活动的情况。(4)雄性混合群,在研究区域内,被监测到2只及以上成年雄性个体在同一地点活动的情况。(5)雌性混合群,在研究区域内,被监测到2只及以上成年雌性个体在同一地点活动的情况(不带幼仔)。(6)雌雄混合群,在研究区域内,被监测到至少有1只成年雄性个体和1只成年雌性个体在同一地点活动的情况。
在本研究中,对不同集群类型出现的频次差异、不同集群类型出现频次的季节差异采用卡方检验进行分析,采用K个独立样本的Kruskal-WallisH检验分析不同集群大小的季节差异、不同集群类型群平均大小的季节差异、不同集群类型群大小的季节差异。使用SPSS 20.0和Excel 2019处理分析数据。
3 结果与分析
3.1 集群类型的频次分布
2014年10月—2015年10月,在贺兰山国家级自然保护区共布设红外相机120台(图1),对红外相机监测照片进行整理,共识别出可供分析的岩羊照片2 771群次,经统计,总计出现岩羊4 942只次,通过计算分析可得岩羊的平均集群大小是(1.78±1.49)只,最大集群内有岩羊20只,最小集群为1只。单个雄性出现频次最高,总计996次(35.94%);单个雌性出现675次(24.36%);母仔群322群(11.62%),790只次;雄性混合群243群(8.77%),703只次;雌性混合群129群(4.66%),323只次;雌雄混合群406群(14.65%),共计1 455只次。不同集群类型出现频次间差异极显著(χ2=1 108.87,df=5,P<0.001)。

图1 红外相机在研究区域内布设位点
3.2 集群类型的季节性动态
卡方检验结果表明,集群类型在冷暖季间存在极显著差异(χ2=188.67,df=5,P<0.001)(表1)。各类型集群中,在冷季和暖季出现频次最高的均为单个雄性,单个雄性、单个雌性、雄性混合群和雌性混合群在暖季出现的频次较高,而母仔群和雌雄混合群更多出现在冷季。
