入侵植物紫茎泽兰在贵州的适生区预测
2022-02-19喻三鹏田茂娟张念念邱建生周倩班启明
喻三鹏田茂娟张念念邱建生周倩班启明
(1.贵州大学林学院,贵州 贵阳 550025;2.贵州省林业科学研究院,贵州 贵阳 550005;3.绥阳县林业局,贵州 绥阳 563300;4.望谟县林业局,贵州 望谟 552300)
紫茎泽兰Eupatorium adenophorum为菊科Asteraceae泽兰属Eupatorium多年生草本植物,原产中美洲热带及温带地区[1],是我国危害严重的外来入侵物种之一[2]。1865年,紫茎泽兰开始作为观赏植物栽培并引入夏威夷及英国,之后引种到澳大利亚及亚洲,后逸为野生[1,3-4],目前在30多个国家和地区都有分布,多国已经泛滥成灾[3]。紫茎泽兰于20世纪30年代由缅甸传入我国云南南部,1935年在云南南部首次发现[5]。20世纪70年代传入贵州兴义市的仓更镇和沧江乡[6],现已广泛分布于我国云南、贵州、四川、重庆、广西、西藏和台湾等地[5],目前仍不断向我国东部和北部快速扩散[2]。紫茎泽兰种子很轻,仅依靠风力就能传到很远的距离,且对环境的适应性很强,在林地、荒地、草地及山石地区都能生长繁殖[7],由于其根部分泌异种克生物质,能够抑制其他植物正常生长,形成密集的单优群落[8],造成生物多样性减少[9],对我国的农、林、牧业造成严重的经济损失[6,10]。
为了摸清紫茎泽兰在贵州省的适生范围,采用MaxEnt生态位模型对紫茎泽兰的适生区进行预测。MaxEnt生态位模型被广泛用来预测物种的适生区,是目前物种分布预测较为准确的模型之一[11-13]。对于紫茎泽兰的适生区预测研究,仅王翀 等[14]在大尺度范围内预测过紫茎泽兰的适生区,结果显示,在当前气候条件下,紫茎泽兰的主要分布区以广西、云南和贵州为主。在省级层面上对紫茎泽兰适生区的预测,未见报道。因此,对紫茎泽兰在贵州省的详细分布预测,将更有利于紫茎泽兰的精准防控。
