外源硼对干旱胁迫下厚皮甜瓜幼苗生长和 抗氧化系统的影响
2022-02-21李艳超张彥萍刘海河武雅娴
李艳超,张彥萍,刘海河,刘 月,封 科,武雅娴
(河北农业大学 园艺学院/河北农业大学科教兴农中心,河北 保定 071001)
厚皮甜瓜在植物学分类上属于葫芦科中甜瓜属的一年蔓生草本植物,起源于非洲埃塞俄比亚高原及其毗邻地区,我国是甜瓜的栽培次级起源中心之一,栽培历史悠久,目前已经成为世界上最大的设施甜瓜栽培生产国[1]。硼是植物必需的微量元素之一[2],是植物生长发育必须的营养元素,对植物的光合系统、产量、品质均有影响[3]。硼是细胞壁必不可少的组成成分,对于植物的生长发育有着不可替代的作用,缺硼会导致细胞壁中硼的含量下降,但其在细胞中所占有的比例却明显提高[4]。缺硼还会影响植株根系的正常生长,降低根系的活力,致使根系吸收养分和水分的速率减慢,抑制根系的伸长和生长,降低了根系的吸收运输能力[5]。目前,世界农业生产中缺硼的现象普遍存在[6],不仅降低作物的产量,而且还会影响着作物的品质,在许多地区作物施用硼肥已经成为了增加产量、提高品质的一种常规性手段。
干旱胁迫会降低植株光合有关酶活性,影响光合系统的结构和特性,引起植物光合速率降低、生长受损,硼可以在一定程度上提升植株的光合速率、提高抗氧化物酶的活性,进而缓解干旱对植株造成的胁迫影响[7]。近年来有大量关于硼对干旱胁迫下玉米、柑橘、油菜、黄瓜等植株生长和抗氧化的研究[8-11],而在硼对干旱胁迫下厚皮甜瓜幼苗的生长和抗氧化系统的影响尚未见明确的研究和报道。本试验采用沙培育苗的方法,以相对含水量为20%的河沙模拟干旱条件,通过叶面喷施不同浓度的硼砂溶液,分析外源硼对干旱胁迫下厚皮甜瓜幼苗生长和抗氧化系统的影响,以期为硼肥提高甜瓜幼苗抗旱性研究提供理论依据。
1 材料与方法
1.1 试验材料
选用厚皮甜瓜品种‘农大103’,由河北农业大学栽培研究室提供。硼砂为化学分析纯,购自天津市瑞金特化工有限公司。
1.2 试验设计
试验于2020 年5—8 月在河北农业大学教学实验中心连栋温室进行。甜瓜幼苗采用沙培方式培育。当幼苗生长到三叶一心时,取长势一致、生长健壮的幼苗,定植于长0.6 m、宽0.4 m、高0.4 m 的大泡沫箱中,每个泡沫箱中定植4 株,泡沫箱中装满洗净消毒的细河沙,在河沙表面覆盖一薄层蛭石和珍珠岩用以减少水分的蒸腾,定植30 箱。在栽植完成后浇灌不含有任何硼成分的改良Hoagland 营养液,营养液每隔 3 d 浇灌1 次。