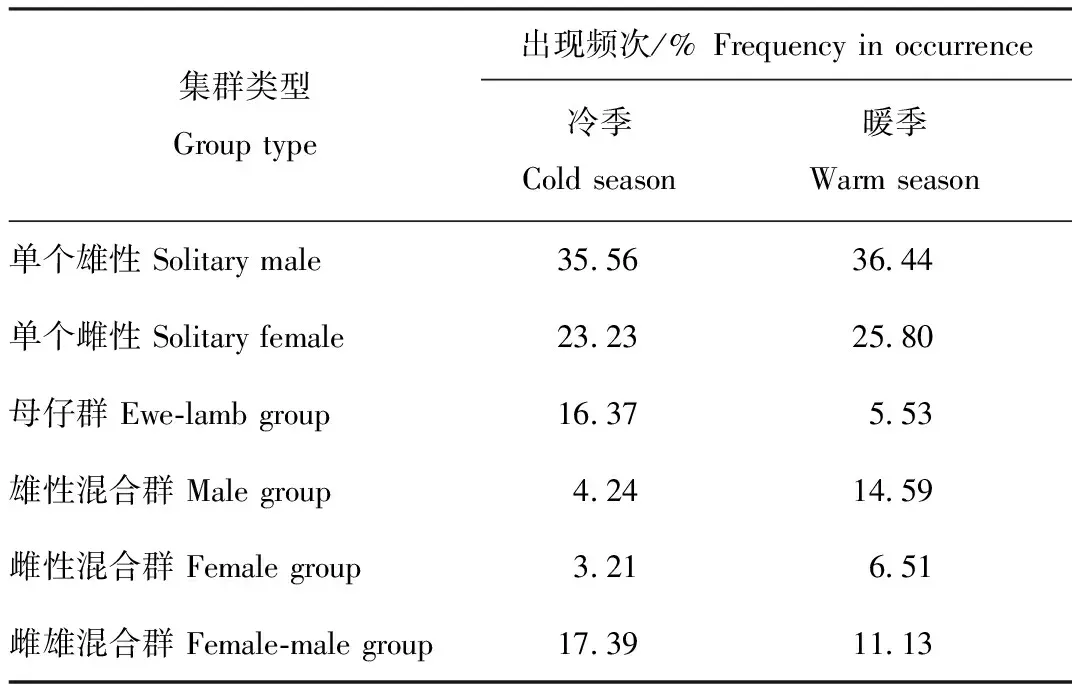
表1 贺兰山岩羊不同集群类型频次的季节性差异
3.3 集群大小的季节性差异
经统计分析,共有岩羊1 558群次被发现于冷季,暖季共发现岩羊1 213群次。通过计算分析可知,冷季集群规模为1~17只,平均集群大小为1.78只;暖季集群规模为1~20只,平均集群大小为1.79只。无论是冷季还是暖季,岩羊均多以单个个体集群类型出现,其出现频次均占各季节总集群数的60%左右(图2)。Kruskal-WallisH检验结果表明,集群大小在冷暖季间差异不显著(χ2=2.29,df=1,P=0.131)。
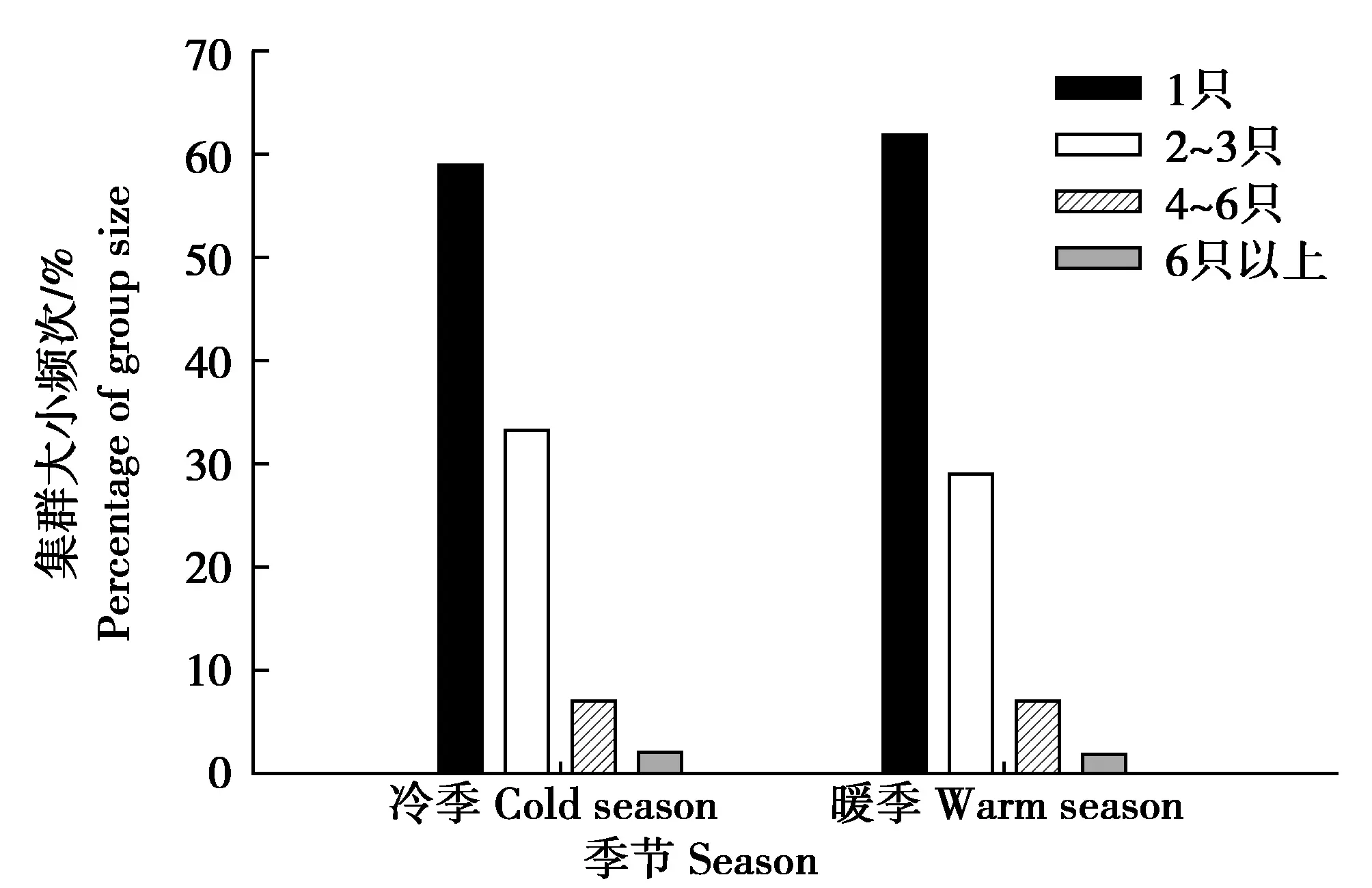
图2 贺兰山岩羊不同集群大小频次的季节性差异
3.3.1 不同集群类型群平均大小的季节性差异
在冷暖两季中,雌雄混合群的群平均大小分别为(3.54±2.14)、(3.67±2.39)只;母仔群群平均大小分别为(2.42±0.92)、(2.55±0.88)只;雄性混合群群平均大小分别为(2.29±0.76)、(3.12±2.14)只;雌性混合群群平均大小分别为(2.54±1.74)、(2.48±0.96)只。Kruskal-WallisH检验结果表明,不同集群类型群平均大小在冷暖季间无显著差异(χ2=0.428,df=1,P=0.513)。
3.3.2 不同集群类型群大小的季节性差异
母仔群无论在冷季还是暖季,其集群大小均以2~3只为主,部分为4~6只群,几乎没有6只以上群,冷暖季无显著差异(χ2=1.434,df=1,P=0.231);在2个季节中,雄性混合群均以2~3只群为主,暖季4~6只群多于冷季,暖季还存在小部分6只以上的群,而冷季则无,冷暖季差异不显著(χ2=0.185,df=1,P=0.667);雌性混合群在冷季和暖季同样以2~3只群为主,冷季4~6只群及6只以上群均多于暖季,冷暖季差异不显著(χ2=0.449,df=1,P=0.513);2~3只群的雌雄混合群在冷暖季出现频次最高,还有部分雌雄混合群在2个季节为4~6只群,6只以上群在冷暖季出现频次最低,冷暖季差异不显著(χ2=0.200,df=1,P=0.655)(表2)。
