小麦田看麦娘对甲基二磺隆的抗性水平及分子机制初探
2022-02-22王玛丽杨佳佳操海群
王玛丽, 李 奇, 杨佳佳, 廖 敏, 操海群, 赵 宁
(作物有害生物综合治理安徽省重点实验室,安徽农业大学 植物保护学院,合肥 230036)
看麦娘Alopecurus aequalis是我国农田常见恶性杂草之一,主要分布在黄河及长江中下游流域等地区稻茬麦田及油菜田,其整株分蘖旺盛、群体竞争能力强,可导致作物严重减产甚至绝收[1],目前主要使用甲基二磺隆、精噁唑禾草灵、异丙隆等化学除草剂对其进行防除。甲基二磺隆属于乙酰乳酸合成酶 (ALS) 抑制剂中的磺酰脲类除草剂,其作用靶标为ALS。ALS 能够催化植物体内支链氨基酸合成反应的第一步,是多种不同化学类别除草剂的共同靶标[2]。由于ALS 抑制剂类除草剂杀草谱广、安全、高效且毒性较低,在世界范围内得到了广泛的应用,但由于其作用位点单一,长期重复使用极易导致杂草产生抗药性[3]。据统计,截至2021年5 月,全球已有167 种杂草生物型对ALS 抑制剂类除草剂产生了抗性,其中17 种来自于中国[4]。
杂草对除草剂的抗性机制主要包括两个方面:一是靶标基因突变或过量表达导致的靶标抗性,二是杂草对除草剂的隔离、传导能力改变或者代谢能力增强导致的非靶标抗性[5-6]。据报道,ALS基因5 个保守区第122 位、第197 位、第205 位、第376 位、第377 位、第574 位、第653位和第654 位8 个氨基酸位点发生突变是杂草对ALS 抑制剂类除草剂产生抗性的主要原因之一[7-8]。同时,关于杂草对ALS 抑制剂代谢能力增强所致抗性的报道也逐渐增多[9]。其中,细胞色素P450s、谷胱甘肽S-转移酶 (GST) 、糖基转移酶(UDP-GT) 等活性的增强是杂草对除草剂产生代谢抗性的重要原因[10]。
近年来,我国江苏、安徽、山东等省部分地区农户普遍反映,多种常用除草剂在大田推荐剂量下已无法对看麦娘等禾本科杂草进行有效防控,即看麦娘可能产生了抗药性。2018 年,Guo等[11]对黄河及长江中下游流域采集的47 个看麦娘种群进行抗药性检测,发现多个种群已对甲基二磺隆产生高水平抗性,而ALS基因第197 位和第574 位氨基酸位点突变是导致抗性产生的主要原因之一。关于抗药性看麦娘的防治目前已成为我国农业生产中亟待解决的问题。