人工模拟干旱环境,保持箱中河沙相对含水量在20%左右(通过滴灌时间控制)作为干旱条件[12],培养 7 d 后,开始进行叶面喷施硼砂溶液处理,硼砂浓度分别为B1 (5 mmol/L),B2(10 mmol/L),B3(20 mmol/L) 和B4(40 mmol/L),以清水作为对照(CK),每个处理设3 次重复,每2 箱为1 次重复,每隔 7 d 喷施1 次硼砂溶液,21 d 后,测定甜瓜幼苗叶片光合色素含量,光合特性及其它生长生理指标。
1.3 测定指标和方法
参照李合生等[13]的方法并加以改进测定超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、丙二醛(MDA)、超氧阴离子和根系活力。
采用SPAD-便携式叶绿素测定显示仪测定叶绿素a、b、a+b、类胡萝卜素,采用便携式光合仪测定甜瓜幼苗叶片气孔导度、胞间CO2浓度、蒸腾速率和叶片净光合速率。
计算壮苗指数(ISG)=(根鲜质量/地上部鲜质量+茎粗/株高)×全株鲜质量[14]。
叶片面积参照王颖等[15]方法测定。根、茎、叶鲜重和干重用电子天平称重。
1.4 数据统计分析
利用Excel 进行图表制作,用SPSS 16.0 软件进行数据显著性分析。
2 结果与分析
2.1 不同浓度的外源硼对干旱胁迫下甜瓜幼苗形态指标的影响
2.1.1 外源硼对干旱胁迫下甜瓜幼苗植株形态的影响 由表1 可知,喷施了外源硼的各浓度处理中,甜瓜幼苗的叶面积、茎粗、壮苗指数、株高等呈现出明显差异。B1 与对照(CK)相比较没有显著性差异,但高于对照;B2 显著高于B1 和CK;当B3 时,所有指标皆达到峰值,显著高于所有处理;然而, B4 却呈下降趋势,但显著高于B1、B2 和CK。
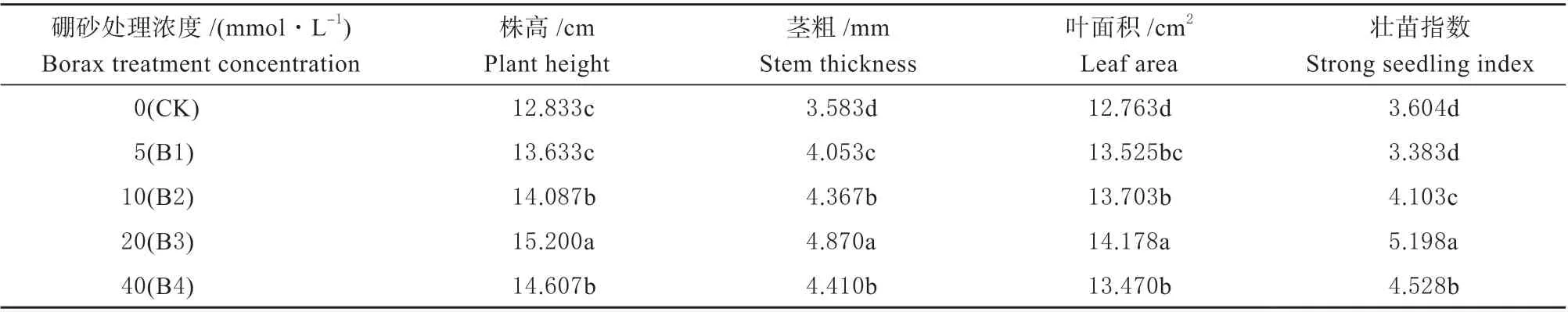
表1 外源硼对干旱胁迫下甜瓜幼苗生长量及植株形态的影响Table 1 Effects of exogenous boron on the growth and plant morphology of muskmelon seedlings under drought stress
2.1.2 外源硼对干旱胁迫下甜瓜幼苗地上部生长量的影响 由表2 可知,在经过外源硼溶液处理后,幼苗植株地上部的生长量,随着溶液浓度的增加,B3 处理植株地上部的生长量显著高于其它所有处理;B4 处理植株地上部的生长量与B2 相比差异不显著,但显著高于B1 处理和CK;B2 处理植株生长量显著高于B1 处理和CK;B1 处理植株生长量与CK 相比较差异不显著,但高于CK。
