五爪金龙两种化感作用途径释放的化学成分分析及其效应研究
2022-02-15陈金慧马慧燕陈煜何禾
陈金慧,马慧燕,陈煜,何禾*
(1.海南大学生态与环境学院,海南 海口 570228;2.海南大学林学院,海南 海口5 70228;3.海南省农林环境过程与生态调控重点实验室,海南 海口 570228)
外来植物入侵严重威胁全球农业生产和生态系统的完整性,给生态环境造成了巨大破坏[1]。一直以来关于外来植物入侵机制的研究从未停止,外来植物能在到达新环境后强烈抑制本地植物甚至形成群落单一优势种的原因仍是生态学的难题之一[2]。新武器假说(new weapons hypothesis)认为,外来植物通过释放化学物质抑制本地植物的生长发育,从而增强在新生境的竞争能力,导致其成功入侵[3]。外来入侵植物通过淋溶、挥发、根系分泌及枯落物腐解等途径向环境中释放化学物质并对其他植物产生直接或间接有害作用的过程被称为化感作用[4],且不同物种释放的化感物质不同。研究发现薇甘菊(Mikania micrantha)分泌的阿魏酸、豆甾醇、α-香树酯醇达到一定浓度后能抑制萝卜(Raphanus sativus)及水稻(Oryza sativa)等幼苗的根长和苗高。硬脂酸甲酯和棕榈酸甲酯是南美蟛蜞菊(Sphagneticola trilobata)产生的最主要化感物质,对玉米(Zea mays)、黄瓜(Cucumissativus)的胚根及胚芽生长产生明显抑制[5-6]。深入了解某种外来植物的入侵机制,除关注其释放化感物质的种类外,还侧重于比较不同途径释放化感物质的差异,如胜红蓟(Ageratum conyzoides)和马缨丹(Lantana camara)以挥发释放为主[7-8],而根系分泌则是凤眼莲(Halerpestes cymbalaria)化感物质释放的最主要渠道[9],He等[10]已证明雨雾淋溶与枯落物分解才是尾叶桉(Eucalyptus urophylla)不可忽视的化感物质释放途径。由此可见,明确外来植物通过不同途径释放的化感物质种类及其各自的作用效果是深入了解其入侵机制的前提。
五爪金龙(Ipomoea cairica)为旋花科番薯属多年生缠绕草质藤本植物,作为广泛分布于我国华南地区的入侵性外来植物,它给当地自然生态系统带来了严重危害,也造成了巨大的社会经济损失[11]。为进一步防控五爪金龙扩散,其入侵能力与化感作用的关系已成为众多学者研究的重点。虽已有试验表明,五爪金龙浸提液对萝卜、白菜(Brassica pekinensis)、芥菜(Brassica juncea)、生菜(Lactuca sativa)、水稻等多种作物种子萌发和幼苗生长具有明显的抑制作用,且随着浓度的增大,抑制作用增强[12-18],但针对以淋溶水为代表的淋溶途径与以挥发物为代表的挥发途径释放的化感物质及其效应研究鲜见报道。
为明确不同释放途径在五爪金龙化感效应中所起的作用,并探讨通过不同途径释放的主要化感物质效果的差异,本研究首先通过模拟自然条件下五爪金龙挥发和淋溶两种途径的作用,比较两者对受体植物萌发影响的差异。再采用顶空固相微萃取和树脂洗脱法分别收集枝叶挥发物与淋溶水,以代表两种途径释放的化合物,并经气相色谱与质谱联用仪分离鉴定各自成分。接着根据定量得出两种途径释放化合物中最主要成分的实际浓度,采用种子萌发法,评价不同物质对受体种子萌发的影响。以鉴别不同途径释放的化感物质具体成分,并探讨不同化感物质特别是某些关键化合物对五爪金龙化感作用贡献的差异,从而通过五爪金龙对三叶鬼针草(Bidenspilosa)和生菜种子萌发的影响加深认知化感作用与五爪金龙入侵过程的联系,为治理其入侵危害提供理论依据。
