锡金小鼠中鼠疫噬菌体的分离及流行病学意义
2022-02-18杨丰义张海鹏吴鹤松钟佑宏
杨丰义,苏 超,张海鹏,吴鹤松,王 鹏,李 伟,钟佑宏
(1.云南省地方病防治所 云南省自然疫源性疾病防控技术重点实验室,大理 671000;2.中国疾病预防控制中心传染病预防控制所,北京 102206)
鼠疫是由鼠疫耶尔森菌引起的烈性传染病,具有病死率高,传染性强的特点,主要经鼠蚤传播,属广泛流行于野生啮齿动物间的一种自然疫源性疾病[1]。鼠疫在世界范围内曾有过三次大流行,至少导致一亿六千万人死亡[2-5],其中1772-1964年间发生的第三次大流行被认为起源于云南[6]。云南省地处我国西南边陲,与人间鼠疫渊源极久,早在公元1108年即有可疑人间鼠疫流行的记载[7],近年来云南鼠疫仍较活跃,2016年发生人间病例[8],因此鼠疫防控仍是云南重要的公共卫生问题。
锡金小鼠属于野生啮齿动物的一种,可携带着许多与人类疾病相关的病原体,鼠疫耶尔森菌便是其中的一种病原体[9]。1983年,就曾在两份锡金小鼠材料中检出鼠疫抗原阳性(云南省流研所资料汇编),但锡金小鼠是否为云南鼠疫储存宿主并可在家鼠和野鼠间传播鼠疫尚不清楚。
1915-1917年英国的Edward和法国的D'Herelle最先报道鼠疫噬菌体,可用于进行鼠疫病原体的鉴定[10-11]。国外的相关研究显示鼠疫噬菌体广泛存在于自然界[12],且鼠疫噬菌体与鼠疫菌相互依存,分离到鼠疫噬菌体应考虑到该材料中存在鼠疫菌或曾被鼠疫菌感染过。本研究在云南鼠疫疫源地中捕获锡金小鼠,了解云南锡金小鼠是否携带有鼠疫噬菌体或鼠疫菌,为研究锡金小鼠在鼠疫传播中的作用等方面提供线索。
1 材料与方法
1.1 材料及试剂鼠疫菌疫苗株EV76,LB液体、半固体及固体培养基;肠道标本DNA提取试剂盒、2×TaqPCR MasteMix购自天根生物科技(北京)有限公司;冷增菌液PEptone Sorbitol Bile Broth购自Sigma-Aldrich公司。
1.2 调查地点2016-2017年,在云南家鼠鼠疫疫源地的3个调查点(宜良县、梁河县、文山市)进行鼠疫宿主动物调查。由于锡金小鼠活动范围广,主要采用夹夜法,以自制油条为诱饵进行捕获并进行属种鉴定。
1.3 样本采集对调查中捕获经鉴定后的锡金小鼠立即在生物安全柜中无菌条件下解剖,取2 cm左右肠道标本,将其剪碎后放置于含有5 mL冷增菌液的15 mL离心管中,充分混匀后保存于4℃冰箱保存备用。
1.4 鼠疫噬菌体的分离与纯化将上述冷增菌液用0.22 μm滤膜过滤,以鼠疫疫苗株(EV76)为宿主菌,28℃下双层平板法分离培养,18~24 h后观察噬菌斑的生长情况[13]。有噬菌斑的噬菌体菌株,单斑增殖4~6次提升效价及纯度后[14],可用于保存及噬菌体的电镜观察。
