根际细菌对番茄幼苗生长与Cd解毒机制研究
2022-02-14郭军康贾红磊刘文建
郭军康, 王 敏, 吕 欣, 魏 婷, 贾红磊, 刘 洵, 刘文建
(陕西科技大学 环境科学与工程学院, 陕西 西安 710021)
0 引言
工业和农业生产的快速发展导致土壤重金属污染急剧增加,治理和修复重金属污染是亟待解决的问题之一.镉(Cd)是农业环境中普遍存在的最严重的污染物之一[1].由于人为和工业活动、含磷肥料的施用、污水污泥、农药等,大量的Cd被输入到农田地区[2],而番茄作为重要的蔬菜作物,会从土壤中吸收养分和重金属[3],进而导致Cd 富集在果蔬作物的可食用部位.据报道,相当大比例的种植土壤受到高度Cd污染[4],生长在Cd污染环境中的植物经常会出现一系列严重的植物毒性,包括产生过量的活性氧(ROS)、光合作用和呼吸作用下降、养分和水分吸收不足、各种代谢途径紊乱,甚至植物死亡[5].
利用重金属富集性植物修复土壤重金属污染存在生物量小、生长速度慢、逆境下重金属耐受性差等问题[6].因此,借助根际细菌协同植物的新型修复策略逐渐被学者重视,成为重金属污染土壤的植物修复研究的重要方向之一.根际环境中的根际微生物在促进植物生长、提高植物生物量和土壤中重金属的生物有效性方面有着重要的作用[7],能够增强植物宿主对环境污染物和其他逆境的耐受性,从而有效地增强共生体系对环境的修复能力.
Han等[8]从受Cd和Pb污染的莴苣根际土壤中分离出两株产多胺细菌,其能够提高植物对Cd和Pb的抗性,促进生菜生长.Manzoor等[9]从超富集植物蜈蚣草根际土壤中分离出一株抗As细菌PG-12,发现其对多种金属具有耐受性,并且能够促进蜈蚣草的生长.Kamran等[10]从重金属污染土壤中分离出一株恶臭假单胞菌,发现接种PGPR促进了Cd超富集植物Eruca sativa的生长和对镉的吸收.Wei等[11]研究显示,野生豆类根部的根瘤菌土壤杆菌同时对铅、铜、镉、锌具有抗性,并且可促进植物生长.黄文[12]筛选出一株能产生物表面活性剂的根际细菌LKS06,其在促进龙葵生长的同时显著地提高了龙葵对镉的富集能力,根和地上部镉的总累积量最高的分别比对照增加了36.7%和42.4%,并且证实该菌株对多种重金属(Pb2+、Zn2+、Cu2+) 也有很高的耐受性.
在本研究中,选用一株从重金属污染的稻田土壤样本中分离出的细菌菌株GBR32,并以番茄幼苗为研究对象,研究接种GBR32菌株对番茄幼苗生长和Cd胁迫响应过程.为根际细菌在作物重金属胁迫响应与受污染农田安全利用领域提供新的知识.
1 实验部分
1.1 实验材料
本试验所用番茄品种为石红三号,购于新疆石河子蔬菜研究所番茄研究开发中心.
于广东省大宝山矿区附近被重金属污染的稻田土壤样本中分离出的寡养单胞菌属 (Stenotrophomonassp.)GBR32作为实验菌株,由本实验室前期保存.
1.2 实验方法
1.2.1 根际促生菌的培养
细菌菌株在LB固体培养基中生长并在28 ℃下孵育24~48 h,挑选单菌落后于液体培养基中振荡培养,28 ℃、180 rpm培养至OD600=1.0,此时有效菌浓度约为1.0×106cfu/mL,5 500 rpm离心10 min,收集菌体并用无菌水洗涤2次,再将菌体重悬于无菌水中并定容至500 mL备用[13],取相同体积的无菌水作为空白对照.
1.2.2 水培实验
番茄种子用15%的次氯酸钠消毒并均匀播种于蛭石盘中,置于人工植物培养室(昼26 ℃/夜20 ℃,昼夜时间为16 h/8 h,相对湿度60%).待幼苗长至“两叶一心”时将其转移至含1/4 Hoagland营养液的水培箱中培养两周,随后用1/2浓度的Hoagland培养液培养一周.选取长势一致的番茄幼苗进行接菌处理,将番茄幼苗根部浸泡于制备好的菌悬液中室温避光培养2 h,未接菌的用去离子水浸泡作为对照,3天后对番茄幼苗进行2 mg/L的Cd(CdCl2·2.5H2O)处理,在Cd胁迫处理一周后收样.