2020 年5 月,本课题组在安徽省凤台县进行田间杂草抗药性调查过程中,采集到一个看麦娘疑似抗性种群AHFT-01,该种群所在地块使用甲基二磺隆已无法对其进行有效防治。为明确其抗药性发生情况及可能存在的抗性机制,本研究首先在整株水平上测定了AHFT-01 对甲基二磺隆的敏感性,并对抗性和敏感种群ALS基因进行了扩增、测序和比对,以明确AHFT-01 对甲基二磺隆的抗性水平及其抗性产生的靶标分子机制,同时测定了该种群对不同ALS 抑制剂的交互抗性,以及对乙酰辅酶A 羧化酶 (ACCase) 及光合系统II(PSII) 抑制剂类除草剂的多抗性,以期为小麦田抗性看麦娘的防治及减缓其抗药性发展提供理论依据。
1 材料与方法
1.1 供试材料
1.1.1 杂草种子 看麦娘Alopecurus aequalis疑似抗性种群AHFT-01 采集自安徽省淮南市凤台县现代农业科技示范园小麦田 (32.57°N,117.65°) ,该地块已连续多年使用甲基二磺隆进行杂草防除;敏感种群AHHN-02 为实验室保存株系,采集自安徽省淮南市大通区九龙岗镇曹店村非耕地 (32.63°N,117.13°E) ,该地块从未使用过除草剂。
1.1.2 药剂与试剂 30 g/L 甲基二磺隆 (mesosulfuronmethyl) 可分散油悬浮剂 (OD) 及69 g/L 精噁唑禾草灵 (fenoxaprop-P-ethyl) 水乳剂 (EW) ,拜耳作物科学 (中国) 有限公司;240 g/L 烯草酮 (clethodim)乳油 (EC) ,山东奥坤作物科学股份有限公司;10% 唑啉草酯 (pinoxaden) OD 及5% 氟唑磺隆(flucarbazone-sodium) OD,安徽尚禾沃达生物科技有限公司;5%咪唑乙烟酸 (imazethapyr) 水剂(AS) ,南通金陵农化有限公司;7.5%啶磺草胺(pyroxsulam) 水分散粒剂 (WG) ,科迪华农业科技有限责任公司;50%异丙隆 (isoproturon) 可湿性粉剂 (WP) ,安徽广信农化股份有限公司。DNA提取试剂盒,江苏康为世纪生物科技股份有限公司;DNA 回收试剂盒,南京诺唯赞生物科技股份有限公司。
1.1.3 主要仪器 T100 型梯度PCR 仪,美国Bio-Rad 公司;万分之一电子天平,上海越平科学仪器 (苏州) 制造有限公司;ERS 200 电泳仪,北京原平皓生物技术有限公司;GL21M 台式高速冷冻离心机,凯达科学仪器有限公司;BIC-400 人工气候箱,上海博迅实业有限公司医疗设备厂;3WP-2000 型行走式喷雾塔,农业部南京农业机械化研究所;ZF-288 凝胶成像分析系统,上海金鹏分析仪器有限公司。
1.2 试验方法