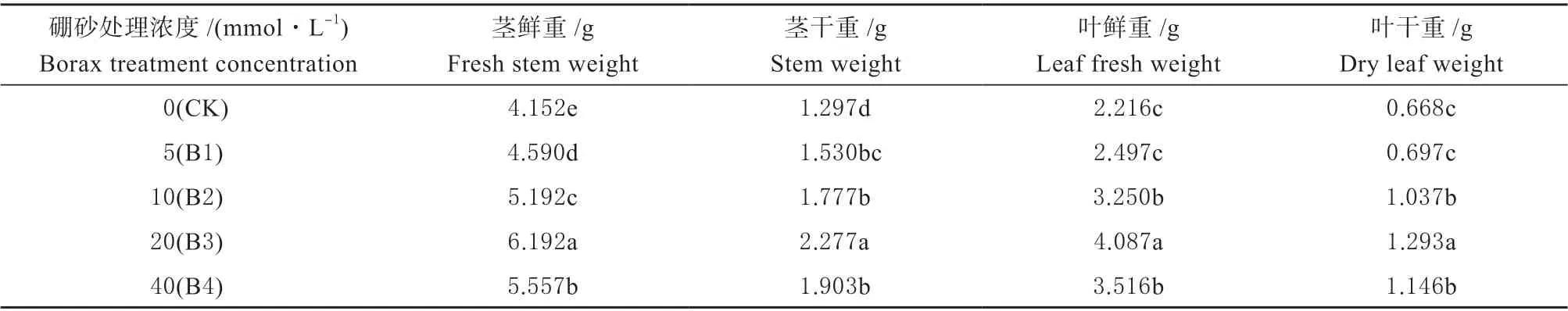
表2 外源硼对干旱胁迫下甜瓜幼苗地上部生长量的影响Table 2 The effect of exogenous boron on the above-ground growth of muskmelon seedlings under drought stress
2.1.3 外源硼对干旱胁迫下甜瓜幼苗地下部生长量的影响 由表3 可知,在甜瓜幼苗植株正常生长和经过外源硼溶液处理下植株地下部生长量存在着很大差异。B3 处理下的植株地下部生长量明显高于其它处理;B4 处理植株生长量低于B3 处理,但显著高于B2、B1 和CK;B2 处理下植株地下部生长量显著高于B1 和CK;B1 处理植株地下部生长量与CK相比差异不显著,但植株地下部生长量高于CK。

表3 外源硼对干旱胁迫下甜瓜幼苗地下部生长量的影响Table 3 The effect of exogenous boron on the underground growth of melon seedlings under drought stress
2.2 不同浓度外源硼处理对干旱胁迫下甜瓜幼苗叶片光合色素和光合特性的影响
2.2.1 外源硼对干旱胁迫下甜瓜幼苗叶片光合色素含量的影响 由表4 可知,在所有外源硼浓度处理中,叶绿素a、叶绿素b、叶绿素a+b、类胡萝卜素都随着处理浓度的升高有着明显变化,在B3 处理时光合色素含量达到最高,到B4 时开始下降,,但各浓度处理的光合色素含量均显著高于对照(CK)。B1 时,光合色素含量高于对照(CK),但与之相比差异性并不显著;B2 显著高于B1 和CK;当B4 时,所有上述指标都降低,但显著高于B2、B1 和CK。
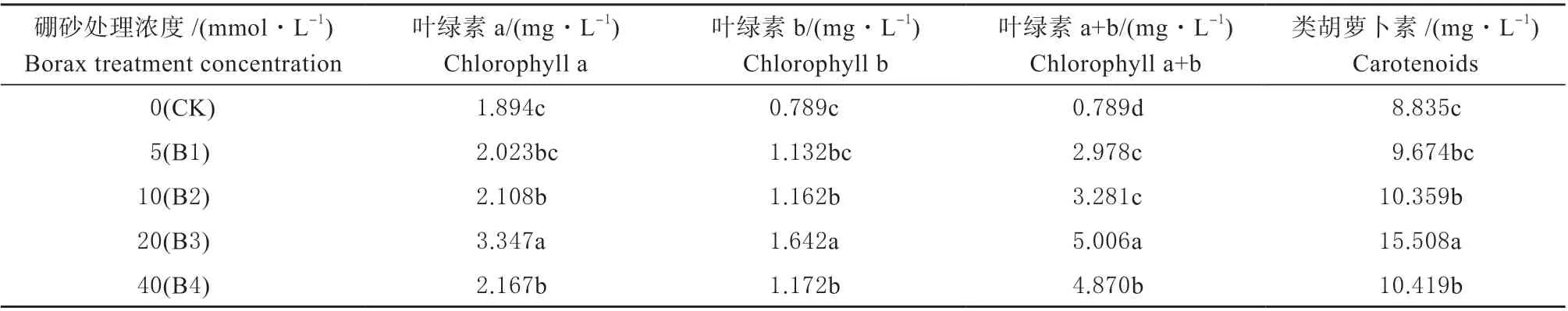
表4 外源硼对干旱胁迫下甜瓜幼苗叶片光合色素含量的影响Table 4 Effects of exogenous boron on photosynthetic pigment content in muskmelon seedling leaves under drought stress