1.2.3 番茄生物量和Cd含量的测定
分开收集番茄幼苗的地上部和根部,根部在5.0 mmol/L 的乙二胺四乙酸二钠( EDTA-2Na) 溶液中浸泡15 min,去除根表面附着的Cd2+,并用去离子水冲洗数次,置于烘箱中烘干至恒重,之后分别对地上部和根部进行称重以确定其生物量[14].将样品研磨混匀,准确称取0.25 g植物样品进行消解,使用原子吸收光谱仪进行Cd含量的测定[15],每组处理设4个重复,同时做空白实验.
采用转运系数(Translocation factor)来评价Cd从根部输送到地上部的能力,其计算公式为:TF=番茄地上部分Cd含量(mg/kg)/根系部分Cd含量(mg/kg).
1.2.4 番茄丙二醛和抗氧化酶活性的测定
将新鲜植物样品 (0.5 g) 在 5 mL 磷酸缓冲液(50 mM,pH7.8)中彻底匀浆,离心后将上清液用于后续测试[16].丙二醛(malondialdehyde,MDA)含量的测定采用硫代巴比妥酸法(TBA)[17].超氧化物歧化酶(SOD)活性测定采用氮蓝四唑(NBT)比色法[18].过氧化物酶(POD)活性测定采用愈创木酚法[19].过氧化氢酶(CAT)活性采用紫外吸收法测定[20].
1.2.5 叶绿素含量的测定
采用95%乙醇提取法,待提取液制备完成后,以95%乙醇为空白,在波长665 nm、649 nm、470 nm下测定吸光度并计算叶绿素a、b和类胡萝卜素的含量[21].
1.2.6 数据分析
本实验使用Excel 2016、SPSS、Origin 8.5进行数据的统计处理、显著性分析与作图.
2 结果与讨论
2.1 接种GBR32对Cd胁迫下番茄生物量的影响
植物对重金属的耐性可以根据植物地上部以及根系生物量的变化作为指标[22],当重金属浓度过高时,会抑制植物幼苗的生长,植株叶片小而失绿,从而导致植物的生物量积累受抑制而降低.
由图1可知,Cd处理组番茄地上部和根部的生物量显著低于CK对照组,分别降低了53.4%和53.8%,表明2 mg/L的Cd胁迫会抑制番茄幼苗的生长.接种GBR32处理组地上部和根部的生物量显著低于CK,分别降低了31.9%和31.2%,说明接种GBR32细菌后,能够诱导植物的系统抗性(ISR)从而抵御对生物胁迫的抗性,使植物进入防御状态,防卫反应的激活使细胞和能量代谢转向于逆境响应,从而打破了正常的细胞发育功能,导致植物生长减缓,降低番茄植株的生物量.GBR32+Cd处理组的生物量与Cd处理相比分别增加了63.6%和98.6%,说明接种GBR32菌株能够显著缓解Cd胁迫引起的番茄生长抑制.

图1 不同处理对番茄生物量的影响
2.2 接种GBR32对Cd胁迫下番茄MDA含量的影响
逆境下植物器官往往会受到伤害,产生膜脂过氧化现象,MDA则是膜脂发生过氧化反应后形成的最终产物,其含量通常用于表示植物的氧化损伤程度[23].
如图2所示,较CK处理组相比,Cd胁迫下番茄地上部和根部的MDA含量分别显著增加了15.2%和115.9%.说明Cd胁迫能够诱导番茄幼苗体内活性氧积累,加剧膜脂过氧化,从而使细胞膜的完整性受到破坏.GBR32和GBR32+Cd处理组较CK处理组相比,地上部的MDA含量分别降低了29.5%和38.2%,说明接种GBR32菌株后,植物的诱导系统抗性被激活,从而引发植物对生物和非生物胁迫的增强抗性,缓解了GBR32和Cd对植物膜脂的氧化损伤.与Cd处理组相比,GBR32+Cd处理组使得番茄地上部和根部的MDA含量分别降低了46.4%和14.5%,说明接种GBR32菌株后番茄地上部和根部MDA含量均有一定的缓解作用.
