基于太平洋甲胁虱裂化线粒体基因组推测甲胁虱属祖先线粒体核型
2022-02-15孙佳宁任天广董文鸽
孙佳宁, 任天广 , 陈 婷, 董文鸽,*
(1.大理大学病原与媒介生物研究所, 云南大理 671000; 2.大理大学护理学院, 云南大理 671000)
节肢动物线粒体基因组是双链闭合环状DNA分子,大小约在15~20 kb之间,共有37个基因,包括22个转运RNA(tRNA)基因、13个蛋白质编码基因(protein-coding genes, PCGs)、2个核糖体RNA(rRNA)基因和1个非编码区(又称为控制区或AT富含区)(Boore, 1999; Sharkey and Chapman, 2017)。吸虱昆虫的线粒体基因组发生裂化,形成多个线粒体微环染色体,这与绝大多数节肢动物极其保守的单染色体线粒体基因组形成鲜明对比。迄今为止,国内外对7科7属14种吸虱的线粒体基因组研究发现节肢动物典型的单染色体线粒体基因组在吸虱亚目(Anoplura)中发生了剧烈裂化现象,其线粒体微环染色体的数量以及每个线粒体微环染色体的基因结构和基因排序彼此不同,甚至在同属中亲缘关系较近的物种间也存在差异(Songetal., 2014)。迄今为止,完成测定的14种吸虱线粒体基因组都因裂化而由9~20个线粒体微环染色体组成,每个线粒体微环染色体的大小为1.5~4 kb,有1~8个基因,且不同种间的裂化速度和裂化程度有很大差异(Shaoetal., 2009, 2012, 2017; Jiangetal., 2013; Dongetal., 2014a, 2014b; Songetal., 2014; Herdetal., 2015; Fuetal., 2020)。这些吸虱线粒体基因在线粒体微环染色体上的分布有很大差异,即使是同属的物种,其线粒体基因组裂化模式也各不相同。
太平洋甲胁虱Hoplopleurapacifica隶属于节肢动物门(Arthropoda)昆虫纲(Insecta)虱目(Phthiraptera)吸虱亚目(Anoplura)甲胁虱科(Hoplopleuridae)甲胁虱属Hoplopleura,是我国云南黄胸鼠Rattustanezumi体表寄生的优势虱种(Guoetal., 2003)。甲胁虱属全世界有136种,是吸虱亚目中物种数最丰富的一个属(Durden and Musser, 1994),我国有22种(金大雄, 1999)。迄今为止,甲胁虱属中只有红姬甲胁虱Hoplopleuraakanezumi和克氏甲胁虱Hoplopleurakitti的线粒体基因组被测定(Dongetal., 2014a),但在这两种甲胁虱中都未找到完整的37个基因,在红姬甲胁虱只找到28个基因,有9个基因(nad1,nad3,nad5,trnF,trnG,trnH,trnL1,trnM和trnQ)未找到,而在克氏甲胁虱中只找到34个基因,有3个基因(nad5,trnF和trnH)未找到(Dongetal., 2014a)。
为了推测甲胁虱属祖先线粒体核型,本研究测定了太平洋甲胁虱的线粒体基因组,并与迄今为止测序的另外2种甲胁虱(红姬甲胁虱和克氏甲胁虱)的线粒体基因组进行了比较,推测甲胁虱属的祖先线粒体核型,以期进一步了解吸虱亚目是如何进化的。
1 材料与方法
1.1 太平洋甲胁虱与宿主采集