1.5 鼠疫噬菌体电镜扫描取200 μL对数生长期的EV76菌悬液加入至6 mL冷却至45℃的LB半固体培养基中,轻微摇匀后倾倒LB薄层固体培养基。待双层平板凝固后滴加10 μL待测噬菌体滤液(噬菌体效价至少达108PFU/mL),待噬菌体滤液吸收后正置于28℃恒温培养箱中培养。16~20 h形成大噬菌斑,用接种环扣下有噬菌斑的上层半固体,浸泡于200 μL SM液中过夜,次日以5000 ×g离心5 min后取上清液,并用醋酸铀染色(注意严格把控染色时间)制备电镜标本,透射电镜下观察鼠疫噬菌体形态结构。
1.6 鼠疫菌基因检测按照TIANamp Genomic DNA KitDNA提取试剂盒要求,对鼠肠道标本总DNA进行提取。根据参考文献[15]对鼠疫菌的特异标识基因caf1进行检测,对鼠肠道标本进行鼠疫菌的初筛。引物序列如下:caf1/F:5'-GGAACCACTAGCACATCTGTT-3';caf1/R:5'-ACAGCATCAGTGTATTTACCTGCTG-3'。对于某些缺失pMT1质粒的鼠疫菌株,可采用董珊珊等[16]建立的可同时检测caf1及YPO0392基因的一体系双重荧光定量PCR方法对鼠疫菌基因进行检测,避免漏检。但目前研究中尚未发现云南鼠疫菌株有缺失pMT1的情况,所以此次研究采用检测caf1基因的方法来进行鼠疫菌的初筛。
1.7 宿主谱裂解实验本次实验选取24株肠道菌株作为实验菌株(表3),采用点滴法测定锡金小鼠中分离的3株鼠疫噬菌体的裂解谱。分别置于28℃、37℃恒温培养箱中培养18~24 h,次日观察结果。
1.8 统计分析应用Microsoft Excel进行数据统计整理。
2 结果
2.1 锡金小鼠的捕获情况在云南鼠疫疫源地3个调查点中共捕获86只锡金小鼠,其中宜良县29只、梁河县11只、文山市46只。
2.2 鼠疫噬菌体分离结果86只锡金小鼠共分离出3株鼠疫噬菌体,分别在梁河县分离到1株,分离率9.09%(1/11);文山市分离到2株,分离率4.35%(2/46);宜良县未分离到鼠疫噬菌体(表2)。

表2 云南锡金小鼠中鼠疫噬菌体分离情况表Table 2 Y. pestis phages isolated from Mus pahari in Yunnan province
2.3 鼠疫噬菌体噬菌斑的生长多样性双层平板法铺板28℃孵育24 h后,通过肉眼观察,3份鼠疫噬菌体的噬菌斑表现出生长多样性。其中LHS 274的噬菌斑表现为小噬斑(<0.5 mm),28℃下噬菌斑非常模糊,在37℃下培养18~24 h后噬菌斑变得稍清晰;WSS 81的噬菌斑表现为中噬菌斑(直径约1 mm),噬菌斑透亮;WSS 198的噬菌斑为特大噬菌斑(直径>3 mm),噬菌斑透明,边界清晰(图1)。