1.2.1 看麦娘对甲基二磺隆的抗性水平测定 采用温室盆栽法[12]在整株水平上进行测定。选取颗粒饱满的看麦娘AHFT-01 和AHHN-02 种群种子,分别置于直径9 cm、垫有一层滤纸的培养皿中,添加纯水保湿;置于光照培养箱中,于25 ℃/20 ℃、12 h/12 h 光周期条件下催芽。将土壤和有机质按1 : 1 质量比混匀,填入长 (宽) 7 cm、高7 cm的方形塑料盆钵内,压实后放入接水托盘。于托盘底部浇水,待塑料盆钵中土壤完全浸湿后播种。待看麦娘种子萌芽至1 cm 左右,选取10 株转移至填有壤土的塑料盆钵,置于光照培养箱中常规培养。待看麦娘生长至2 ~ 3 叶期时间苗,保留6 株长势一致的幼苗;至3 ~ 4 叶期时采用行走式喷雾塔进行茎叶喷雾处理,喷头距植株叶片约50 cm,喷液压力275 kPa,喷液量450 L/hm2。
根据预试验结果设置甲基二磺隆施用剂量,以喷施清水为空白对照 (表1) 。于施药后21 d 剪取看麦娘地上部分,称量并记录鲜重。每个处理重复3 次,每个重复1 盆,整个试验重复2 次。
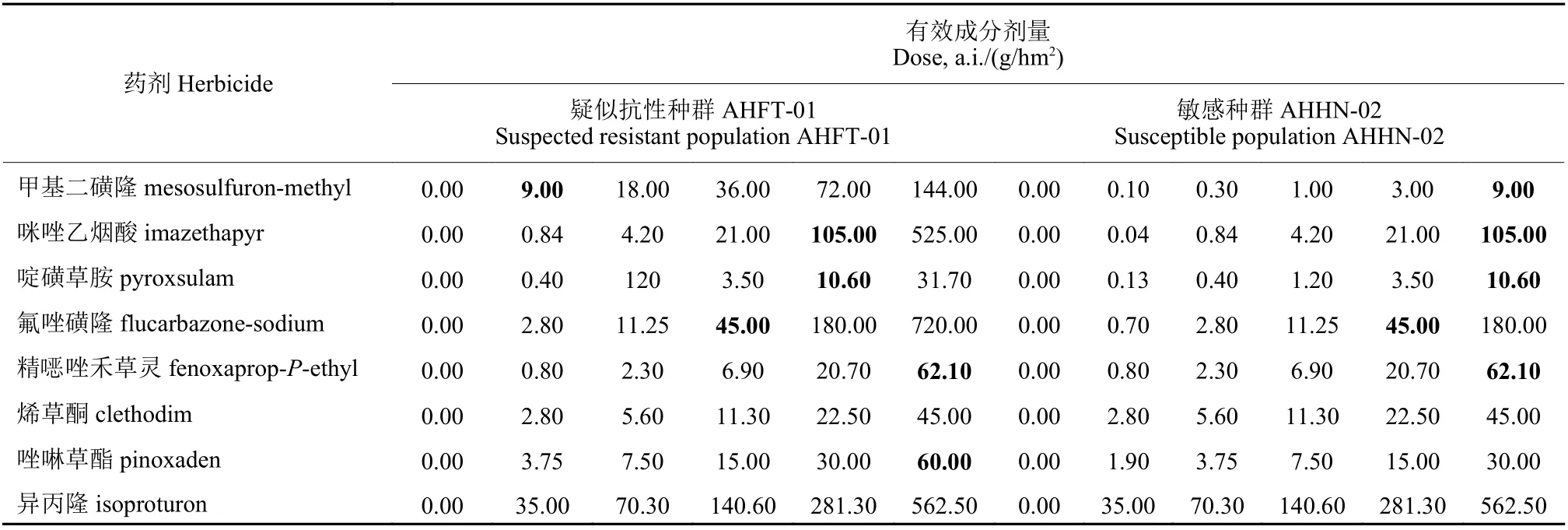
表1 看麦娘种群敏感性测定中各除草剂施用剂量Table 1 Herbicide doses used for determining the susceptibilities of Alopecurus aequalis populations
1.2.2 看麦娘抗性、敏感种群ALS基因序列扩增
DNA 提取:待看麦娘幼苗培养至3 ~ 4 叶期时,从AHFT-01 和AHHN-02 种群中随机选取10 个单株,每株剪取约50 mg 幼嫩叶片组织,根据植物DNA 提取试剂盒使用说明提取其基因组DNA,于 −20℃冰箱保存,备用。
引物合成:参考Iwakami 等[13]报道的引物序列 (表2) ,由北京擎科生物科技有限公司进行引物合成。所用引物对能够扩增看麦娘ALS基因不同拷贝全长序列,且包含已报道的8 个ALS基因氨基酸突变位点。
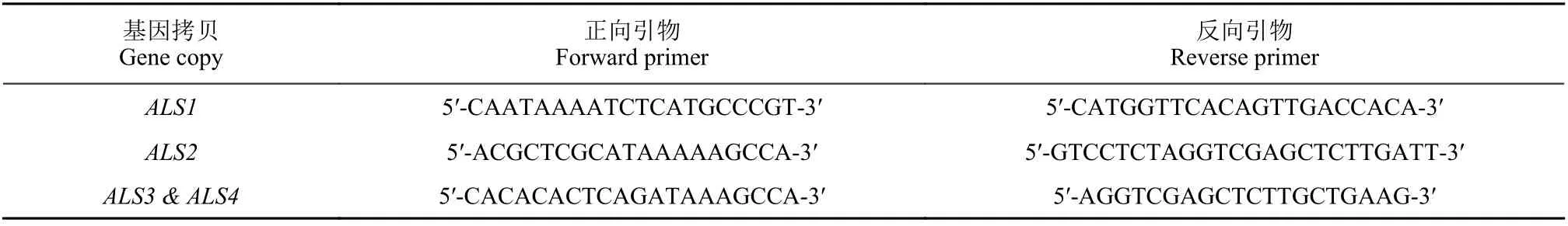
表2 ALS 基因全长序列扩增引物Table 2 Primers used for amplifying the full-length ALS genes