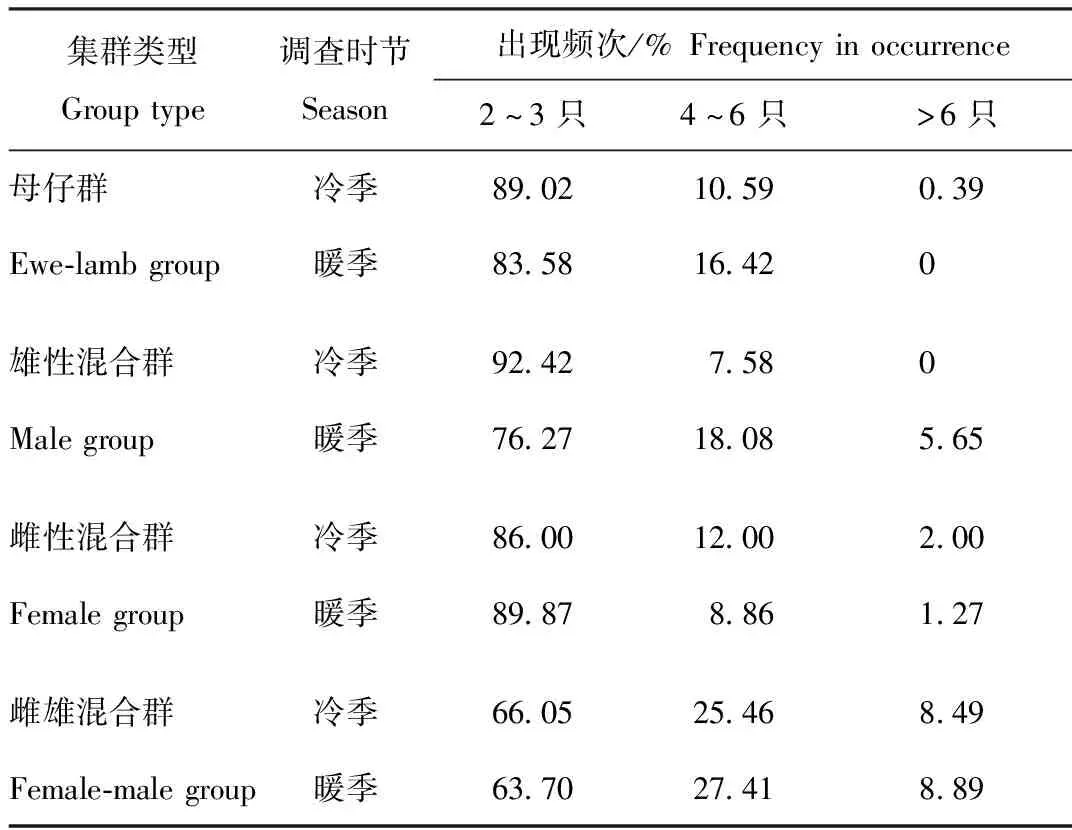
表2 贺兰山岩羊不同集群类型群大小频次的季节性差异
4 讨论
岩羊集群规模与被捕食风险、栖息地特征、食物丰富度等密切相关[3,22-25]。本研究发现,岩羊在冷暖季集群大小无显著差异,均以1~3只群为主,呈集小群特征,这与曹丽荣等[23]对贺兰山岩羊春冬两季集群特征的研究结果相似。岩羊的天敌,如狼(Canislupus)、猞猁(Lynxlynx)等,自20世纪80年代起在贺兰山已基本灭绝,虽然赤狐(Vulpesvulpes)、秃鹫(Aegypiusmonachus)等对岩羊仍能构成一定威胁[6],但这不足以让岩羊扩大集群规模来降低被捕食风险。栖息地特征对集群规模有重要影响,与生存在封闭环境中的偶蹄类动物相比,生存在开阔环境中的偶蹄类动物更易形成大规模集群[22],贺兰山地势陡峭险峻,故此处岩羊更喜欢集小群。贺兰山植被类型简单且覆盖率低[23],可供岩羊采食的植物较少,集小群可以有效减少种群内个体之间对食物资源的竞争,因此不同集群类型群平均大小在冷暖季间无显著差异。
本研究结果表明,贺兰山岩羊平均集群大小为(1.78±1.49)只,小于本课题组2004—2005年样线调查结果(4.86±2.54)只[6],本研究还发现不同集群类型群大小在冷暖季间无显著差异,这也与过去不同,这可能与研究地生境类型有关。本研究是在贺兰山森林生境中开展的,而过去则是在贺兰山全境内进行的,包括荒漠、草原等生境类型,为适应不同生境类型,岩羊的集群行为也会发生改变。此外,过去的研究中将季节划分成四季,而本次研究按冷暖季划分,这可能造成检验结果差异重合。