1 数据来源与研究方法
1.1 数据来源
1.1.1 紫茎泽兰全球分布数据
紫茎泽兰全球数据分布通过3种方式获取。1)通过全球生物多样性信息网络(Global Biodiversity Information Facility,GBIF)数据库(https://www.gbif.org/)获取,对下载的数据点进行筛选,剔除无效数据点,得到紫茎泽兰经纬度分布点1 600个;2)通过在贵州省实际调查,获得40个紫茎泽兰分布点;3)通过文献获取紫茎泽兰分布点61个。3种方式共获得紫茎泽兰分布点1 701个。
1.1.2 环境因子数据
研究所使用的环境变量因子共20个,包含19个气候变量因子(bio01-19)和1个海拔变量因子(elevation)。气候因子包含当前气候模式数据(1970—2000年)和未来气候模式数据(2021—2040年、2041—2060 年、2061—2080 年、2081—2100年),主要包括研究区域内年、季度、月的温度变化等。20个环境因子均来源于世界气候数据库(https://worldclim.org/),当前气候模式数据空间分辨率约为100 hm2,未来气候模式数据空间分辨率约为2 100 hm2。
1.1.3 地图数据
贵州省行政区划图来源于国家基础地理信息中心(http://www.ngcc.cn)。
1.2 研究方法
1.2.1 MaxEnt模型
MaxEnt模型由 Phillips等[11,15]基于最大熵原理编写,广泛应用于物种的适生区预测[16-17],特别是在外来入侵生物的适生区预测方面应用较多[18]。
1.2.2 参数选择