2.2.2 外源硼对干旱胁迫下甜瓜幼苗光合特性的影响 由表5 可知,外源硼浓度处理显著提高甜瓜幼苗光合特性,随着喷施浓度的不断增加,甜瓜幼苗的净光合速率(Pn)和胞间二氧化碳浓度(Ci)与CK 相比变化明显。可以看出在B3 处理时Ci和Pn最高,显著高于B1、B2、B4 和CK;到B4 处理时,植株Ci和Pn呈下降趋势,显著高于B1 处理和CK,与B2处理相比差异不显著;B2 处理与B4 处理相比较差异不显著,显著高于B1 处理和CK;B1 处理与CK相比较差异不明显,但高于CK。气孔导度(Gs)和蒸腾速率的变化趋势同以上2 指标呈先升高后降低的趋势,各浓度处理和上述2 个指标变化规趋势相同,在B3 处理时达到最大值,显著高于所有处理水平。

表5 外源硼对干旱胁迫下甜瓜幼苗光合特性的影响Table 5 Effects of exogenous boron on photosynthetic characteristics of muskmelon seedlings under drought stress
2.3 不同浓度的外源硼对干旱胁迫下甜瓜幼苗抗氧化性的影响
2.3.1 外源硼对干旱胁迫下甜瓜幼苗超氧阴离子活性和丙二醛(MDA)含量的影响 如图1、2 可知,在进行外源硼溶液处理后随着溶液浓度的提升,植株体内的超氧阴离子活性和丙二醛(MDA)的含量与CK相比显著降低。
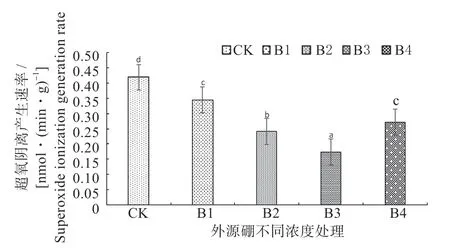
图1 外源硼对干旱胁迫下甜瓜幼苗超氧阴离子的影响Fig.1 The effect of exogenous boron on superoxide anion of muskmelon seedlings under drought stress
在B3 时,植株内的超氧阴离子活性和丙二醛(MDA)含量均处于最低值,显著低于其它浓度处理;B4 处理显著低于B1 处理和CK,与B2 处理相比较差异不显著;B2 处理显著低于B1 处理和CK;B1 处理与CK 相比差异不显著,但低于CK。
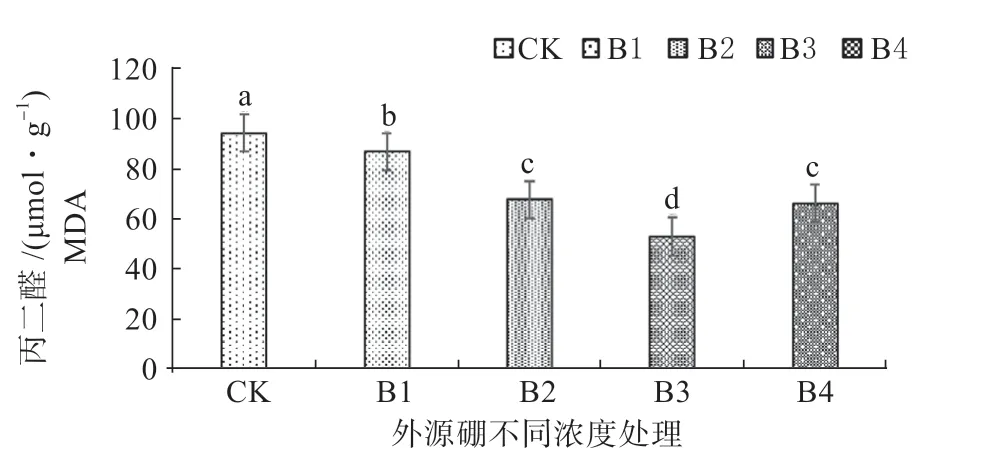
图2 外源硼对干旱胁迫下甜瓜幼苗丙二醛(MDA)的影响Fig. 2 The effect of exogenous boron on malondialdehyde (MDA) of muskmelon seedlings under drought stress