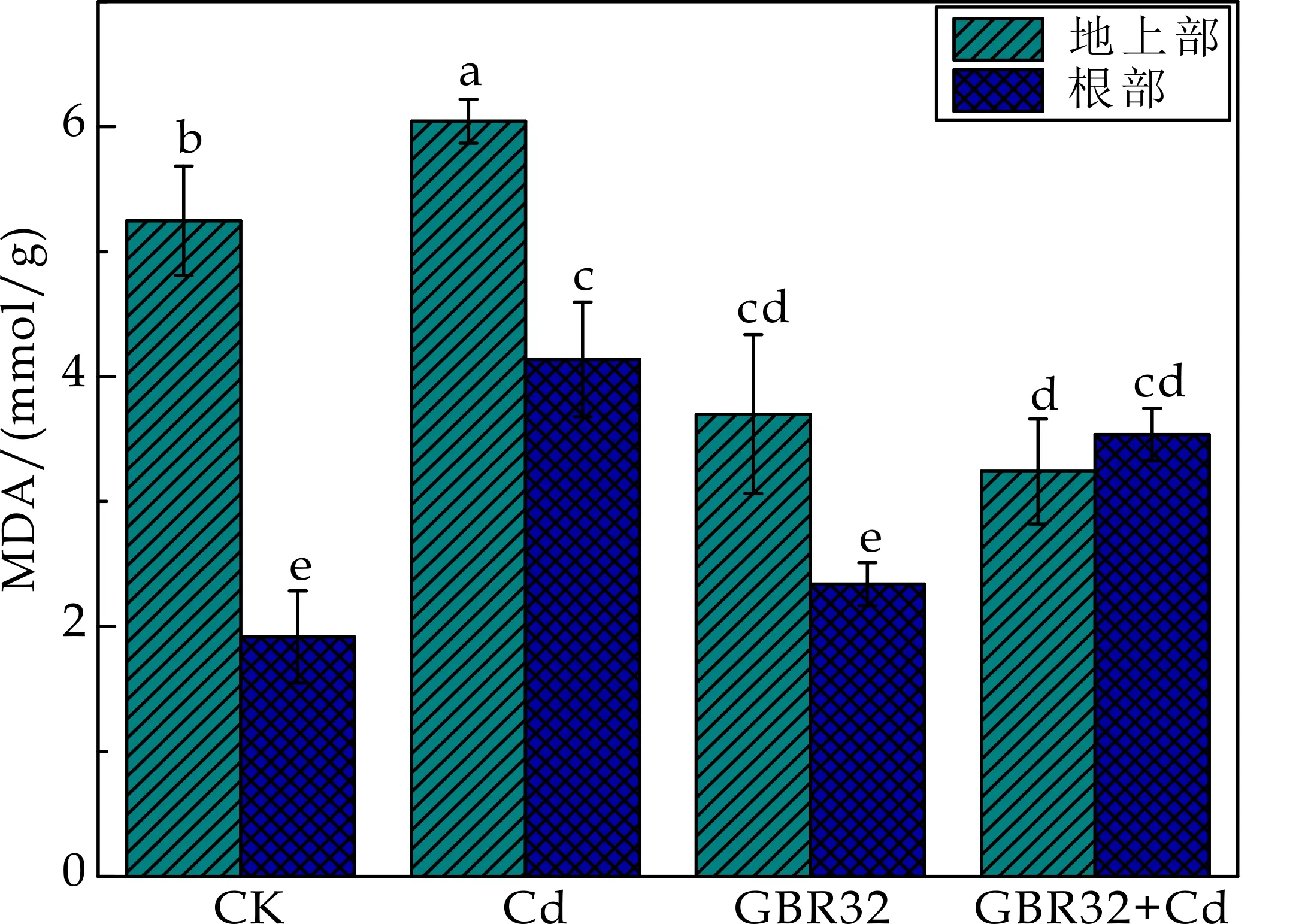
图2 不同处理对番茄MDA含量的影响
2.3 接种GBR32对Cd胁迫下番茄抗氧化酶活性的影响
在正常条件下,植物能有效地清除体内的活性氧,使细胞免受伤害.但在逆境条件下,植物体内的活性氧自由基产生速度超过了植物清除活性氧的能力[24].因此,为了减轻ROS造成的氧化损伤,植物体内已经进化出了一种由超氧化物歧化酶 (SOD)、过氧化物酶 (POD)、过氧化氢酶 (CAT)等构成的保护酶系统[25].