以紫茎泽兰分布点、气候因子和海拔因子作为基础数据,导入MaxEnt模型中,选择绘制环境因子影响曲线(Creat response curves)、绘制分布预测图(Make pictures of predictions)、刀切法对环境因子的重要性分析(Do jacknife to measure variable importance),输出格式选择逻辑斯蒂格式(Logistic),模型重复运行3次,取平均值作为紫茎泽兰的适生区预测结果,设置25%的点作为验证集,通过验证集中的实际分布点和预测值,验证模型预测的准确性;由MaxEnt模型生成的受试者工作特征曲线(Receiver operating characteristic curve,ROC),用其曲线下面积(Area under curve,AUC)评价模型预测的准确性,AUC值越大,证明离随机分布越远,环境变量与模型对应地理分布的相关性越大,即模型预测效果越好[19]。
1.2.3 适生区划分
预测结果通过ArcGIS软件转化为栅格格式,用贵州省行政区划图对转化结果进行掩膜提取,通过空间分析对提取的栅格数据进行重分类,运用自然间断点分级法(Jenks),将紫茎泽兰的适生区划分为4个等级,分别为高适生区(0.473 605~0.691 651)、中适生区(0.323 396~<0.473 605)、低适生区(0.207 105~<0.323 396)和非适生区(0.071 432~<0.207 105),并得到紫茎泽兰在贵州省存在的阈值为0.207 105。
1.2.4 模型准确度检验
使用ROC曲线下AUC面积判断模型预测的效果,AUC的取值范围为0~1。当AUC的取值小于0.6时,表示模型预测不成功;当 AUC的取值为0.6~0.9时,模型预测精度可用;当 AUC值大于0.9时,表明模型预测精度高,效果好[20]。
2 结果与分析
2.1 紫茎泽兰在贵州的适生区