1 材料与方法
1.1 植物材料
本试验选取生菜和三叶鬼针草作为受体植物,前者因发芽整齐且发芽率高,常被作为受体植物使用[19],后者则是南方常见与五爪金龙伴生的杂草之一。五爪金龙采自海口市近郊无人管理的抛荒区域,该区域以五爪金龙为单优物种,受体植物三叶鬼针草种子采自海南大学海甸校区校园内。生菜种子购于江西省丰城种业有限公司。选取大小均一,饱满度高的受体种子置于4℃冰箱保存备用。
1.2 淋溶水化学成分分析
为更好地了解在自然条件下通过雨雾淋溶途径释放化感物质的情况,本研究参照Song等[20]的方法,在海口市近郊无人管理的抛荒区域选取以五爪金龙为单优物种的样地,于叶片下方随机设置大小为1 m×1 m×0.3 m的PTFE采样盒,紧邻样地外的空地上放置同样大小采样盒作为对照,盒内容纳水量与样地所在区域降水量相当。2016年10月-2017年1月期间每隔1~3 d或在暴雨后测定收集的淋溶液体积,并于每两周将盒中收集的淋溶水取回实验室,4℃冰箱中保存备用。整个试验期间共收集野外淋溶水358.26 L,取回的淋溶水先分别经滤纸及水相滤膜(PALL,USA)过滤。层析柱内装填活化后的XAD-4树脂(Sigma-Aldrich,St.Louis,MO,USA),后将浸提液以150 mL·h-1的流速通过层析柱,再用10倍柱体积量的1∶1∶2∶1石油醚-乙酸乙酯-乙醇-水混合液对树脂进行洗脱,得到的洗脱液经0.45μm PTFE膜(Pall,USA)过滤,在旋转蒸发仪(Eyela,JPN)上浓缩蒸干。加入二氯甲烷溶解蒸干后的残留物质,并将溶液转移至试管。在残余干物质中加入500μL乙腈和500μL衍生化试剂(99%BSTFA+1%TMCS),70℃下反应1 h。待溶液冷却后经0.45μm的PTFE膜过滤后装入样品瓶,4℃冰箱中保存备用。
采用气相色谱质谱联用仪(GC-MS,Agilent7890B-5975C)对五爪金龙淋溶水成分进行分离与鉴定。色谱条件:弹性石英毛细管柱HP-5MS(30 m×0.25 mm×0.25µm);进样量1µL;不分流;进样口温度250℃;升温程序从50℃开始,以8℃·min-1的速度升温到70℃,保持2 min,然后以5℃·min-1的速度升温到90℃,保持3 min,最后以8℃·min-1的速度升温到220℃;载气(He)流速为1 mL·min-1;离子源为EI源;电离电压为70 eV;离子源温度为230℃;质量扫描范围为50~550 amu。
1.3 挥发物化学成分分析
为更好模拟野外条件下叶片挥发情况,根据野外调查已知本区域五爪金龙群落叶片密度7.5 kg·m-3,故称取7.5 g五爪金龙叶片置于1000 mL锥形瓶中,用封口膜将瓶口密封,将装有65μm PDMS/DVB萃取头的SPME装置插入锥形瓶于室温下进行吸附,整个过程持续30 min。每个样品吸附3次,重复上述过程3次。每次萃取后将萃取头置于250℃的GC进样口中热处理3 min以除去表面残留。另取同体积空白玻璃瓶作为对照,进行相同操作。