太平洋甲胁虱采自云南省金平县黄胸鼠体表,采集地分为室内(畜舍和人房)和室外(房周、灌丛、森林和耕地)两种不同的生境,共采集标本13头(8头雌成虫,5头雄成虫),将采集的太平洋甲胁虱标本置于盛有95%乙醇的EP小管内,并放置于-80℃超低温冰箱中保存备用,太平洋甲胁虱及其宿主黄胸鼠标本均保存于大理大学病原与媒介生物研究所。
1.2 太平洋甲胁虱DNA的提取
将1.1节采集的标本从盛有95%乙醇的EP小管内取出,置于S8APO体视显微镜下鉴别太平洋甲胁虱种类和性别。太平洋甲胁虱的主要特征在于腹部背面第1列的2根刚毛比第2列的短;除侧背片Ⅱ及侧背片Ⅵ的腹侧后叶突为尖状外,其余节Ⅲ~Ⅵ的各后叶突均为叶片状,后缘具较深的切刻。太平洋甲胁虱种类及性别的鉴定参考金大雄(1999年)编写的《中国吸虱的分类和检索》。将采集到的全部13头太平洋甲胁虱的标本用刀片在锡箔纸上切碎并分别放入1.5 mL的EP管中,再用DNeasy Tissue Kit(QIAGEN)试剂盒提取每头太平洋甲胁虱基因组DNA。
1.3 太平洋甲胁虱线粒体基因组测序与分析
以1.2节提取的基因组DNA为模板,参考吸虱亚目昆虫的通用引物(Shaoetal., 2017)(表1),PCR扩增太平洋甲胁虱的线粒体微环染色体rrnS和rrnL基因的短片段。PCR扩增反应体系(25 μL): 正反向引物(20 μmol/L)各0.5 μL, 模板DNA 1 μL, dNTP Mixture(2.5 mmol/L)2 μL, 10×Ex Taq Buffer(Mg+Plus)2.5 μL, Ex Taq(TaKaRa)0.25 μL, ddH2O 18.25 μL。PCR反应条件:94℃ 1 min; 98℃ 10 s, 45~48℃ 30 s, 72℃ 2~3 min, 30~40个循环; 72℃ 5 min。将获得的扩增产物送往上海生工生物工程技术服务有限公司进行Sanger双脱氧链终止法测序,获得rrnS和rrnL基因的部分序列。在rrnS和rrnL基因序列的保守区设计正反向特异性引物(表1),进行Long-PCR扩增,扩增出包含rrnS和rrnL基因的全长或近乎全长的大片段目的序列。PCR反应体系(25 μL):正反向引物(20 μmol/L)各0.5 μL, MgCl2(25 mmol/L)2.5 μL, dNTPs(各2.5 mmol/L)4 μL, 10×LA PCR BufferⅡ 2.5 μL, LA Taq(TaKaRa)(5 U/μL)0.5 μL, ddH2O 14.5 μL。反应条件: 94℃ 1 min; 98℃ 10 s, 45~65℃ 30 s, 68℃ 1~4 min, 30~40个循环; 72℃ 2~6 min。从rrnS和rrnL基因大片段目的序列中找出保守区序列后设计1对特异性引物(表1)扩增出太平洋甲胁虱全部线粒体微环染色体,大小为690~1 773 bp,扩增出的所有条带,仅包含太平洋甲胁虱所有线粒体微环染色体的编码区。特异性引物均通过OligoAnalyzer(https:∥sg.idtdna.com/site/account/login?returnurl=%2Fcalc%2Fanalyzer)网站设计。

表1 本研究所用的引物
