枯草芽孢杆菌细菌素研究进展
2022-02-15范修强喻家晓汪招雄张明辉高学军
范修强,喻家晓,刘 峰,汪招雄,张明辉,高学军
(长江大学动物科学学院,湖北 荆州 434023)
枯草芽孢杆菌(Bacillus subtilis)是芽孢杆菌属的一种,为革兰氏阳性需氧菌,广泛分布在土壤及腐败的枯草等有机物中。该菌无荚膜、有鞭毛,菌落表面粗糙, 呈白色或微黄色。菌体生长速度快, 对营养要求较低,在普通碳源培养基中生长良好,在工业生产条件下保持较好的活力。在生殖生长期,幼龄B.subtilis能产生大量芽孢,在生长环境恶劣、营养物质缺乏等不适宜的环境下进入孢子休眠期,形成抗逆、耐高温、耐酸碱等抗性作用。在环境适宜生长条件下,芽胞会重新生长为B.subtilis[1]。B.subtilis是一种重要的益生菌种,不含内毒素和无致病性,能够产生多种抗菌活性物质,还能够分泌淀粉酶、脂肪酶、蛋白酶、和纤维素酶等多种内源酶,被美国FDA 认定为“Generally regarded as safe”(GRAS)级别益生菌种,在医药、食品、农业、畜牧业、水环境改善等方面用途很广[2]。B.subtilis在中国现已作为益生菌被批准为饲料添加剂。此外,该菌无显著的密码子偏爱性,适于作为“细菌工厂”生产外源重组蛋白。
1945 年首次发现B.subtilis可以产生抗菌活性物质[3],至今已发现B.subtilis能产生几十种不同分子结构的抗生素(antibiotics),包括由核糖体合成的多肽(又称为Bacteriocins,细菌素)、非核糖体的多肽合成酶催化形成的多肽以及非肽类化合物(包括聚酮类化合物、磷脂类化合物等)。多肽类抗生素是B.subtilis最主要的抗菌物质,细菌素抗菌谱较窄,主要抑制与自身种属较密切的细菌;而非核糖体合成的多肽具有广谱抑制细菌和真菌的活性[4]。随着基因组学的快速进展,新的细菌素正在不断被认知,是传统抗生素的有前途的替代品。与非核糖体合成的代谢产物相比,由核糖体合成的细菌素更易于经基因工程等分子生物学技术进行生产。文章主要概述了枯草芽孢杆菌产生细菌素的研究进展,旨在为细菌素(基因)的开发应用提供理论依据。
1 枯草芽孢杆菌细菌素的分类
在革兰氏阳性菌中,芽孢杆菌属是一个十分多样的菌属,具有200 多个亚种[5]。芽孢杆菌属产生的细菌素具有不同的分子大小、序列、结构和抗菌活性,其分类与乳酸菌细菌素分类不同。芽孢杆菌细菌素依据分子结构特征,不同细菌产生的细菌素被分为3 类(Ⅰ~Ⅲ)[5-7]。
Ⅰ类细菌素,又称为羊毛硫细菌素(lantibiotics),为小分子阳离子多肽(<5kDa),具有两亲性结构,作用于生物膜,氨基酸序列中含有特殊的羊毛硫氨酸(lanthionine)和甲基羊毛硫氨酸(methyllanthionine),因而多肽序列内部形成多个环。Ⅰ类细菌素又分为4个亚类:Ⅰ.1 亚类为单链多肽、相对长的形状,例如subtilin;Ⅰ.2 亚类为单链多肽、具有球形或其它的形状,例如mersacidin;Ⅰ.3 亚类有两个多肽组成,例如haloduracin(其包括Halα 和Halβ 2 个多肽);Ⅰ.4 亚类为环状多肽,例如Subtilosin A。Ⅱ类细菌素不含修饰的氨基酸,为线性小分子多肽(0.77k~10kDa)。Ⅱ类细菌素又分为3 个亚类。Ⅱ.1 亚类细菌素含有37~48个氨基酸残基,在多肽链N-末端具有保守的YGNGV/L 基序,例如pediocin;Ⅱ.2 亚类细菌素在多肽链N-末端具有保守的DWTXWSXL 基序,例如thuricin H;Ⅱ.3 亚类细菌素为没有保守的N-端基序的其它线性多肽,例如cerein 7A。Ⅲ类细菌素为具有较高分子质量(>30kDa)的没有修饰的蛋白质,对热不稳定,例如megacin A-216。