采样结束后,将SPME采样针插入GC-MS进样口解析5 min,对五爪金龙叶片挥发物进行分离与鉴定。除分流比5∶1以及柱升温程序变为初始温度80℃,以8℃·min-1的速度升温到160℃,保持2 min,之后以5℃·min-1的速度升温到240℃,其余检测条件与淋溶水样品测定相同。
1.4 模拟挥发与淋溶途径作用的生物测定
30粒生菜或三叶鬼针草种子均匀放置在铺有2层定性滤纸的密封瓶(3.5 cm×3.5 cm×12.5 cm)中,每个瓶中分别加入仅经过滤的野外采集淋溶水及对照组雨水4 mL,每处理设置5个重复。将培养瓶置于人工气候箱中,以相对湿度70%,温度为25℃,光照12 h,黑暗12 h培养。试验开始后每隔24 h观察一次,并适当添加水分保持盒内湿润,以胚根突破种皮视为萌发,每日记录发芽种子数量。
按照1.3中叶片与容器体积比,在15 L的密封培养盒底部放置新鲜五爪金龙叶片600 g,对照培养盒底部不放任何东西。将不加盖的培养皿放在培养盒中部的隔板上,并在培养皿底部铺两层经适量蒸馏水润湿后的滤纸,每皿放30粒受体植物种子。用封口膜缠绕瓶口保证密封彻底,每处理设置5个重复。放置于相对湿度为70%,温度为25℃的人工气候箱中,每日光照12 h,黑暗12 h。试验开始后每隔24 h观察1次,以胚根突破种皮视为萌发,每日记录发芽种子的数量。培养皿中定时补充蒸馏水保湿。6 d后统计发芽率和发芽速率。
1.5 主要成分的生物测定
采用标准品对1.2结果中相对含量超过5%的待测物质进行外标定量,再根据野外条件下收集的淋溶液总体积,得到其实际浓度。待测物质的标准品溶于适量蒸馏水中,得到与其实际浓度一致的化感物质溶液。将4 mL化感物质溶液滴入铺有2层定性滤纸的瓶(3.5 cm×3.5 cm×12.5 cm)中,30粒生菜或三叶鬼针草种子均匀放置,将培养瓶置于人工气候箱中,以相对湿度为70%,温度为25℃,光照12 h,黑暗12 h培养,每种处理设置5个重复。试验开始后每隔24 h观察1次,并适当添加化感物质溶液保持盒内湿润,以胚根突破种皮视为萌发,每日记录发芽种子数量。
采用标准品对1.3结果中相对含量超过5%的待测物质进行外标定量,再根据收集气态化合物容器总体积计算其实际释放浓度。根据其实际浓度进行生物测定,先将30粒生菜或三叶鬼针草种子均匀放置在铺有2层定性滤纸的瓶(3.5 cm×3.5 cm×12.5 cm)中,添加4 mL蒸馏水,滤纸中间放置一个敞口的PTFE小盒,盒中滴入适量待测物质纯品后迅速将瓶盖盖严,使物质均匀散开,最终瓶内物质挥发后的浓度与实际释放浓度一致。瓶口用封口膜缠绕以保证培养瓶密封彻底,每处理设置5个重复。放置于相对湿度70%,温度为25℃的人工气候箱中,每日光照12 h,黑暗12 h。试验开始后每隔24 h观察一次,以胚根突破种皮视为萌发,每日记录发芽种子的数量。培养皿中定时补充蒸馏水保湿。6 d后统计发芽率和发芽速率。
1.6 数据分析
发芽率(%)=最终发芽数/供试种子数×100%。
以修改后的Timson指数(T)表征发芽速率[21]:,第t天发芽率;t,第t天),Timson指数越大,发芽速率越快。