当前气候条件下,紫茎泽兰在贵州的高适生区主要位于黔西南州、六盘水市,以及安顺市和黔南州的部分地区,区域范围 (24.62°~26.53 °N,104.31°~107.53 °E),面积约 316.41 万 hm2,占全省区划面积的17.9%;中适生区主要位于毕节市、贵阳市、黔南州和安顺市的部分区域,区域范围(25.12°~27.76 °N,103.62°~108.95 °E),面积约372.73万hm2,占全省区划面积的21.1%。两者面积合计 689.14万 hm2,占全省行政区划面积的39.0%(图1)。总体看,紫茎泽兰的适生区面积较大,对区域经济、社会、生态均构成严重威胁。
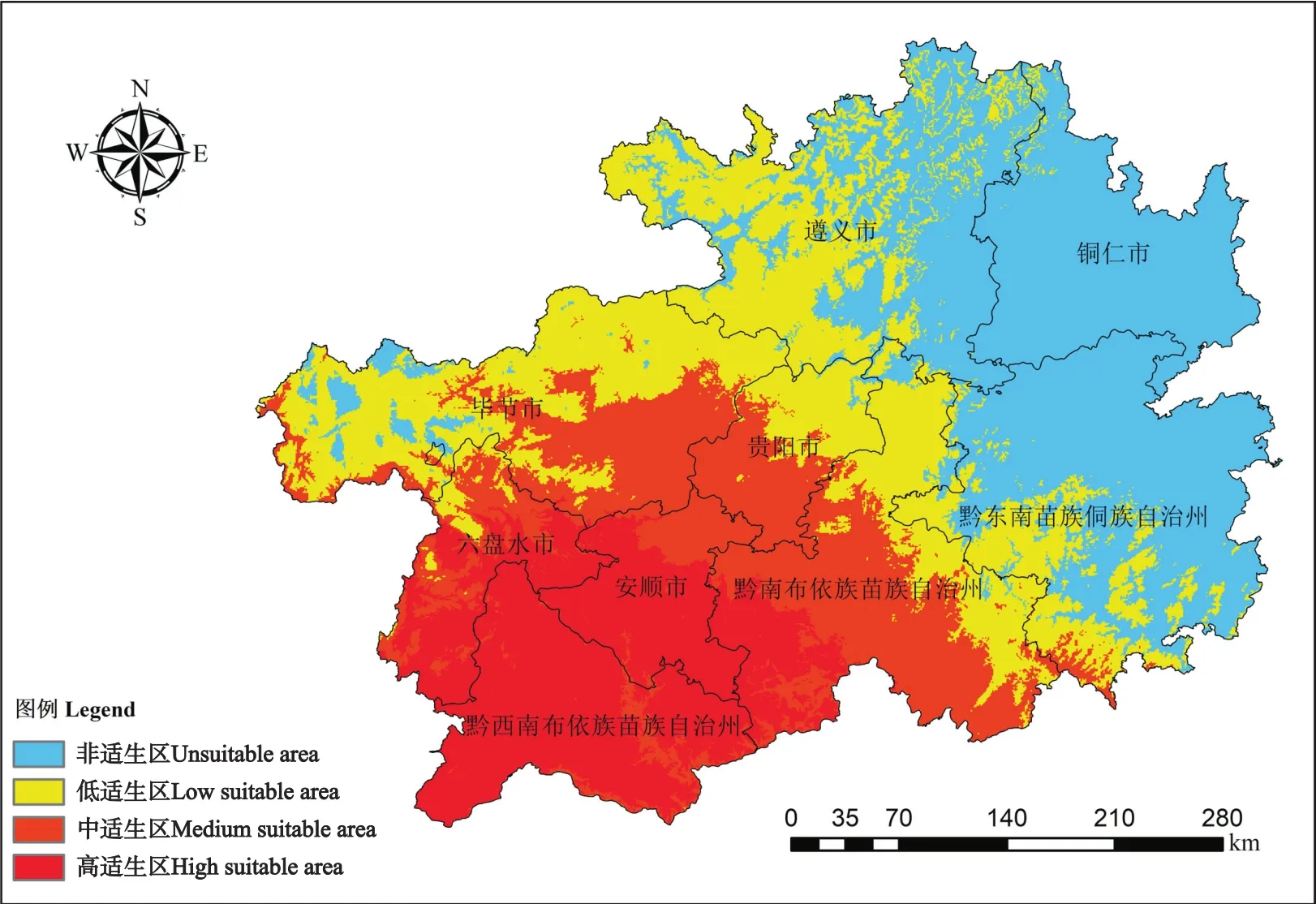
图1 紫茎泽兰在贵州的适生区预测Fig.1 Prediction of suitable area of E.adenophorum in Guizhou
未来气候条件下,紫茎泽兰在贵州的适生区呈先减后增的状态,整体向贵州北部方向扩散(图2)。2021—2040年,其扩张区将小于收缩区,适生面积减少约19.00万hm2;2041—2060年,其扩张区也将小于收缩区,适生面积减少约74.00万hm2;2061—2080年,紫茎泽兰将扩张113.00万hm2,几乎无收缩区域;2081—2100年,其在贵州的适生区面积将增加约213.00 万hm2,收缩区面积约10.00 万hm2。
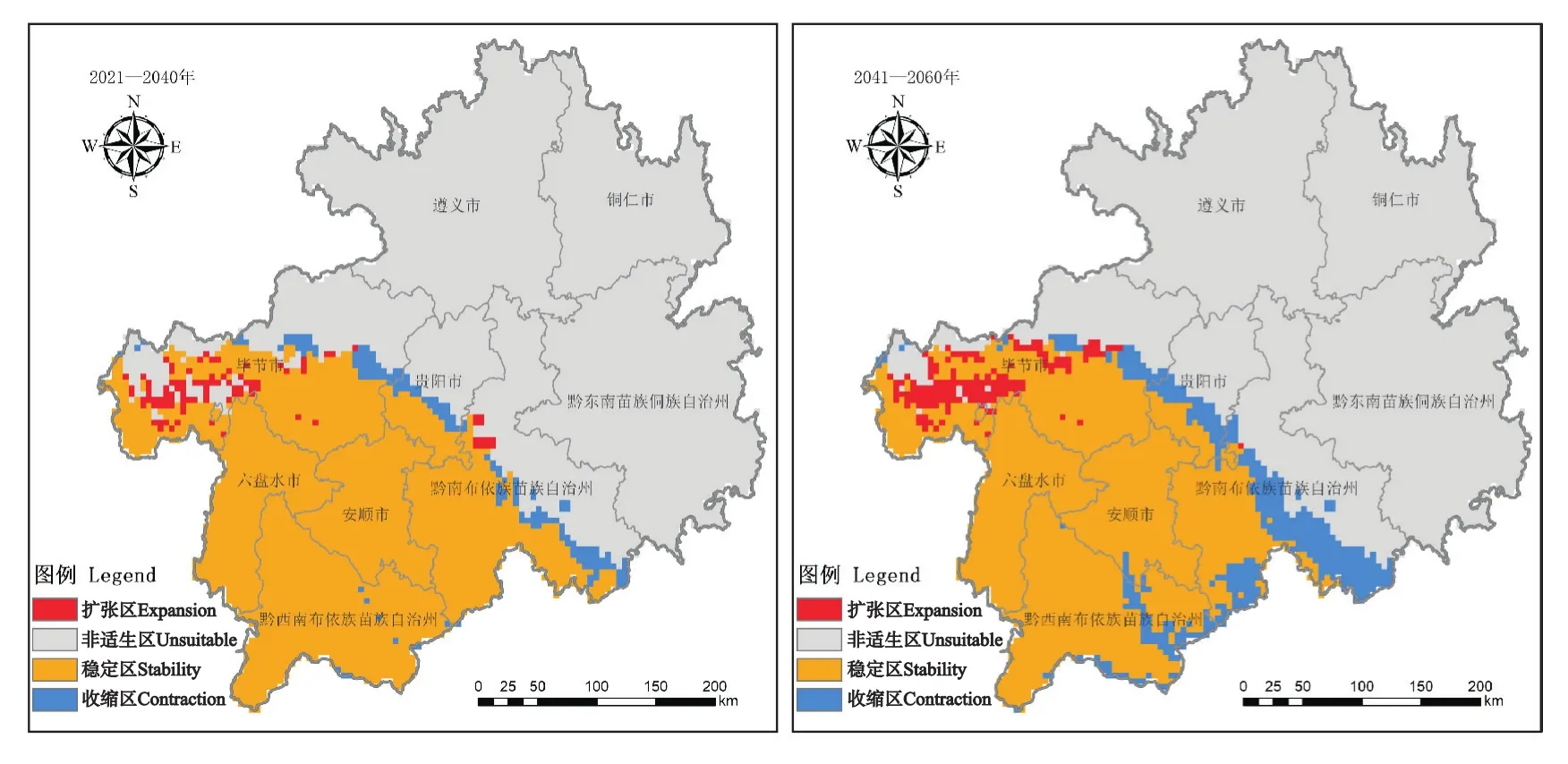
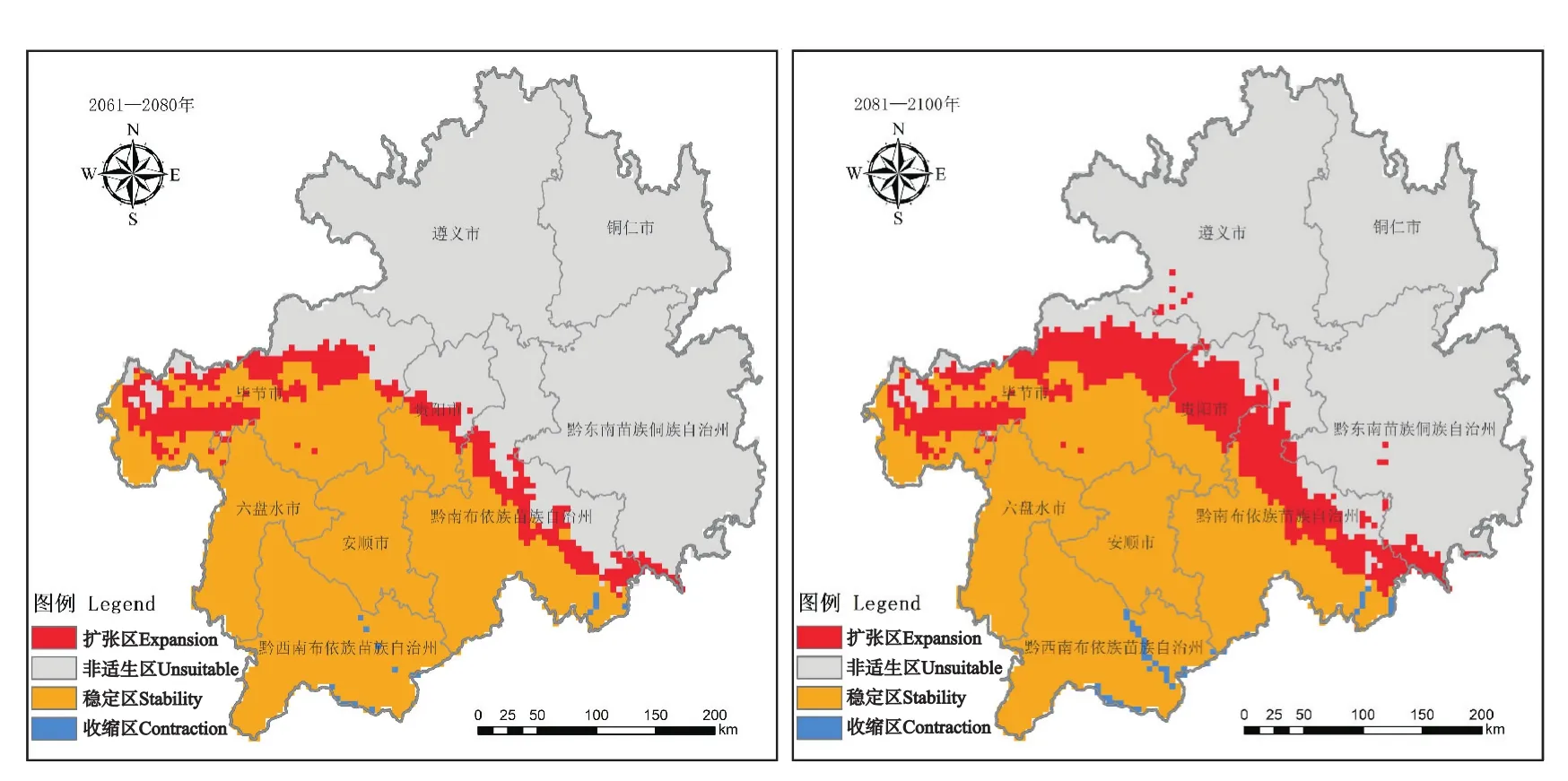
图2 未来不同时期紫茎泽兰在贵州的适生区变化Fig.2 Changes of suitable areas of E.adenophorum in Guizhou in different periods in the future
2.2 紫茎泽兰适生区预测模型准确性检验