2 枯草芽孢杆菌细菌素的结构特征、作用机理和抑菌谱
多数B.subtilis细菌素具有类似的结构特征,因此也具有类似的分子机理和抑菌谱。多数B.subtilis细菌素具有多肽链N-端具有带有正电荷的亲水基团,能够结合细菌细胞膜的磷脂,当这一结合发生后,疏水的C-端区域能够穿透至疏水的细胞膜内部,使细胞膜形成穿孔而导致细胞死亡[5]。Ⅰ.3 亚类由2 种多肽形成,这2 种多肽的基因在同一操纵子中彼此相邻,2 种多肽以1:1 的比例共同形成一种膜穿透螺旋-螺旋结构[6]。B.subtilis细菌素主要抑制革兰氏阳性菌包括其他芽孢杆菌、单增李斯特菌、金黄色葡萄球菌等,其抑制机理在于这些革兰氏阳性菌具有细菌素能够结合的含肽聚糖的细胞壁,而革兰氏阴性菌不具有细菌素能够结合的相应细胞壁结构[5,8,9]。B.subtilis细菌素是天然合成的,具有热稳定性、耐强碱、水溶性好、抗菌谱广、抗菌活性高、不易产生耐药性等特点[10,11]。
Ⅰ细菌素subtilin 是B. subtilisATCC 6633 产生的最有效的lantibiotic 细菌素,研究工作也最全面和深入[5,9]。以下以subtilin 为例,说明枯草芽孢杆菌细菌素的结构特征、作用机理和抑菌谱。Subtilin多肽链具有56 个氨基酸残基,结构分为24 个氨基酸残基的N-端前导肽(leader peptide)序列和32 个氨基酸残基的C-端核心肽(core peptide)序列2 个部分。前导肽(leader peptide)序列不含Cys 残基,与细菌素分泌到细胞外有关,在其分泌至细胞外时被切除。核心肽含有Ser、Thr 和Cys 氨基酸残基,是lanthionine 和methyllanthionine 硫醚环形成所需要的。Ser 和Thr 分别脱水生成2,3-二氢丙氨酸(2,3-didehydroalanine,Dha)和2,3-二氢氨基丁酸(2,3-didehydrobutyrine,Dhb),Dha 和Dhb 与Cys 的巯基反应,使肽链形成环状结构(图1)[9]。这样的分子结构使得subtilin 能够结合细胞壁发挥作用,并且分子结构具有耐热、耐酸碱等特征。Subtilin 能够靶向细胞壁的前体物lipid II 的焦磷酸杆菌烯基抑制肽聚糖(peptidoglycan)的组装,使敏感细菌细胞膜形成穿孔和透性化导致具体死亡[12]。 Subtilin 与nisin类似,对革兰氏阳性菌具有广谱的抑制作用,如抑制其他芽孢杆菌属菌株、乳球菌属(Lactococcus spp.)和藤黄微球菌(Micrococcus luteus),其对M.luteus菌株NDCO8166 的最小抑制浓度(minimal inhibitory concentration,MIC)为0.05μg/mL[5,9]。
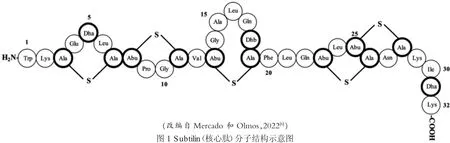
图1 Subtilin(核心肽)分子结构示意图
3 枯草芽孢杆菌细菌素的基因簇
3.1 枯草芽孢杆菌基因组简介