由Agilent chemstation软件分析GC-MS数据,通过仪器自带的NIST 2013标准谱库对物质进行初步定性,再应用科瓦茨保留指数(Kovats’retention index,RI)辅助定性。在前两种方法基础上,部分物质还采用标准品进行定性。采用峰面积计算得出物质相对含量。采用外标法对气态样品及液体进样进行定量,将标准品制成不同浓度梯度的标准溶液,在与样品相同的条件下检测,形成不同物质的标准曲线,在此基础上对样品中物质进行定量。
所有数据均为平均值±标准误;采用SPSS软件进行单因素方差分析,重复测量方差分析和显著性检验;采用Sigmaplot 10.0制图。
2 结果与分析
2.1 淋溶与挥发途径的生物测定
图1所示为第6天发芽率,结果表明五爪金龙枝叶淋溶水显著抑制了两受体植物种子发芽率,其处理组三叶鬼针草和生菜种子发芽率分别为55.6%和82.2%,比对照组分别降低31.4%和14.0%;五爪金龙叶片挥发物处理后的三叶鬼针草和生菜种子发芽率分别为33.3%和70.0%,比对照组降低了59.2%和25.8%,两者都受到了显著抑制作用。

图1 五爪金龙淋溶水(A)和挥发物(B)对三叶鬼针草和生菜种子发芽率的影响Fig.1 Effects of rain leachates and volatiles from I.cairica on the germination percentage of the seeds of B.pilosa and L.sativa
五爪金龙枝叶淋溶水和叶片挥发物总体延缓两种受体植物种子的发芽速率。经淋溶水处理后三叶鬼针草种子发芽速率从第3天开始极显著降低,生菜种子则从第2天开始极显著降低(P<0.01),经挥发物处理后,三叶鬼针草和生菜种子发芽速率分别从第2天和第1天开始极显著降低(P<0.01)。无论对照组还是处理组,两种受体植物发芽速率均随天数的增加而不断增大且同一处理下生菜发芽速率高于三叶鬼针草(图2)。

图2 五爪金龙淋溶水(A、B)和挥发物(C、D)对三叶鬼针草种子(A、C)和生菜种子(B、D)发芽速率的影响Fig.2 Effects of r ain leachates(A,B)and volatiles(C,D)fr om I.cairica on germination r ate of the seeds of B.pilosa(A,C)and L.sativa(B,D)
2.2 淋溶水与挥发物成分分析
五爪金龙枝叶淋溶水的检出成分总含量达99.95%(表1),其中酸类占比84.48%。酸类成分中肉桂酸的相对含量最高,达39.21%,此外,酸类成分还包括琥珀酸(15.15%),丙酸(11.12%),月桂酸(9.38%)等9种;淋溶水混合物中共检出4种酯类,相对含量占3.28%,以苯甲酸2-甲基苯基酯(1.67%)为代表;烷烃类共有6种,相对含量为8.98%;淋溶水中还检测出醇类、酚类和烯类物质各1种。五爪金龙枝叶淋溶水中相对含量较高的4种检出成分(肉桂酸,琥珀酸,丙酸,月桂酸)的绝对浓度分别为378.59,263.17,175.62和166.73μg·L-1。
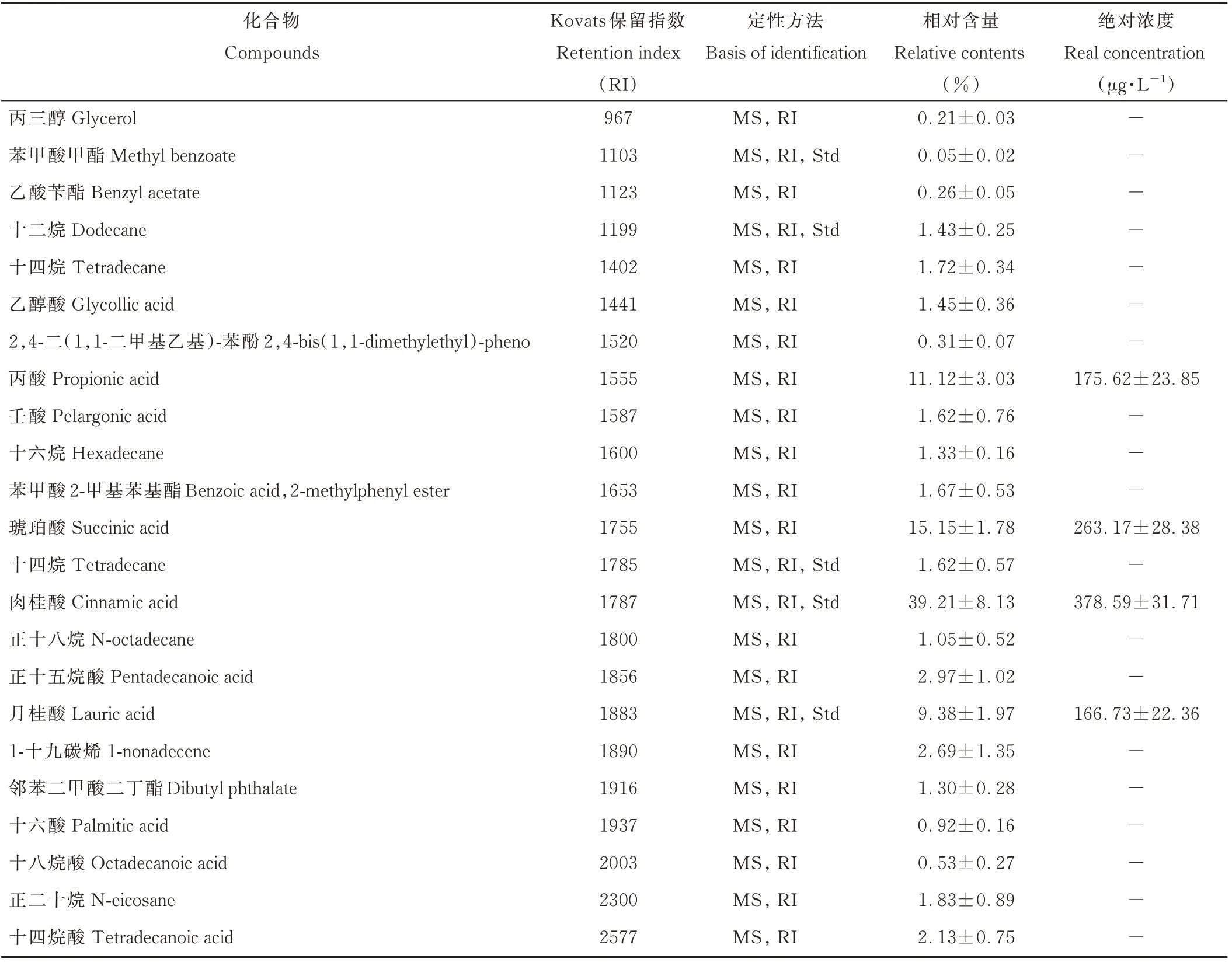
表1 五爪金龙淋溶水成分分析Table 1 Chemicals compounds of the r ain leachates from I.cairica