根据MaxEnt运行生成预测模型遗漏率分析显示(图3),测试样本的遗漏率(Mean omission on test data)与预测遗漏率(Predicted omission)基本吻合,表明模型构建的效果较好,数据不存在空间自相关。根据受试者工作特征曲线显示(图4),AUC值(Mean AUC)为0.971,标准差(Mean+ /-one stddev)为0.001,显著高于随机分布模型(Random prediction)的AUC值0.5,说明该模型对紫茎泽兰的适生区预测效果优于随机分布模型,且曲线下面积值接近1,表明该模型预测结果具有较高的可信度,是有规律的非随机性预测结果。
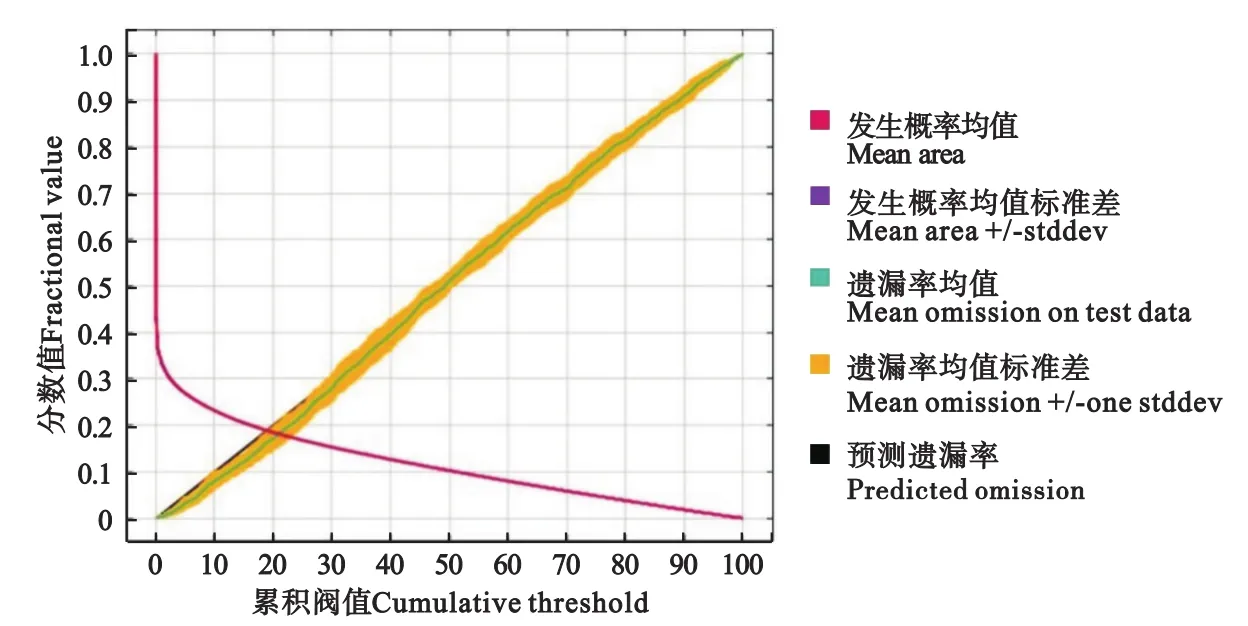
图3 预测模型遗漏率分析Fig.3 Prediction model omission rate analysis

图4 受试者工作特征曲线Fig.4 Receiver operating characteristic curve
2.3 主导因子筛选
2.3.1 环境因子对建模贡献率大小及置换重要性
环境变量最冷季度平均温度,最冷季度降水量,年平均温度对建模的贡献率依次分别为30.5%,16.6%,15.0%,累积贡献率达 62.1%,显著高于其余17个因子;环境变量最冷季度平均温度,年平均温度,年温度变化范围的置换重要性依次分别为 61.9%,8.3%,12.7%,累积置换重要性为82.9%,在建模过程中起关键性作用(表1)。可确定最冷季度平均温度、最冷季度降水量、年平均温度和年温度变化范围为影响紫茎泽兰分布的主要环境变量。

表1 环境变量贡献率及置换重要性Tab.1 Contribution rate of environmental variables and importance of permutation
2.3.2 刀切法检验
基于刀切法的环境变量重要性得分超过1.5的有年平均温度、最冷季度平均温度和温度季节变化3个变量(图5),表示三者对紫茎泽兰的分布预测有重要影响。昼夜温差月均值和海拔2个变量得分最低,表明其对紫茎泽兰的分布影响很小。得分超过1.7的有年平均温度和最冷季度平均温度,说明温度是影响紫茎泽兰分布的最主要因素。