PCR 扩增体系为50 μL,分别包含:2 × Es Taq MasterMix 25 μL、ddH2O 20 μL、10 μmol/L 正向引物2 μL、10 μmol/L 反向引物2 μL 和基因组DNA 1 μL。PCR 反应条件为:98 ℃预变性3 min;98 ℃变性30 s,58 ~ 63 ℃退火30 s,72 ℃延伸30 s,共33 个循环;最后于72℃终延伸10 min。
1.2.3ALS基因序列差异分析 将PCR 扩增产物于1%的琼脂糖凝胶中进行电泳检测,切取目的片段,使用DNA 回收试剂盒对目的基因进行回收纯化,然后送擎科生物科技有限公司测序,测序引物为PCR 扩增引物。看麦娘ALS 氨基酸序列编号以模式植物拟南芥Arabidopsis thalianaALS 全长序列为标准,使用DNAMAN 6.0.3 软件分别对疑似抗性种群AHFT-01 和敏感种群AHHN-02 的ALS基因序列进行比对,分析其ALS基因序列差异。
1.2.4 看麦娘种群对不同除草剂的敏感性测定
按照1.2.1 节所述方法,分别测定看麦娘疑似抗性种群AHFT-01 和敏感种群AHHN-02 对ALS 抑制剂类除草剂咪唑乙烟酸、啶磺草胺、氟唑磺隆,ACCase 抑制剂类除草剂精噁唑禾草灵、烯草酮、唑啉草酯,以及PSII 抑制剂类除草剂异丙隆的敏感性水平,各除草剂具体施药剂量见表1。
1.2.5 数据处理 药剂处理后21 d,剪取看麦娘地上部分并称量鲜重,按照公式 (1) 计算相对鲜重。

式中:y为相对鲜重;mt为处理组杂草平均鲜重,g;m0为空白对照组杂草平均鲜重,g。
采用SPSS v.19.0 软件的一般线性模型对重复试验数据进行组间方差分析,结果显示两组数据间无显著差异 (P> 0.05) 。将重复试验数据合并,采用SigmaPlot v.12.5 软件四参数双逻辑非线性回归模型 (2) 进行数据处理,绘制剂量响应曲线,并计算抑制杂草生长50%所需的除草剂剂量(GR50,g/hm2) 。

式中:yx为特定除草剂用量x下杂草的相对鲜重,%;C为剂量反应下限;D为剂量反应上限;b为斜率。
按照公式 (3) 计算种群的抗性倍数 (resistance index, RI) 。

式中:GR50(R)为抗性种群杂草GR50值,g/hm2;GR50(S)为敏感种群杂草GR50值,g/hm2。
抗性水平分级标准[14]:RI<2,敏感;2≤RI<5,低抗;5≤RI<10,中抗;RI≥10,高抗。
2 结果与分析
2.1 看麦娘种群对甲基二磺隆的抗性水平