五爪金龙叶片挥发物中共分离鉴定出18种萜烯类化合物(表2),总相对含量达96.12%,其中石竹烯(28.17%),β-罗勒烯(16.59%),α-蒎烯(13.65%)和β-柏木烯(9.43%)的相对含量较高,其绝对浓度分别为3.62,1.86,1.77和1.20μg·L-1。其余含量大于1%的物质有11种,包括大根香叶烯(4.63%),α-古巴烯(3.08%)等。

表2 五爪金龙挥发物成分分析Table 2 Chemicals of the volatiles from I.cairica leaves
2.3 淋溶水主要成分生物测定
与对照组相比,经肉桂酸和月桂酸处理后的三叶鬼针草和生菜种子发芽率显著降低(P<0.05)且肉桂酸和月桂酸对两种受体植物种子发芽率的抑制作用无明显差异,但丙酸和琥珀酸处理组则未对受体植物种子发芽率产生明显影响。肉桂酸处理组三叶鬼针草和生菜种子发芽率分别为46.7%和83.3%,较对照组降低了43.9%和15.3%,月桂酸处理组三叶鬼针草和生菜种子发芽率则分别为47.8%和84.4%,较对照组降低了41.5%和14.3%(图3)。

图3 五爪金龙淋溶水4种主要成分对三叶鬼针草和生菜种子发芽率的影响Fig.3 Effects of main components of rain leachates fr om I.cairica on the ger mination rate of B.pilosa and L.sativa seeds
如表3所示,肉桂酸、丙酸、月桂酸总体抑制三叶鬼针草种子发芽速率,琥珀酸在第4和5天对三叶鬼针草发芽速率有促进作用。肉桂酸、丙酸、月桂酸3个处理组的三叶鬼针草种子发芽速率在第3天开始显著低于对照组(P<0.05),肉桂酸和月桂酸处理对发芽速率的抑制作用无显著差异且均明显强于丙酸;琥珀酸处理组则在第4和5天发芽速率显著高于对照组(P<0.05)。肉桂酸和月桂酸处理组第1天即开始显著抑制生菜种子发芽速率(P<0.05),且抑制作用无明显差异,琥珀酸和丙酸对生菜种子发芽速率作用不明显。不同处理组两种受体植物发芽速率均随天数增加而不断上升。

表3 五爪金龙淋溶水主要成分对三叶鬼针草和生菜种子Timson指数的影响Table 3 Effects of main components of r ain leachates from I.cairica on Timson index of the seeds of B.pilosa and L.sativa
2.4 挥发物主要成分生物测定
4种通过挥发途径释放的成分对两种受体植物种子第6天发芽率的影响不同。生菜发芽率无显著变化,三叶鬼针草经β-柏木烯和石竹烯处理后发芽率分别为53.3%和46.7%,较对照组显著降低,另外两种物质α-蒎烯和β-罗勒烯对三叶鬼针草发芽率则无显著影响(图4)。

图4 五爪金龙挥发物主要成分对三叶鬼针草和生菜种子发芽率的影响Fig.4 Effects of main components of volatiles from I.cairica on the germination rate of B.pilosa and L.sativa seeds
α-蒎烯和β-罗勒烯对三叶鬼针草和生菜发芽速率无显著影响,β-柏木烯和石竹烯前中期显著降低生菜发芽速率,自第3天开始显著降低三叶鬼针草发芽速率,二者的抑制作用无显著差异。不同处理组两受体植物发芽速率均随天数增加而不断上升(表4)。