图1 云南锡金小鼠肠道分离出的3株鼠疫噬菌体噬菌斑图Fig.1 The plaques of 3 Yersinia pestis phages isolated from the intestine of Yunnan Mus pahari
2.4 鼠疫噬菌体电镜下形态观察对3株鼠疫噬菌体进行电镜扫描显示,两株均为较典型的肌尾型鼠疫噬菌体。其中LHS 274的头部直径约52 nm,尾部约118 nm;WSS 81的头部直径约70 nm,尾部约100 nm;WSS 198的头部直径约45 nm,尾部约135 nm。

图2 电镜下锡金小鼠肠道中分离出的3株鼠疫噬菌体形态(×100 000)Fig.2 Morphology of 3 Yersinia pestis phages isolated from the intestine of Mus pahari under electron microscope (×100 000)
2.5 鼠疫菌的特异标识基因caf1检测结果86只锡金小鼠脾脏、肝脏、肠道中特异标识基因caf1进行检测的结果均为阴性。
2.6 3株鼠疫噬菌体宿主谱分析3株鼠疫噬菌体在28℃及37℃下除裂解EV76外,WSS 81还可裂解福氏1a、福氏2a、福氏3a、福氏X变种、索氏志贺、宋内Ⅰ、宋内Ⅱ、ATCC25922及ATCC35218;WSS 198还可裂解福氏Ⅹ变种及ATCC25922;LHS 274可裂解PSTⅠ、PSTⅣ、PSTⅤ、PSTⅥ,此外LHS 274在37℃温度下还可裂解PSTⅡ(表3)。
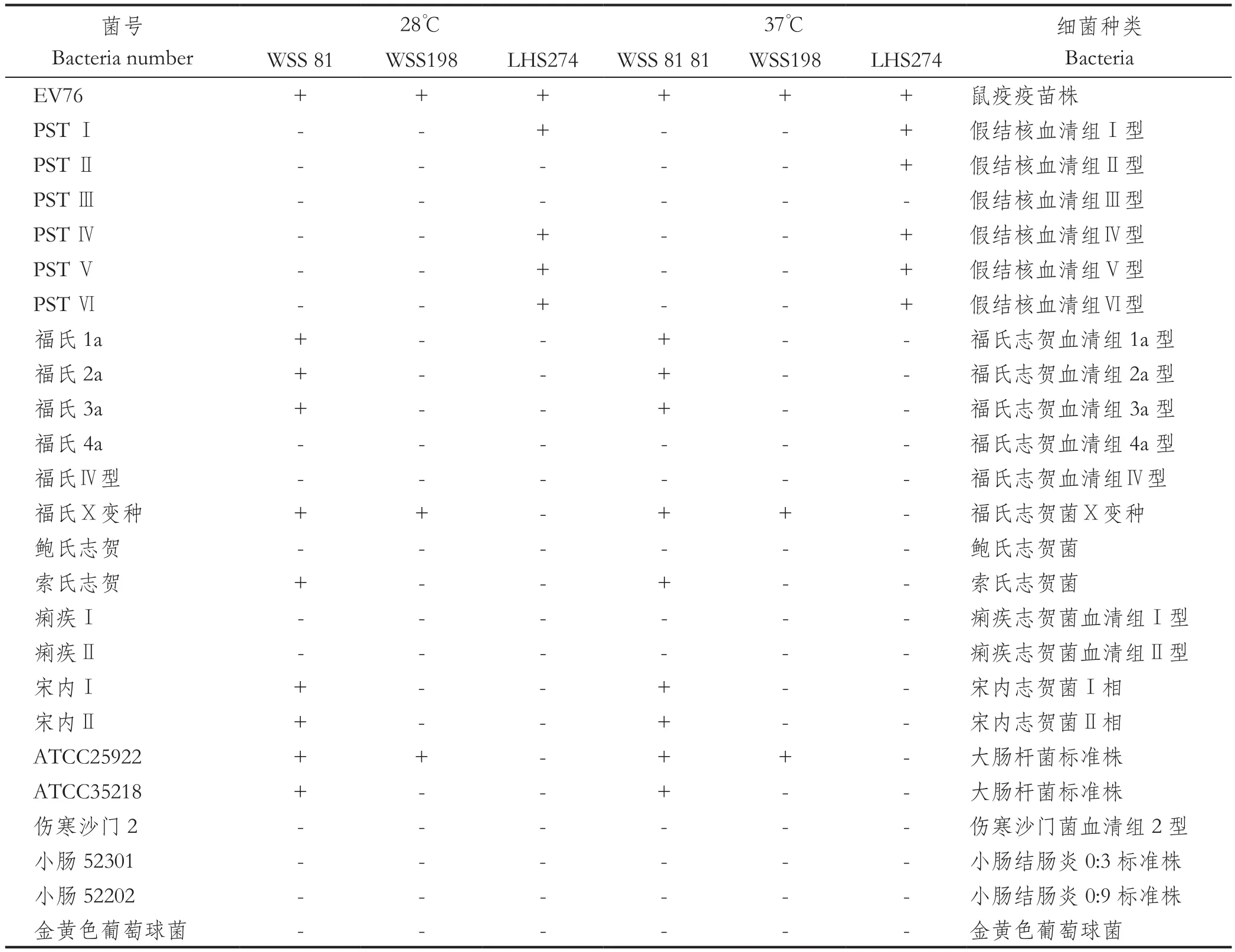
表3 3株鼠疫噬菌体宿主谱结果Table 3 Results of host spectrum of 3 Yersinia pestis phages
3 讨论
3.1 锡金小鼠与鼠疫的关系锡金小鼠(Mus pahari)属于野栖小型鼠类,隶属于鼠科中的小鼠属。分布区域相对较广,在我国首先于云南与缅甸接壤的南丁河被发现,云、贵、川等地均有分布[17],在滇南山地、林区为优势种[18-19]。在梁河县高(1880-1978 m)、中(1475-1490 m)、低(1068-1246 m)三个不同海拔地区均有分布,且在高海拔地区为优势种,跨带分布能力强[20]。
1983年,在陇川县景坎公社芒软山区发生鼠间鼠疫,从该山区耕作地捕获的锡金小鼠中检出鼠疫抗原阳性(云南省流研所资料汇编)。根据文献[21]分析总结,锡金小鼠可携带印鼠客蚤、特新蚤指名亚种、无孔微棒蚤直指亚种等鼠蚤。本次研究的3个调查点都为家鼠鼠疫疫源地,印鼠客蚤为其主要传播媒介[22]。特新蚤指名亚种为云南野鼠鼠疫疫源地的主要传播媒介;无孔微棒蚤直指亚种在云南省家鼠鼠疫疫源地数量较多且分布广,1991年还从两匹无孔微棒蚤直指亚种中分离出鼠疫菌[23]。蚤类既是鼠疫菌的储存宿主,又是传播媒介,在鼠疫流行传播中承担重要角色。鼠类作为鼠蚤的寄主,锡金小鼠通过鼠疫菌的鼠-蚤-鼠传播,可能会在家-野型鼠疫疫源地传播中发挥作用。