扩增产物通过Wizard SV Gel and PCR Clean-up System(Promega)试剂盒纯化后送至美吉生物公司用Illumina HiSeq X Ten Platform进行高通量测序。测序结果用Geneious 2020.2.2(Kearseetal., 2012)软件组装。tRNA基因用tRNAscan-SE(Lowe and Eddy, 1997)和ARWEN(Laslett and Canbäck, 2008)查找,蛋白质编码基因和rRNA基因用BLAST(Gish and States, 1993; Altschuletal., 1997)查找。
1.4 系统发育分析
将本研究中获得的太平洋甲胁虱的线粒体基因组蛋白质编码基因序列与GenBank中7科7属14种吸虱和外群象虱Haematomyzuselephantis的线粒体基因组蛋白质编码基因序列结合构建吸虱的系统发育树,8科8属16种寄生虱(含外群象虱)共有的7个蛋白质编码基因(atp6,atp8,cox1,cox3,nad4,nad6和nad4L)序列通过Geneious 2020.2.2软件联配后用于系统发育分析,其余蛋白质编码基因(cob,cox2,nad1,nad2,nad3和nad5)序列并不适用于以上所有物种,因此在我们的分析中排除。蛋白质编码基因序列在TranslatorX(Federicoetal., 2010)在线平台(http:∥pc16141.mncn.csic.es/index_v4.html)用MAFFT算法基于氨基酸序列进行比对(Abascaletal., 2010)。使用IQ-TREE在线平台(Nguyenetal., 2015)(http:∥iqtree.cibiv.univie.ac.at/)进行最大似然法(maximum likelihood, ML)系统发育树的构建(Stamatakis, 2006),使用MEGA X软件构建邻接法(neighbor-joining, NJ)系统发育树。系统发育树节点支持率由自举检验值(bootstrap confidence level, BCL)进行评估,共计检测1 000次。
1.5 甲胁虱属祖先线粒体核型推断
用简约法(Shaoetal., 2017),根据甲胁虱属的3个物种太平洋甲胁虱、红姬甲胁虱和克氏甲胁虱并结合其他12种吸虱和象虱的线粒体基因组序列推断甲胁虱属祖先线粒体核型。我们将线粒体核型特征绘制到基于线粒体基因组序列构建的系统发育树上,以确定共同特征。我们推测一个线粒体微环染色体的核型特征是甲胁虱属的祖先特征,必须满足以下条件:1)其存在于至少1种甲胁虱中并且也存在于1个或多个不是甲胁虱属的吸虱中;或2)在3种甲胁虱中同时存在。由于tRNA基因在染色体上的移位比蛋白质编码基因和rRNA基因更活跃,因此我们将tRNA基因与rRNA基因和蛋白质编码基因分开考虑。若两个或多个线粒体基因组基因排序特征相互冲突,则把变化最少的基因排序特征推测为祖先特征,而其他基因排序特征则被拒绝。
2 结果
2.1 太平洋甲胁虱线粒体基因组的结构和基因组成
Illumina HiSeq X Ten Platform高通量测序获得太平洋甲胁虱线粒体基因组的优质序列2 150 795 reads(双端测序,序列读长250 bp),组装后获得太平洋甲胁虱线粒体基因组,共找到29个基因,包括11个蛋白质编码基因(atp6,atp8,cox1,cox3,nad1,nad2,nad3,nad4,nad4L,nad5和nad6),16个tRNA基因以及2个rRNA基因(rrnL和rrnS)。这29个基因不均匀地分布于10个线粒体微环染色体上(表2和图1)。