表4 五爪金龙挥发物主要成分对三叶鬼针草和生菜种子Timson指数的影响Table 4 Effects of main components of volatiles fr om I.cairica on Timson index of the seeds of B.pilosa and L.sativa
3 讨论
3.1 两种化感物质释放途径作用效果
外来植物入侵机制是生态学研究的热点,越来越多的研究表明,产生化感物质作为“新武器”抑制本地植物生长发育促进了外来植物形成群落单一优势种[1],而挥发与淋溶是外来植物不可忽视的释放化感物质重要途径[22-23]。许多试验通过浸提液模拟自然淋 溶,如 李 澳 然 等[24]发 现 意 大 利 苍 耳(Xanthium strumarium)根、茎、叶和果实的水浸提液均极显著降低了油葵(Helianthus annuus)种子的萌发指数。关于植物挥发物的研究发现高等植物可通过茎、叶、花等地上部向外界释放挥发性物质进而影响其他植物生长发育,如土荆芥(Dysphania ambrosioides)挥发物具有较强的细胞毒性,使得蚕豆(Vicia faba),花生(Arachis hypogaea),荞麦(Fagopyrum esculentum)等多种作物保卫细胞产生核固缩、核降解、核错位等现象并危害作物的生长发育[25];胜红蓟茎叶挥发物对稗草(Echinochloa crusgalli),黑麦草(Lolium perenne)和三叶鬼针草的根长、苗高和鲜质量均有抑制作用[26]。本研究结果表明五爪金龙挥发和淋溶两种作用途径可显著抑制受体植物种子萌发,这与前人对外来入侵植物淋溶和挥发两种化感作用途径的研究具有一致性。
许多植物通过多种作用途径发挥化感作用,如黄花蒿(Artemisia annua)通过挥发和地上部淋溶共同作用向外界释放化感物质[27],节裂角茴香(Hypecoum leptocarpum)通过地上部挥发和土壤残体分解对黄瓜、萝卜、油菜(Brassica campetris)、小麦(Triticum aestivum)种子萌发和幼苗根长产生抑制作用[28];但不同植物可能有不同的主要化感作用途径,如胜红蓟和马缨丹以挥发作为化感物质进入周围环境的主要途径[7-8],窿 缘 桉(Eucalyptus exsert)和 尾 叶 桉(Eucalyptusurophylla)叶片中水溶性的化感物质通过淋溶途径产生较强的化感作用[3],造成此种差异的原因可能有两方面,一方面是植物自身产生的主要化感物质不同,其释放途径不同,有研究结果表明具有较强化感活性的酚酸类物质进入环境的主要方式之一是雨雾淋溶[29],而萜类物质在高等植物中多以挥发物形式存在[30];另一方面是所处环境不同,植物释放化感物质途径也不同,研究发现豚草(Ambrosia artemisiifolia)在降水较多时以淋溶为主要化感作用途径,在干旱高温条件下则主要通过挥发向周围环境释放化感物质[31]。通过对五爪金龙的研究可发现,挥发和淋溶两种途径都能抑制受体植物的生长发育,在自然界这两种途径可共同作用,这与黄花蒿具有一致性,但不同化感作用途径的效果和影响程度以及它们之间相互关系仍需进一步试验证明。
3.2 不同类型化感物质效应的差异