2.3.2 外源硼干旱胁迫下对甜瓜幼苗抗氧化性的影响 从图3 ~5 可以知,由于植株内部超氧阴离子和OH-等大量含氧自由基的增多,在外源硼溶液处理下超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)等抗氧化同工酶随着处理溶液浓度的逐渐升高,经外源硼处理的植株酶活性明显高于对照。其中,所有酶活的峰值出现在B3 处理,显著高于所有处理水平;与B3 处理相比B4 处理呈下降趋势低于B3 处理,但显著高于B1 处理、B2 处理和CK;B2 处理显著高于B1 处理和CK;B1 处理与CK 相比,B1 处理显著高于CK。
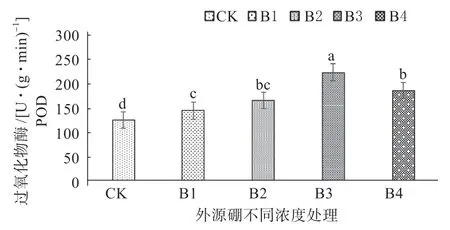
图3 外源硼干旱胁迫下对甜瓜幼苗过氧化物酶(POD)活性的影响Fig. 3 The effect of exogenous boron on the activity of peroxidase (POD) in muskmelon seedlings under drought stress
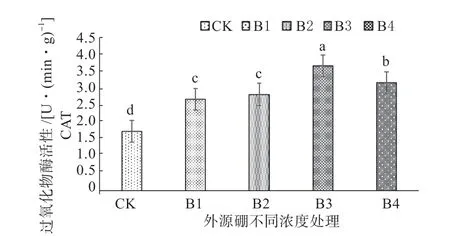
图4 外源硼干旱胁迫下对甜瓜幼苗过氧化氢酶(CAT)活性的影响Fig. 4 The effect of exogenous boron on catalase (CAT) activity of muskmelon seedlings under drought stress
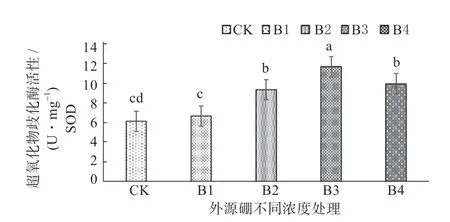
图5 外源硼干旱胁迫下对甜瓜幼苗超氧化物歧化酶(SOD)活性的影响Fig. 5 The effect of exogenous boron on the activity of superoxide dismutase (SOD) in muskmelon seedlings under drought stress
3 讨论与结论
干旱胁迫下经过不同的外源硼溶液处理,植株的生长量和形态指标表现出明显的差异。开始随着处理溶液浓度的升高植株的生长量快速提升、叶面积增大、茎秆增粗,但达到一定浓度后生长势开始下降植株长势开始减缓。试验结果与李自龙等[12]报道一致,叶面喷施适宜浓度的硼溶液可以提高植株地上部的生长量。