采用甲基二磺隆按大田推荐剂量处理后,AHFT-01 种群所有植株均可正常生长,而AHHN-02 种群植株均枯萎死亡,表明AHFT-01 种群已对甲基二磺隆产生了抗性。根据整株剂量响应曲线可得,甲基二磺隆对AHFT-01 和AHHN-02 种群的GR50值分别为有效成分44.74 和1.57 g/hm2,表明AHFT-01 对甲基二磺隆产生了高水平抗性,其抗性倍数为28.50 (表3) 。

表3 看麦娘种群对甲基二磺隆的抗性水平Table 3 Resistance index to mesosulfuron-methyl of different Alopecurus aequalis populations
2.2 看麦娘抗性和敏感种群ALS 基因序列差异
对看麦娘不同种群ALS基因进行扩增,共得到ALS1、ALS2、ALS3和ALS44 个基因拷贝。其中,ALS1和ALS2在所有植株DNA 中均得到了扩增产物,而ALS3和ALS4仅在部分植株DNA 中可扩增到,且ALS3和ALS4存在单碱基缺失导致的移码错位,氨基酸翻译异常,被鉴定为假基因[13]。因此本研究仅分析不同种群ALS1和ALS2的序列差异。
本研究中,ALS1和ALS2序列长度均为1917 bp,共编码639 个氨基酸,包含已报道的8 个能够导致杂草对除草剂产生抗性的氨基酸位点。序列比对结果显示:AHHN-02 种群ALS不同拷贝已知位点均未发生突变;与AHHN-02 相比,AHFT-01ALS1第197 位氨基酸由脯氨酸Pro (CCC) 突变为苏氨酸Thr (ACC) ,且测定的10 个单株ALS1均发生了纯合突变,突变频率为100%,其余7 个位点及ALS28 个位点则均未发生任何突变 (表4) 。
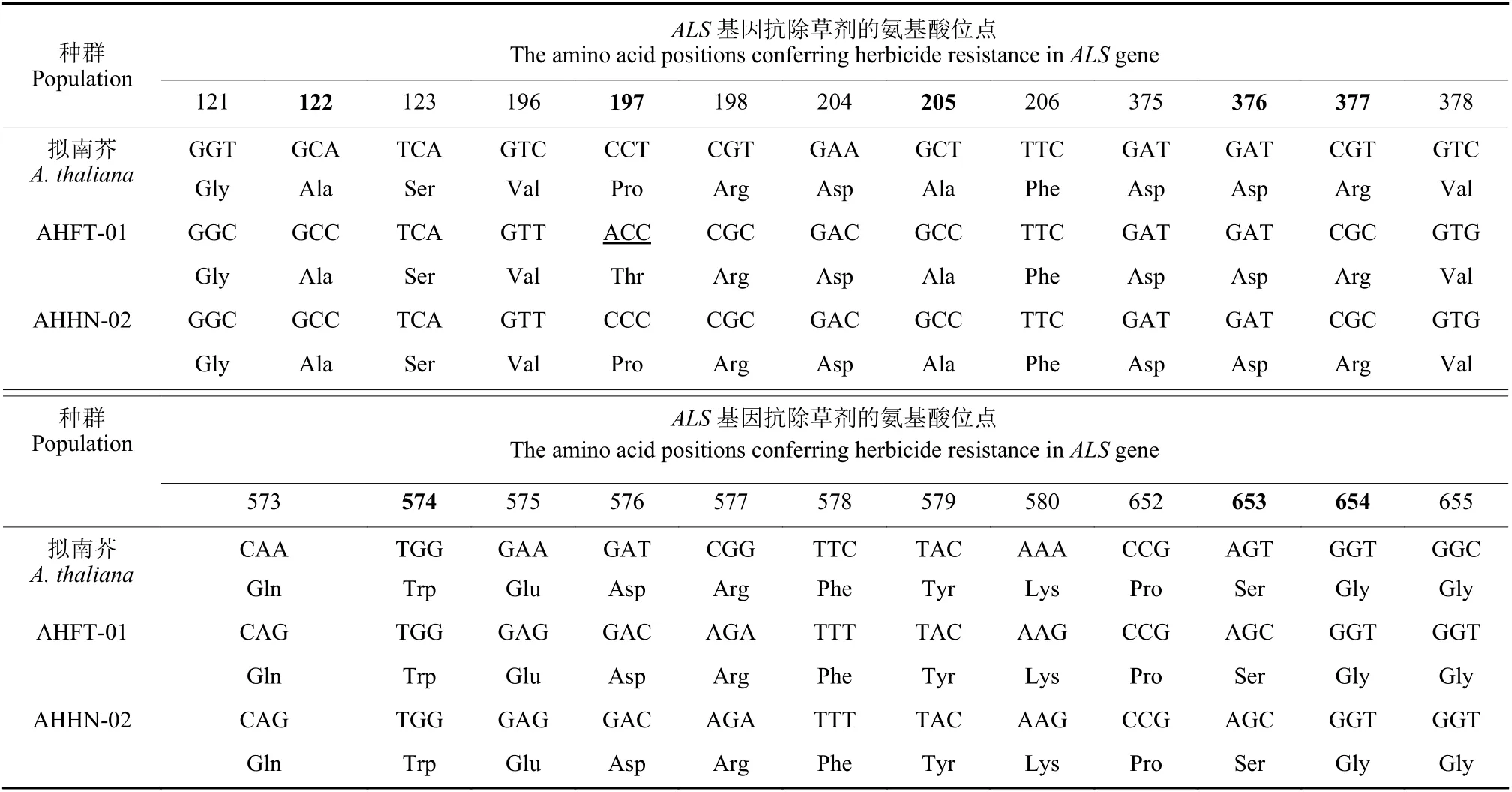
表4 不同看麦娘种群ALS1 基因部分序列Table 4 Partial sequences of ALS1 genes from different Alopecurus aequalis populations
2.3 看麦娘抗性种群对其他不同除草剂的敏感性
看麦娘AHFT-01 和AHHN-02 种群对不同除草剂的剂量响应曲线如图1 所示。正如预期,敏感种群AHHN-02 在所有测试药剂大田推荐剂量处理下均发生了死亡,与之相比,抗性种群AHFT-01对ALS 抑制剂类的啶磺草胺及氟唑磺隆产生了高水平抗性,抗性倍数分别为10.15 和10.13;对咪唑乙烟酸产生了中等水平抗性,抗性倍数为8.18(表5) 。
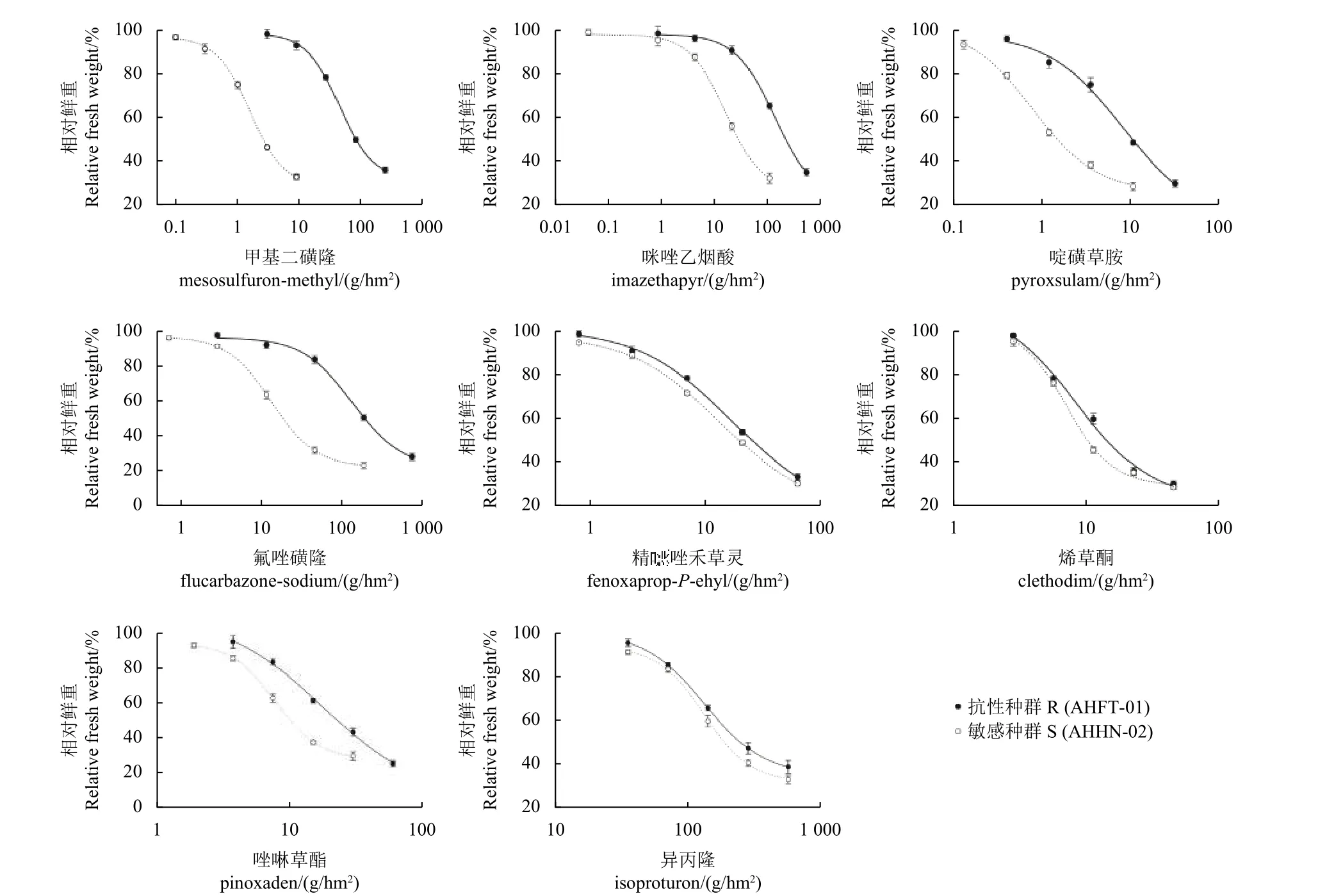