如图3所示,较CK处理组相比,Cd胁迫下番茄根部SOD活性提高了4.6%,说明番茄幼苗根系对Cd胁迫更为敏感,诱导SOD活性提高,从而清除植株体内的O2-.与Cd处理组相比,GBR32+Cd处理组番茄幼苗根部SOD活性降低了19.8%.这与MDA含量的结果相一致,说明GBR32菌株能够降低Cd胁迫下植株所受到的伤害程度,缓解氧化应激,因此SOD活性降低.

图3 不同处理对番茄SOD活性的影响
如图4所示,Cd胁迫下,番茄地上部和根部的POD活性较CK处理组相比分别提高了117.1%和29.5%.说明Cd胁迫下番茄植株为了清除体内过多的H2O2和其他过氧化物,从而诱导了POD活性的提高,以维持自身的正常代谢[26].与Cd处理组相比,GBR32+Cd处理组的地上部和根部的POD活性分别降低了62.9%和10.4%.这可能是由于接种GBR32菌株后,植株体内自由基积累量减少,对植株造成的氧化损伤程度减小,因此POD活性降低以调节植物的适应能力.

图4 不同处理对番茄POD活性的影响
2.4 接种GBR32对Cd胁迫下番茄叶绿素含量的影响
重金属通过干扰植物呼吸、破坏叶绿体的完整性、影响光合过程中的电子传递从而影响植物的光合作用[27].叶绿素含量的降低是植物在受到重金属毒害后而表现出来的现象.Pramanik等[28]研究表明,在Cd胁迫下接种根际促生菌显著提高了植物的叶绿素a、叶绿素b和总叶绿素含量(>2.5倍),从而缓解了Cd对植物光合作用所带来的毒害作用.
通过分析叶绿素含量发现(图5),与CK相比,Cd处理组的番茄叶片叶绿素a、叶绿素b和类胡萝卜素含量分别下降了41.1%、49.2%和35.3%,说明Cd能够抑制原叶绿素酸酯还原酶活性并阻碍氨基-γ-酮戊二酸的合成,从而影响叶绿素的生物合成[29],也可能与合成叶绿素所需的酶受Cd破坏有关[30].但与Cd处理相比,GBR32+Cd处理组的番茄叶片叶绿素a、叶绿素b和类胡萝卜素含量均有显著的增加,分别提高了20.5%、38.9%和14.1%.说明接种GBR32菌株有利于促进Cd胁迫环境下番茄幼苗叶绿素含量的增加,从而缓解Cd对番茄光合作用所带来的毒害.

图5 不同处理对番茄叶绿素含量的影响
2.5 接种GBR32对Cd胁迫下番茄内源茉莉酸含量的影响
茉莉酸(JA)作为重要的逆境信号分子在调控重金属胁迫等逆境胁迫中起着重要的作用.如图6所示,与CK对照组相比,Cd胁迫处理组番茄地上部和根部内源茉莉酸含量无显著变化,而接种GBR32菌株后,地上部和根部内源茉莉酸含量均有显著增加.与Cd处理组相比,GBR32+Cd处理组使得番茄地上部和根部茉莉酸含量分别显著增加了41.9%和9.3%.说明接种GBR32菌株可能通过诱导内源JA的合成从而激活JA/ET介导的诱导系统抗性.