3.2 云南锡金小鼠中存在鼠疫噬菌体此次噬菌体分离实验共从86只锡金小鼠中成功分离出3株鼠疫噬菌体,分离率为3.49%,证明锡金小鼠中存在一定量的鼠疫噬菌体。从分离率上看,锡金小鼠中鼠疫噬菌体分离率低于玉龙野鼠鼠疫疫源地中储存宿主齐氏姬鼠的分离率(10.85%),高于流行宿主大绒鼠的分离率(2.60%)[13];而与鹤庆野鼠鼠疫疫源地及剑川野鼠鼠疫疫源地相比,其分离率均高于主要宿主齐氏姬鼠(鹤庆3.45%,剑川2.40%)及大绒鼠(鹤庆为0%和剑川0.42%)[14,24]。锡金小鼠中分离出鼠疫噬菌体,不排除其感染过鼠疫菌的可能性,如果是这样,在感染后又康复,那么锡金小鼠在鼠疫流行中就可充当储存宿主及鼠疫的流行宿主,由此锡金小鼠在鼠疫流行中就具有较大的流行病学意义,值得进一步关注。
3.3 3株鼠疫噬菌体具有宽宿主谱3株鼠疫噬菌体除在28℃及37℃下均裂解鼠疫疫苗株EV76外,还可裂解非鼠疫菌株。与钟佑宏等[25]研究的LJ-7的宿主谱(除鼠疫菌外,只裂解福氏志贺菌)相比,3株鼠疫噬菌体除裂解EV76外,WSS81及WSS198还可裂解大肠杆菌、志贺菌属,LHS274还可裂解5株假结核耶尔森菌,宿主谱范围更广。从微生态学方面来说,当鼠疫噬菌体有除鼠疫菌外的其他宿主菌存在时,鼠疫噬菌体可以不依赖鼠疫菌生长繁殖。那么针对此次有鼠疫噬菌体存在,但未检出鼠疫菌,一方面可能就是由于这3株噬菌体依赖于其他宿主菌生存;另一方面这是否与噬菌体存活时间长及二者之间存在协同共生又此消彼长有关,噬菌体的种群扩大抑制了鼠疫菌的生长。由此具有宽宿主谱的鼠疫噬菌体可能会在动物间鼠疫中起到一个调节作用,这对以后将微生态学应用于鼠疫防治有重要意义。
3.4 噬菌体多态性此次分离的3株鼠疫噬菌体,从噬菌斑大小及状态来看,有3种形态,LHS274表现为针尖样大小的小噬菌斑;WSS81表现为直径约1 mm的中噬菌斑,噬菌斑形态较LHS274好;WSS198表现为直径>3 mm的特大噬菌斑,噬菌斑透亮。以上鼠疫噬菌体与玉龙,鹤庆及剑川所分离到的鼠疫噬菌体相比较[13-14,24],LHS274噬菌斑大小及状态除与玉龙野鼠鼠疫疫源地中齐氏姬鼠中分离到的LJ 123及大绒鼠中分离到的LJ218噬菌斑相似,与上述三处鼠中分离所得的其他鼠疫噬菌体均有差异。WSS 81与鹤庆疫源地中分离的鼠疫噬菌体HQ1、HQ2噬菌斑大小相似,但噬菌斑更为透亮,与其他几株鼠疫噬菌体无相似性。WSS198与以上三个野鼠鼠疫疫源地鼠种中分离的噬菌体对比,噬菌斑均较大。不论是此次分离的3株噬菌体比较或是与相关文献已分离的比较,噬菌斑均具有多态性,加之宿主谱有差异,由此推断噬菌体具有多态性,有待进一步深入研究。