Rice[4]将高等植物化感物质分为含氮化合物类、萜类、酚类、聚合物类和香豆素类等,其中酚酸类和萜类化合物较为常见[23]。有学者指出酚酸类物质能改变细胞膜的透性,影响细胞对养分和水分的吸收[32-33],还能通过影响关键酶类,干预光合作用等抑制种子萌发和幼苗生长,例如咖啡酸、绿原酸和儿茶酚能通过抑制磷酸化酶进而抑制种子萌发[34];苜蓿(Medicago sativa)幼苗的根茎长在酚酸作用下会变短小[35];大豆(Glycine max)种子萌发,叶绿素积累,幼苗根茎长及鲜干重都受酚酸不同程度地抑制[36]。肉桂酸作为一类典型酚酸类物质,其单一施用会对生菜幼苗根茎长等生长指标产生明显的抑制作用,高浓度溶液会使根尖细胞失去活力[37],肉桂酸的施用还会降低西瓜(Citrulluslanatus)细胞内超氧化物歧化酶(super oxide dismutase,SOD)活性[38],目前关于月桂酸化感作用的研究报道很少,本研究结果表明其对三叶鬼针草和生菜种子萌发有明显的抑制作用。本研究在淋溶水中检测出较多以肉桂酸和月桂酸为代表的酚酸类物质,其对三叶鬼针草和生菜种子萌发有明显的抑制作用,与前人研究结果一致。
已有研究证明倍半萜是一类具有很强化感作用的萜类物质[39],南美蟛蜞菊中分离出的倍半萜内酯可影响花生种子酶活性,降低其呼吸速率[30],并对萝卜、番茄(Lycopersicon esculentum)等作物幼苗生长具有抑制作用[40],张晓芳[41]发现β-石竹烯对苘麻(Abutilon theophrasti)根长的抑制率可达30.6%;目前对β-柏木烯化感作用的相关研究较少,本研究结果表明其对受体植物有明显抑制作用。五爪金龙挥发物能显著抑制三叶鬼针草和生菜种子萌发,其中起显著抑制作用的物质是倍半萜类物质β-柏木烯和石竹烯,可能是因为其能对细胞膜功能和代谢过程产生影响,进而对受体植物起作用。
自然界中一种植物能合成多种化学物质,这些物质通过拮抗、加和、协同等方式共同发挥作用[39]。多种萜类物质共同施用对稗草的抑制率高于单种萜类物质施用[42];酚酸类物质中,肉桂酸和苯甲酸共同施用对大豆种子的发芽率、胚根伸长率和胚根干重的抑制作用强于单一物质施用[43];此外肉桂酸和香草酸[41],β-石竹烯和香芹烯等也具有协同作用[44]。五爪金龙通过不同途径产生的不同萜类物质和酚酸类物质是否存在上述的各种共同作用方式,有待进一步研究。
3.3 化感作用模拟的准确性
适当选择生物测定指标是评价化感作用的关键因素。种子萌发对物种天然更新十分重要[45],其变化会影响受体植物在群落中的多度及竞争力[46],因此选取发芽率作为评价植物化感作用敏感性指标[47]是化感作用生物测定的主要方法之一。此外发芽速率的降低会严重影响该植物在生态系统中的竞争能力[46],仅以发芽率为测定指标并不能很好地应对种子延迟发芽的状况[48],故本研究将发芽速率作为衡量化感作用强弱的另一指标,结果与前人研究具有一致性[46],五爪金龙两种作用途径对两种受体植物发芽率和发芽速率均有抑制作用。