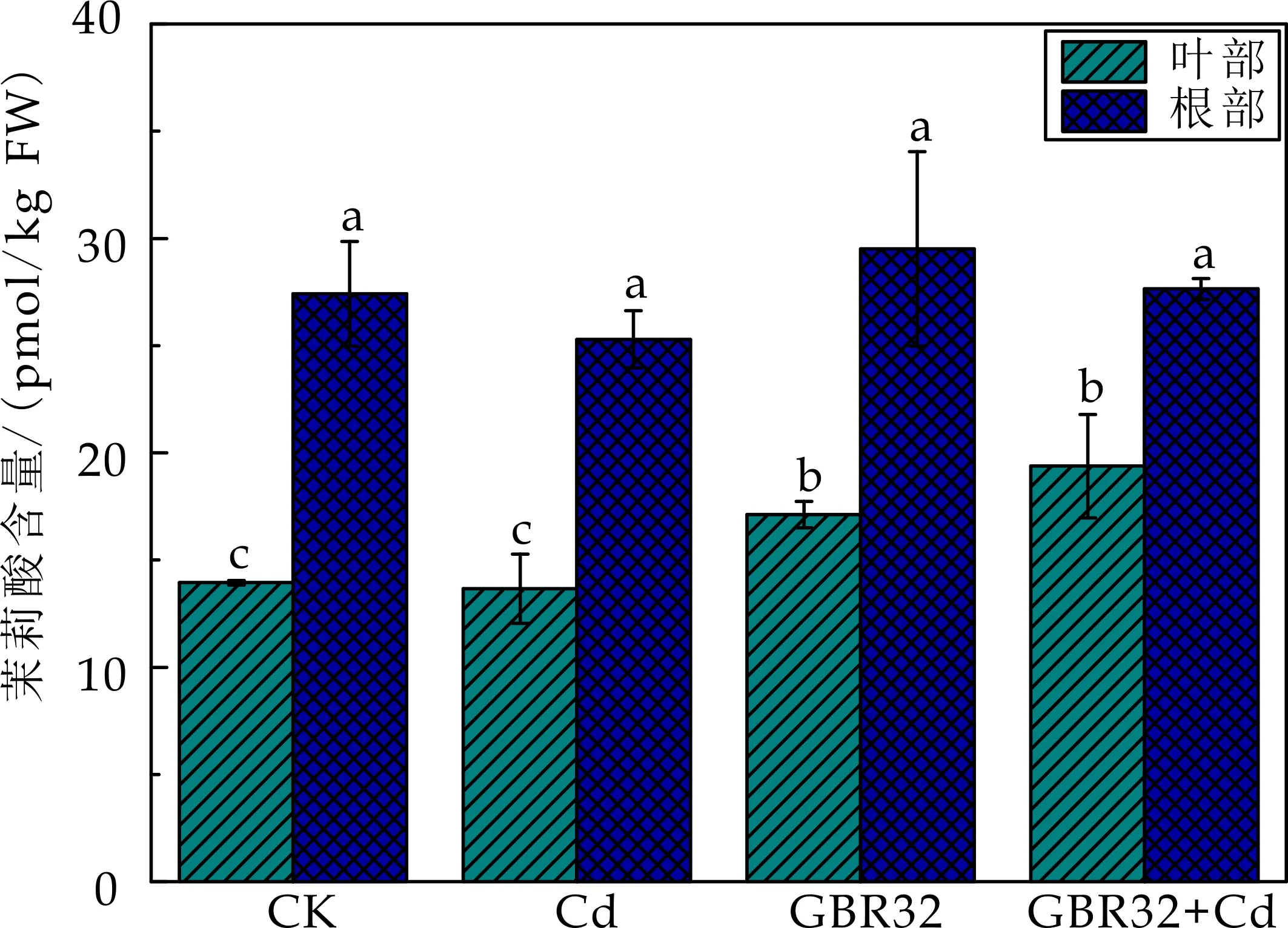
图6 不同处理对番茄JA含量的影响
2.6 接种GBR32对Cd胁迫下番茄内源乙烯含量的影响
植物在重金属胁迫下乙烯含量会增加,ET浓度的升高,会阻碍侧根的生长、根系伸长和根毛的形成,从而抑制植物生长[31].目前有益内生菌和根际促生菌的特点之一是可以产生1-氨基环丙烷-1-羧酸(ACC)脱氨酶,调控ACC是细菌协助植物抵抗非生物胁迫的重要途径之一[32].病原菌的入侵能导致细胞内ET的积累[33].
如图7所示,与CK处理组相比,Cd胁迫下番茄地上部的内源乙烯含量无显著变化,而根部的乙烯含量显著降低.与Cd处理组相比,GBR32+Cd处理组使得番茄地上部和根部乙烯含量分别显著增加了13.8%和11.8%.说明GBR32菌株可能通过乙烯含量的增加来诱导乙烯信号途径进而提高番茄对Cd的抗性.

图7 不同处理对番茄ET含量的影响
2.7 接种GBR32对Cd胁迫下番茄Cd累积量和转运系数的影响
重金属Cd存在于土壤中,并被植物吸收累积.研究表明,根际细菌能够调控不同植物对重金属的吸收[34].如图8所示,与Cd处理组相比,GBR32+Cd处理组使得番茄地上部和根部的Cd含量分别提高了46.7%和60.9%,显著增加了地上部和根部的Cd含量.