本研究利用红外相机拍摄贺兰山岩羊2 771群次,共计4 942只次。6种类型集群出现频次差异显著,经统计,其中出现频次最高的为单个雄性,单个雌性、雌雄混合群、母仔群、雄性混合群以及雌性混合群出现频次依次降低。这是因为雄性岩羊体型较大,被捕食风险较低,更易获得可利用的食物资源[3,24];且雄性个体多具有较强的领域意识[24],个体间竞争较大,不喜其他雄性进入领地,故雄性岩羊多为单独活动。此外,该研究区域可能雄性数量较多。
不同集群类型在冷暖季间存在显著差异,母仔群和雌雄混合群更多出现在冷季,而其他类型集群则在暖季出现频次较高。每年11—12月为岩羊发情交配期[19],随着季节由暖季转入冷季,成年的雄性个体和雌性个体会逐渐脱离原始群体,加入或重新组成雌雄混合群[3,26,27],这样的季节性繁殖导致了雌雄混合群在冷季出现频次更多。相对于雄性个体,雌性和幼仔面临更大的捕食风险,相关研究表明,在选择生境时,雌性有蹄类动物更注重安全性,营养需求反而是其次[5,28-30]。岩羊产仔期处于暖季,为保证幼仔的存活,雌性岩羊会选择隐蔽性较好的生境(即便该区域食物缺乏),以此降低幼仔被捕食的风险,这也是母仔群在暖季出现频次较低的原因。从暖季到冷季,贺兰山地区的食物质量和丰富度明显下降[6,22],营养压力让岩羊不得不增加活动时间,面对更大的被捕食风险去获得高质量食物,以满足自身能量需求和维持机体平衡的需要并弥补季节性营养不足,因此母仔群在冷季出现频次增多。
虽然红外相机技术在野外调查中具有很多优势,但其仍存在一定局限性。红外相机只能拍摄相机前一定扇形范围内的动物情况(感应半径),当个体活动区域超过红外相机拍摄范围,就会造成原始数据的缺失[31]。此外,本研究是在森林生境中开展的,遮蔽物相对较多,也会对拍摄及数据收集造成一定影响。在今后的研究过程中,可以采用更先进的红外相机,对贺兰山岩羊进行全方位监测[32],进一步提高数据采集的完整性与准确性。