光合色素参与植物吸收、传递光能和引起原初光化学反应的重要色素,是促进植物正常进行光合作用的保障,硼参与调节光合机构的发育、促进光合色素的合成[16]。本试验结果表明,干旱胁迫下在5 ~40 mmol/L 浓度范围内,甜瓜幼苗叶片中叶绿素a、叶绿素b、叶绿素a+b 和类胡萝卜素含量呈先升高后下降的变化趋势,与熊博等[17]研究结果相一致。硼元素可以有利于促进植株幼苗对矿质元素的吸收,而硼元素可以直接或间接参与叶绿素的合成,还是植物叶肉细胞中叶绿体形成的重要组成部分[18],另外硼对植物叶细胞中栅栏组织和海绵组织也有着很大影响[19]。缺硼还会影响植株的根系生长,降低根系的活力,致使根系吸收水分和养分的速率减慢。然而在干旱胁迫下当硼浓度超出一定的阈值时,植物叶片形态发生变化,光合速率下降,这是因为硼浓度超过植物能忍受的最大值时,导致形成了盐害环境,引起细胞活性下降,细胞损伤[20]。
植株中缺硼会使植株叶片光合速率下降、蒸腾作用变缓、叶片光合特性降低,主要是因为缺硼会抑制叶片光合,气孔导度控制着植株的蒸腾速率强弱[21],导致气孔导度和蒸腾速率下降[22],干旱胁迫下植物体会受到伤害,叶片萎蔫,导致气孔关闭。干旱胁迫主要是气孔受限制、开度减少甚至关闭,造成植物体内水分亏缺,从而影响植物对CO2的吸收,从而降低植物光合速率[23]。植物叶片是植物进行光合作用积累养分的主要部位和场所。采用叶面喷施硼肥,有效硼能够通过叶片的气孔进入到植株体内,进而促进植物的生长[24]。本试验结果表明,在干旱胁迫条件下对甜瓜幼苗进行不同浓度的外源硼溶液处理发现,随着处理溶液浓度的升高幼苗植株中胞间二氧化碳浓度、净光合速率、气孔导度和蒸腾速率呈现出先增高后降低的变化趋势,与张晓文等[25]研究结果相一致。本试验中在叶面喷施不同浓度的硼溶液后,超氧阴离子和丙二醛(MDA)呈现出先升高后降低的变化曲线状态,而植株中CAT、SOD、POD 活性出现先降低后增高的变化趋势,与陈晟报道结果相似[26]。这是因为CAT 与SOD、POD 共同组成了生物体内活性氧防御系统[27],H2O2、超氧阴离子和MDA 的大量积累刺激了POD的活性增加[28],植株内过氧化氢酶、过氧化物酶和超氧化物歧化酶等通过彼此协调合作来清除细胞内的活性氧和影响植株正常生长发育的物质,抗氧化物酶活性的增高延缓了植株叶片的氧化衰老过程,维持了叶片细胞的活力[29]。干旱条件下,叶面喷施硼肥可以显著提高植株叶片中超氧化物歧化酶、过氧化氢酶、过氧化物酶的活性[30],植物细胞内的抗氧化酶SOD、POD、CAT 等含量升高,帮助清除了植物体内产生的活性氧, 提高植物抗旱性[31]。
试验结果表明,在干旱胁迫下对甜瓜幼苗进行外源硼叶面喷施处理,所有浓度处理均可以提升甜瓜植株的生长量和相关酶的活性;硼砂浓度在20 mmol/L 时幼苗植株所有的形态和生理指标皆达到最好水平。