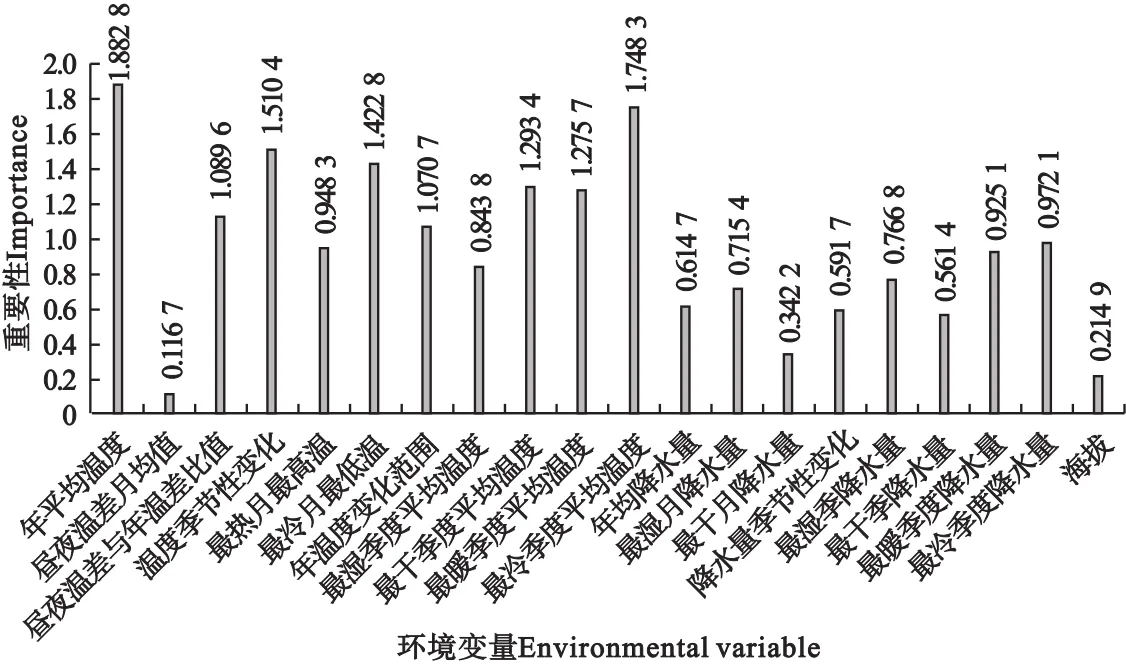
图5 基于刀切法的环境变量重要性Fig.5 Importance of environmental variables by Jackknife method
2.3.3 主导环境因子适宜范围
把MaxEnt模型绘制的仅适用单一环境因子的逻辑斯蒂曲线与影响紫茎泽兰分布的主要环境变量(最冷季度平均温度、最冷季度降水量、年平均温度和年温度变化范围)结合,以紫茎泽兰存在概率大于0.5为适宜范围[21],由主要气候因子影响曲线(图6)可以得出,最冷季度平均温度为8~15℃,最冷季度降水量为200~600 mm,年平均温度为14~18℃,年温度变化范围为-5~25℃时,紫茎泽兰的存在概率大于0.5。
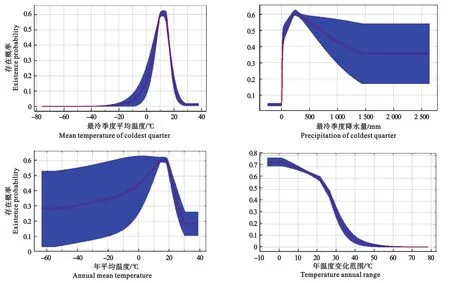
图6 主要气候因子影响曲线Fig.6 Response curves of main climate factors
3 结论与讨论
MaxEnt模型的刀切法检验和主导因子响应曲线分析表明,对紫茎泽兰适生区分布影响较大的环境因子有最冷季度平均温度,最冷季度降水量,年平均温度,年温度变化范围,其区间值依次分别为8~15℃,200~600 mm,14~18℃,-5~25℃。 由此可见,温度是影响紫茎泽兰分布的最主要因素,降水量次之。欧国腾 等[22-23]的研究表明,温度是影响紫茎泽兰生长的主要因素,对紫茎泽兰的定殖,生长、迁移和扩散非常重要。预测研究也表明,最冷季平均温度是影响紫茎泽兰潜在适生区的最重要变量[24]。