每个线粒体微环染色体编码区有1~5个基因,编码区大小从C-nad6-W-L2的690 bp到H-nad5-F的1 773 bp。太平洋甲胁虱的10个线粒体微环染色体中有7个线粒体微环染色体具有1个蛋白质编码基因或1个rRNA基因,其余的3个线粒体微环染色体各有2个蛋白质编码基因。16个tRNA基因分布在10个线粒体微环染色体上(nad2线粒体微环染色体和rrnS线粒体微环染色体没有tRNA基因),每个线粒体微环染色体有1~3个tRNA基因。太平洋甲胁虱现有的29个基因,每个基因仅在一个线粒体微环染色体上出现,即不同线粒体微环染色体之间没有相同的基因。相对于非编码区,除了trnQ和nad1基因外,所有基因的转录方向一致(图1)。

图1 太平洋甲胁虱裂化线粒体基因组
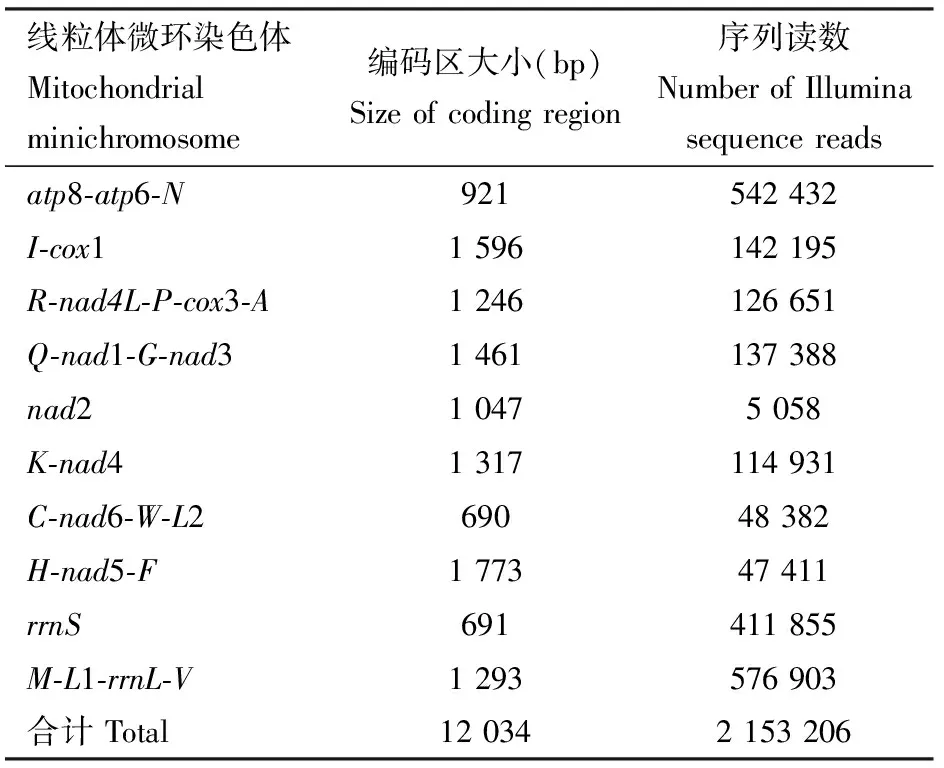
表2 太平洋甲胁虱线粒体微环染色体
2.2 同属内吸虱线粒体线粒体微环染色体组成差异
太平洋甲胁虱线粒体基因组裂化成10个线粒体微环染色体(未找到8个基因),在不考虑未找到的基因的情况下,太平洋甲胁虱与克氏甲胁虱有9个线粒体微环染色体一样,这9个线粒体微环染色体是:atp8-atp6-N,I-cox1,nad2,K-nad4,C-nad6-W-L2,rrnS,R-nad4L-P-cox3-A,Q-nad1-G-nad3和M-L1-rrnL-V(线粒体微环染色体用微环上的基因命名)。太平洋甲胁虱与红姬甲胁虱只有6个线粒体微环染色体一样,这6个线粒体微环染色体是:atp8-atp6-N,I-cox1,nad2,K-nad4,C-nad6-W-L2和rrnS。甲胁虱属的这3种吸虱差异主要涉及trnT基因。
2.3 系统发育
基于7科7属15种吸虱的7个线粒体基因组蛋白质编码基因序列构建的ML树(图2)和NJ树(图3),以象虱亚目(Rhynchophthirina)象虱的作为外群。除象虱外,其余寄生虱均为吸虱亚目。结果表明两种方法构建的分子系统发育进化树基本一致。进化树主要分为两大支:一个大的进化支包括甲胁虱科、多板虱科(Polyplacidae)、血虱科(Haematopinida)和微胸虱科(Microthoraciidae),另一个大的进化支包括虱科(Pediculidae)、阴虱科(Pthiridae)和猴虱科(Pedicinidae)。太平洋甲胁虱作为红姬甲胁虱和克氏甲胁虱的姊妹群,bootstrap为100,红姬甲胁虱与克氏甲胁虱关系更相近。甲胁虱科与多板虱科构成姊妹群,ML树中bootstrap为51,NJ树中bootstrap为42。

图2 最大似然法构建的基于线粒体基因组7个蛋白质编码基因序列的16种寄生虱的系统发育树
2.4 甲胁虱属祖先线粒体核型的推测
根据太平洋甲胁虱(本研究)、红姬甲胁虱和克氏甲胁虱,并结合其他12种吸虱和象虱的线粒体基因组数据推测甲胁虱属祖先线粒体核型(图4)。我们根据这些吸虱和象虱的线粒体基因组序列推断出系统发育树并绘制线粒体核型图,将线粒体核型特征定位于系统发育图上,以确定其共有特征(图5和6)。可推测出甲胁虱属祖先线粒体核型由12个线粒体微环染色体组成,每个线粒体微环染色体由一个编码区和一个非编码区构成,编码区包含1~6个基因(图4)。其中9个线粒体微环染色体编码区包括1个蛋白质编码基因或1个rRNA基因,其余3个线粒体微环染色体编码区包括2个蛋白质编码基因,分别为atp6和atp8、cox3和nad4L以及nad1(nad1与其他基因的转录方向相反)和nad3。