图1 看麦娘不同种群对8 种除草剂的敏感性水平Fig. 1 The susceptibility levels of different Alopecurus aequalis populations to eight herbicides
同时,本研究还测定了不同种群看麦娘对ACCase 和PSII 抑制剂类除草剂的多抗性,相比AHHN-02,AHFT-01 种群对唑啉草酯同时产生了低水平抗性 (RI = 2.27) ,对精噁唑禾草灵、烯草酮及异丙隆则依旧较为敏感 (RI<2) (表5) 。

表5 看麦娘不同种群对其他除草剂的敏感性水平Table 5 The susceptibility levels of different Alopecurus aequalis populations to other herbicides
3 讨论与结论
ALS 抑制剂是目前世界上应用最为广泛的除草剂种类之一,其除草活性高、安全性好,但是长期大量重复使用容易导致杂草抗药性的快速发生和蔓延[15]。有研究表明,ALS 抑制剂类除草剂使用不到10 次就会导致杂草对其产生抗性[14]。截至目前,全世界范围内已有167 种杂草生物型对ALS 抑制剂类除草剂产生了抗性,例如澳大利亚的硬直黑麦草Lolium rigidum[16]和野荠菜Raphanus raphanistrum[17]、加拿大的地肤Kochia scoparia[18]和长芒苋Amaranthus tuberculatus[19]、中国的日本看麦娘Alopecurus japonicus[20]等。本研究中,从安徽省凤台县采集的看麦娘种群AHFT-01 也已经对甲基二磺隆产生高水平抗性 (RI > 10) ,而根据记录,该种群所在地块已连续多年使用ALS 抑制剂类除草剂进行杂草防除。这再一次印证了单一作用机制除草剂长期大量重复使用是导致杂草产生抗药性的重要原因之一。
据报道,大部分ALS 抑制剂类除草剂抗性案例是由于ALS 发生氨基酸突变引起的,氨基酸突变导致靶标酶构象改变,进而阻碍除草剂与靶标的结合[21]。迄今为止,已经在多种杂草中发现ALS基因8 个位点可以发生多种氨基酸突变形式。例如,ALS基因Ala-122-Tyr 突变是澳大利亚野荠菜种群对氯磺隆、磺草唑胺、甲氧咪草烟产生抗性的主要原因[22];Pro-197-Thr 和Trp-574-Leu两种ALS基因突变是我国山东省播娘蒿Descurainia sophia种群对ALS 抑制剂类除草剂产生抗性的重要原因之一[23];Trp-574-Leu 和Ala-205-Val 两种ALS基因突变导致了黑龙江省反枝苋Amaranthus retroflexus种群对噻吩磺隆和氟磺胺草醚的抗性[24]。