紫茎泽兰在贵州的高度适生区主要位于水城—六枝—普定—西秀区—长顺—惠水—罗甸一线以南地区,此区域年积温较高,冬季少雨雪、霜冻天气,冬季平均温度5℃以上,不易冻死,3—4月能够正常开花结实。中适生区主要位于水城—六枝—普定—西秀区—长顺—惠水—罗甸一线以北,威宁—赫章—大方—金沙—修文—龙里—都匀—荔波一带以南区域,冬季平均温度低于5℃,多雨雪、霜冻天气,冬季时,植株被冻伤或冻死,开花结实很少或不能开花结实,不适合生长。向业勋的研究表明,紫茎泽兰种子以每年60 km的速度向东北部传播[25]。但就目前贵州发生情况来看,种子传播后,受环境和气候的影响,很难正常生长发育和定殖扩散。
当前环境下的适生区基于1950—2000年数据预测获得,在全球气候不断变化的情况下,未来紫茎泽兰在贵州的适生区变化采用SSP1-2.6碳排放模式获得。在植物基因的研究中,已表明植物可通过OsICE1基因的去甲基化提高其耐寒性[26],这将有助于紫茎泽兰向温度更低的地区扩散蔓延。本研究为小尺度区域范围内研究,后续可进行大尺度区域范围研究,可为我国紫茎泽兰的防控提供参考区域。在今后的研究中,可引入坡度、坡向、空气湿度、土壤类型、人类活动等因子,更全面探索紫茎泽兰分布与环境的关系。
MaxEnt模型预测结果表明,紫茎泽兰在贵州的适生区范围广;未来气候条件下,紫茎泽兰在贵州的适生区呈先减后增状态,整体向贵州北部方向扩散。在环境变量贡献率和刀切法分析中,得到最冷季度平均温度、最冷季度降水量、年平均温度和年温度变化范围4个变量对紫茎泽兰的适生区分布影响较大。
紫茎泽兰在高适生区和中适生区内,持续影响农林经济及生态环境,为阻止其进一步的扩散和危害,提出以下管理思路以供参考。1)禁止紫茎泽兰以任何方式进入其他未传入地区,阻断传播源。2)在实际发生区域与未发生区域建立生态阻隔带,防止进一步扩散传播;实际发生区域内加大除治力度,减小种群面积。3)政府制定防控策略时,应参考紫茎泽兰适生区预测范围,高适生区内积极除治,中适生区内加强监测。4)根据紫茎泽兰生殖生物学特性,在2—4月开花结实期,对其进行物理割除,减少种子产生和传播。