除trnT基因外,其余基因均可推测出其在祖先线粒体核型中的位置,trnT基因的位置可能在cox2基因的下游或在trnA基因的下游。trnT基因紧靠非编码区的可能性较大,且其上游也有存在一小段非编码区的可能。截至目前仅3种甲胁虱的线粒体基因组数据并不能确定trnT基因在甲胁虱属祖先线粒体核型中的分布位置和排列方式,因此需要更多甲胁虱属的物种以更准确确定trnT基因位置。如果排除tRNA基因,这3种甲胁虱在12个线粒体微环染色体中蛋白质编码基因和rRNA基因的分布是相同的(图1和4)。

图4 推测的甲胁虱属祖先线粒体核型
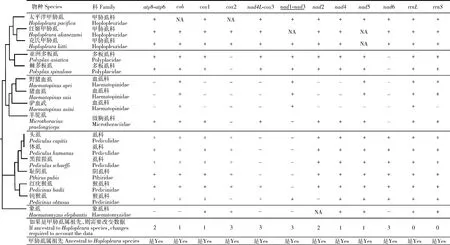
图5 基于线粒体基因组序列的系统发育树推测的甲胁虱属祖先线粒体核型(排除tRNA基因)

图6 基于线粒体基因组序列的系统发育树推测的与蛋白质编码基因和rRNA基因相关的tRNA基因在甲胁虱属祖先线粒体基因组中的位置
3 讨论
本研究对太平洋甲胁虱线粒体基因组进行测序与分析,共找到29个基因:16个tRNA基因,11个蛋白质编码基因(atp6,atp8,cox1,cox3,nad1,nad2,nad3,nad4,nad4L,nad5和nad6),以及2个rRNA基因(rrnL和rrnS)。29个基因不均匀地分布于10个线粒体微环染色体(表2; 图1)。每个线粒体微环染色体的编码区包含1~5个基因,编码区大小在690~1 773 bp之间。有8个基因(cob,cox2,trnD,trnE,trnS1,trnS2,trnT和trnY)未找到,然而这8个基因在人虱、猪虱、马虱、猴虱、黑猩猩虱、微胸虱和多板虱被发现,太平洋甲胁虱未找到的这8个基因在同属的克氏甲胁虱刚好是这2个线粒体微环染色体:trnE-cob-trnS1-trnS2环和trnD-trnY-cox2-trnT环(Dongetal., 2014b),这进一步说明甲胁虱属至少裂化成12个线粒体微环染色体。故推测以上8个线粒体基因缺失的可能性较小,未发现的原因更倾向于引物设计不保守造成。太平洋甲胁虱的rrnS基因单独存在一个微环染色体上,这是目前吸虱亚目中甲胁虱属吸虱特有现象。
分别将血虱属Haematopinus(3种)、虱属Pediculus(3种)、多板虱属Polyplax(2种)、猴虱属Pedicinus(2种)和甲胁虱属(3种)这5属内吸虱线粒体微环染色体的组成(Jiangetal., 2013; Songetal., 2014)进行比较,我们发现血虱属的猪血虱Haematopinussuis和野猪血虱Haematopinusapri线粒体微环染色体组成相同,而与同属的驴血虱Haematopinusasini线粒体微环染色体的组成有较大差异,这种差异主要涉及4个基因:trnR,trnM,nad6和nad4L;虱属的头虱Pediculuscapitis和体虱Pediculushumanus线粒体微环染色体组成相同,但与同属的黑猩猩虱Pediculusschaeffi线粒体微环染色体的组成有差异,这种差异主要涉及4个基因:trnE,trnM,trnN和trnS1(Shaoetal., 2012; Herdetal., 2015);多板虱属的棘多板虱Polyplaxspinulosa和亚洲多板虱Polyplaxasiatica线粒体微环染色体结构组成差异较大,主要涉及8个基因:trnA,trnF,trnL1,trnL2,trnP,trnS1,trnS2和trnT(Dongetal., 2014b);猴虱属的钝猴虱Pedicinusobtusus和红疣红虱Pedicinusbadii线粒体微环染色体结构组成差异最大,主要涉及11个基因:trnA,trnC,trnD,trnG,trnI,trnS2,trnV,trnW,nad2,nad3和rrnS(Fuetal., 2020)。