有学者研究发现不同外来植物的化感作用对受体具有选择性[49]。前人研究发现外来入侵植物加拿大一枝黄花(Solidago canadensis)根部浸提液能显著抑制鸡眼草(Kummerow ia striata)种子萌发,但对萝卜、生菜种子萌发的影响不显著[50];薇甘菊水浸液对黄瓜幼苗的生长有明显抑制作用,对菜心(Brassica rapavar.chinensis)幼苗则有促进作用[41]。五爪金龙通过挥发释放的物质对两种受体植物的萌发影响不同,叶片通过挥发途径对三叶鬼针草发芽率的抑制作用强于生菜,推测此现象与萜类物质受体选择性有关;五爪金龙淋溶水对生菜种子发芽率的抑制作用弱于三叶鬼针草,可推测酚酸类物质对受体植物也具有选择性。五爪金龙释放的不同类型的化感物质对受体植物作用的强弱可反映其对受体植物强烈的选择性。
供体植物和受体植物生长期不同,化感作用效果不同。研究表明,加拿大蓬(Conyza canadensis)在主茎旺盛生长时期对白菜、萝卜等种子萌发和幼苗生长的化感作用强于其他时期,其水浸液对玉米不同生长发育时期的化感作用强弱排列顺序为:幼苗期>吐丝期>灌浆期>成熟期>种子[51];萌发阶段刺苍耳(Xanthium spinosum)水浸液对小白菜(Brassica chinensis)、莴苣(Lactuca sativa)、黑麦草的抑制作用在刺苍耳幼苗时期最强[52]。本试验选取五爪金龙作为供体植物,经研究发现其对两种受体植物种子萌发均有明显的抑制作用,五爪金龙在不同生长时期的化感作用是否不同有待进一步研究证明。
研究表明,植物化感作用具有浓度效应,即随着化感物质浓度增加,化感作用强度不断增加[7]。外来植物自然条件下产生化感物质的浓度与实验室设置的浓度不同,因而化感作用可能也不同,这一点可能导致研究结果与真实情况产生偏差。本试验收集了自然状态下五爪金龙的淋溶水和挥发物,且后期采用五爪金龙淋溶水和挥发物中具体物质绝对浓度进行生物测定,试验结果能较好模拟自然条件下五爪金龙对生菜和三叶鬼针草的化感作用,此外,还能初步确定化感作用较强的具体物质。据试验结果可推测在自然界中,五爪金龙通过雨雾淋溶会携带酚酸类等化感物质进入环境,同时其叶片产生的萜类气态挥发物不仅会直接作用于本地植物,也会经雨水溶解进入土壤二次影响本地植物的生长和发育。五爪金龙能产生石竹烯等萜类和肉桂酸等酚酸类化感物质,这可能是其抑制海南本地植物生长发育的重要原因。这与前人发现紫茎泽兰(Eupatorium adenophorum)产生萜类挥发物抑制杉木(Cunninghamia lanceolata)种子发芽,加拿大蓬产生酚酸类物质抑制莴苣种子萌发和幼苗生长[53-54]等研究一致。因此在五爪金龙防治过程中不仅要对其进行杀灭,还要注意残体移除,避免化感物质以挥发或雨水冲洗再次影响其他植物。现有的试验还无法确定其他化感作用途径的效果,也无法判断其他入侵机制在其成功占领新生境中的影响程度,以及这些化感物质中具体起作用的成分仍都有待后续试验进一步证明。
4 结论
五爪金龙挥发与淋溶两种作用途径可明显降低三叶鬼针草和生菜种子的发芽率,总体延缓两种受体植物发芽速率。通过对两种作用途径产生的化感物质进行定性与定量研究发现,五爪金龙淋溶水中的优势成分是肉桂酸、丙酸、琥珀酸和月桂酸,其中肉桂酸和月桂酸对两种受体植物发芽率与发芽速率的抑制作用更明显,所以这两种物质是五爪金龙通过淋溶释放的主要化感物质;作为五爪金龙挥发物主要成分,α-蒎烯、β-罗勒烯、β-柏木烯和石竹烯中,β-柏木烯和石竹烯能显著降低三叶鬼针草发芽率,且对两种受体植物发芽速率有延缓作用,但α-蒎烯,β-罗勒烯对其无明显影响,可见与其他成分相比,β-柏木烯和石竹烯是五爪金龙挥发途径的主要作用物质。