早在1997 年,已完成模式菌B.subtilis168 的全基因组DNA 序列测定(登录号AL009126)[13]。近几年近百个B.subtilis亚种(或菌株)的全基因组DNA序列测定业已完成,提供了更多的B.subtilis细菌素基因簇的信息。例如,Bacillus subtilisMJ01 染色体基因组大小为4108293 bp,其中有4269 个编码序列[14]。B.subtilisN2-20 基因组大小为4036899bp,含有4163 个编码序列,在抗菌物质合成方面,N2-20含有10 个基因簇、168 菌株含有11 个基因簇[15]。B.subtilisCGMCC 12426 完整基因组序列由1 个环形的4138265bp 的染色体与1 个74165 bp 的质粒组成,共编码4222 个编码序列[16]。在B.subtilis基因组中,与抗菌代谢产物合成相关的基因总共达到基因组的4-5%。近年来通过基因组挖掘和蛋白分离纯化,新发现一些B.subtilis细菌素,例如:BAC-IB17[17],mejucin[18],Subtilin L-Q11[19]等。还有很多新发现的细菌素,尚不清楚其具体分子结构、编码基因或编码相应合成酶的基因以及分子作用机理,这些细菌素被称为细菌素样抑制物质(bacteriocin-like inhibitory substances,BLIS)[6]。尚有很多编码序列的多肽产物依据序列特征被预测为细菌素,有待实验证实。
3.2 细菌素基因簇结构特征
大部分细菌素的基因和相关基因以基因簇的形式存在,包括细菌素结构基因、负责转运细菌素转运的基因、细菌素自身免疫基因(保护细菌不被自身的细菌素所破坏)、细菌素表达调节基因、细菌素加工成熟所需酶类基因。细菌素结构基因表达产物为带有N-端信号肽的未成熟蛋白,在运输到细胞外时信号肽被细菌产生的酶所切除[8]。
细菌素的基因簇具有相似的结构,以下以subtilin 基因簇为例进行具体说明[8]。Subtilin 基因簇总长度为12kb,包含10 个基因:spaBTCSIFEGRK。spaS 为细菌素结构基因,编码一个56aa 的subtilin前体蛋白;spaB 和spaC 为修饰subtilin 前体蛋白产生lanthionine 和methyllanthionine 环状结构的酶类;spaT 编码ABC 转运蛋白,该蛋白定位于细胞膜,将subtilin 前体蛋白运输到胞外;spaI 表达蛋白形成一个膜结合的脂肽,通过结合subtilin 起到自身免疫作用;spaF、spaE 和spaG 表达蛋白在细胞膜上形成复合物,将subtilin 分子从细胞内泵出到细胞外,同时也起到自身免疫作用;spaK 表达一个感应(sensor)激酶,能够识别subtilin,进而通过磷酸化激活spaR 表达的应答调节因子spaR 蛋白,磷酸化spaR 能够激活spaBTCSIFEG 基因的3 个启动子;spaR 启动子受到σH 转录因子的正向调节,在营养环境趋紧时σH 促进subtilin 基因簇的表达[8]。subtilin 基因簇的具体结构示意图见图2。

图2 Subtilin 基因簇的具体结构示意图
4 研究展望
近年来多种类抗生素在农业、畜牧业及医药领域被禁用,因而开发益生菌中不易产生抗性的Bacteriocins 具有重要的实际意义。B.subtilis细菌素主要抑制革兰氏阳性菌,在防治相关消化道细菌性疾病及食品贮藏中已有很多实际应用。通过基因组挖掘鉴定B.subtilis中新的抑制革兰氏阴性菌的细菌素或其他肽类代谢产物等具有抗菌性质的抗生素,以及利用基因工程手段将抑制革兰氏阴性菌的Bacteriocins 或来源于昆虫等的其它抗菌肽转入B.subtilis[20,21],将进一步拓展B.subtilis的应用,如用于抑制大肠杆菌(Escherichia coli)、沙门氏菌(Salmonella)等革兰氏阴性菌引起的腹泻等疾病[22]。