由于甲胁虱属的克氏甲胁虱、红姬甲胁虱和太平洋甲胁虱均未能获得线粒体基因组全部基因,在不考虑未找到基因的情况下,甲胁虱属吸虱线粒体微环染色体结构组成差异仅表现在trnT基因。由此可以发现线粒体微环染色体组成差异体现在tRNA、与NADH脱氢酶亚基相关的蛋白质编码基因和rRNA基因,其中tRNA位置变化最为活跃,每属之间线粒体微环染色体的组成差异均涉及tRNA。
结合系统发育推测甲胁虱属祖先线粒体核型(图5和6),甲胁虱属祖先线粒体核型由12个线粒体微环染色体组成,每个线粒体微环染色体的编码区由1~6个基因构成(图4)。可以确定大部分基因的位置,trnT基因的位置目前不能确定,其可能在cox2或者trnA基因的下游位置,但可确定trnT基因在非编码区上游。将Fu等(2020)推测的宿主为灵长类动物吸虱的祖先线粒体核型(只能确定蛋白质编码基因和rRNA基因位置)与甲胁虱属祖先线粒体核型对比发现,蛋白质编码基因和rRNA基因在线粒体微环染色体中的位置完全一致。而Shao等(2017)推测的吸虱祖先线粒体核型,只有nad6基因的位置不同,吸虱祖先线粒体核型nad6与cox2基因在同一个线粒体微环染色体中,而另外两个祖先线粒体核型的nad6基因均单独在一个线粒体微环染色体中。由此可见在即使裂化线粒体基因组中,蛋白质编码基因和rRNA基因的位置也很稳定,tRNA基因流动性很强。
这些寄生虱的线粒体核型,即线粒体微环染色体的数量、以及每个线粒体微环染色体的基因组成和基因排列彼此不同,甚至在同属中亲缘关系较近的物种之间也不相同,这与大多数动物及其保守的单染色体线粒体基因组形成鲜明对比。本研究基于7科7属15种吸虱的蛋白质编码基因序列构建的进化树强烈支持吸虱亚目分为两大进化支,这与Light等(2010)根据18S rRNA,EF-1α和cox1基因推测49种吸虱的系统发育关系基本一致。在线粒体核型上的共有特征用简约法构建甲胁虱属的祖先线粒体核型。我们推测甲胁虱属的祖先线粒体核型特征时,若两个或多个线粒体基因组基因排序特征相互冲突,则把变化最少的基因排序特征推测为祖先特征,而其他基因排序特征则被拒绝。我们根据这些吸虱和象虱的线粒体基因组序列推断出系统发育树并绘制线粒体核型图,将线粒体核型特征定位于系统发育图上,以确定其共有特征(图5和6)。吸虱中2种人虱(头虱和体虱)、黑猩猩虱和啮齿动物体表寄生的5种吸虱(太平洋甲胁虱、红姬甲胁虱、克氏甲胁虱、棘多板虱和亚洲多板虱)谱系中多次发生线粒体微环染色体的裂化,而2种猪虱(野猪虱和猪虱)和马虱(驴血虱)谱系中线粒体微环染色体发生合并(Shaoetal., 2009, 2012, 2017; Jiangetal., 2013; Dongetal., 2014a, 2014b; Songetal., 2014; Herdetal., 2015)。线粒体微环染色体的裂化和合并共同创建了一个非常复杂动态的线粒体基因组结构。