第197 位和第574 位是ALS基因最常见的突变位点[15],其中第197 位已发现至少13 种氨基酸突变形式,包括Pro-197-Arg、Pro-197-Ser 和Pro-197-Thr 等[25-26]。Bi 等[27]报道,ALS基因Pro-197-Thr 突变是日本看麦娘对甲基二磺隆产生抗性的原因;Zhao 等[28]报道,ALS1基因Pro-197-Tyr 突变是看麦娘对甲基二磺隆产生抗性的重要靶标分子机制。本研究中,看麦娘种群AHFT-01 的ALS1基因发生了Pro-197-Thr 纯合突变,且突变发生频率高达100%,表明ALS1基因第197 位突变也是AHFT-01 对甲基二磺隆产生高水平抗性的重要原因之一。
不同突变位点或同一位点的不同突变形式,会导致杂草产生不同的交互抗性谱[29-30]。据报道,ALS基因第197 位氨基酸突变通常会导致对磺酰脲类除草剂的抗性[31]。Xia 等[2]报道,看麦娘种群JTJY-1 对磺酰脲类和三唑并嘧啶类除草剂具有中等或较高水平交互抗性,对咪唑啉酮类抗性水平较低;而看麦娘种群JHHZ-1 则对上述3 种类别的ALS 抑制剂均具有抗性;两个种群的ALS基因分别检测出了Pro-197-Thr 和Trp-574-Leu 突变,但是该研究并未表明突变具体发生在ALS的哪个拷贝上。本研究中,ALS1基因发生Pro-197-Thr突变的看麦娘种群AHFT-01 对3 种不同类别的ALS 抑制剂类除草剂均产生了高水平或中等水平的交互抗性,这与前人的报道结果相似[2],但是也存在一定差异,这可能是由于种群用药历史背景存在差异等原因所致[29]。
本研究鉴定得到的看麦娘种群AHFT-01 对常用ALS 抑制剂类除草剂均产生了抗药性,已严重威胁到小麦的安全生产,因此,本研究同时测定了该种群对ACCase 抑制剂类和PSII 抑制剂类除草剂的敏感性,以期发现可用于该抗性种群看麦娘防治的药剂。结果表明,AHFT-01 对ACCase抑制剂唑啉草酯也产生了低水平抗性,但是对其他ACCase 抑制剂如精噁唑禾草灵、烯草酮以及PSII 抑制剂异丙隆依旧敏感,因此在实际生产中,仍可采用精噁唑禾草灵和异丙隆继续对该小麦田抗性看麦娘进行有效防控。此外,将化学药剂防治与轮作、深翻耕等农业措施相结合,是延缓看麦娘等农田恶性杂草产生抗药性的有效措施之一。