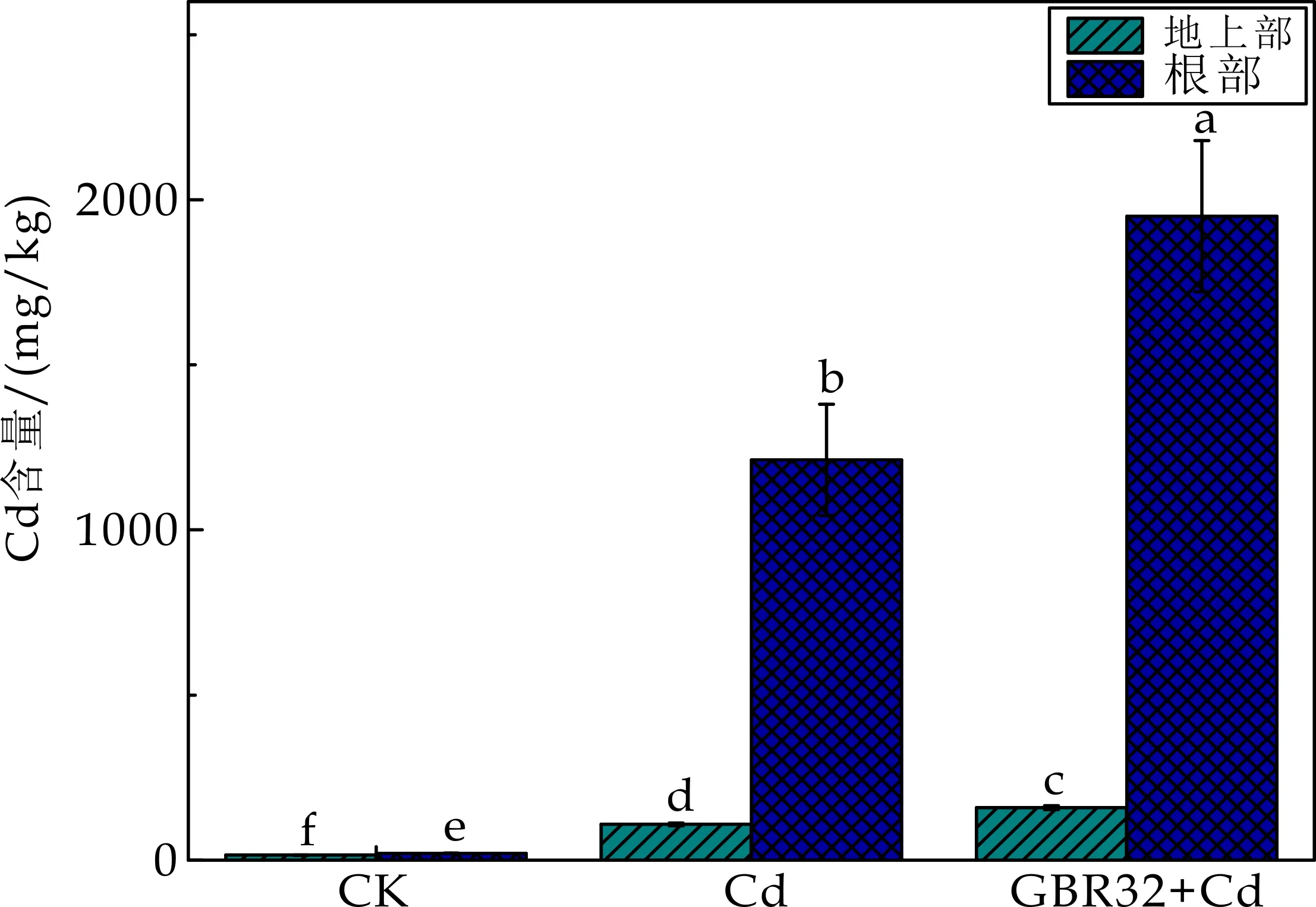
图8 不同处理对番茄Cd含量的影响
接种GBR32对番茄Cd的累积量的影响如表1所示.由表1可知,GBR32能够显著提高Cd胁迫下番茄根部Cd累积量.计算其转运系数结果如表1所示.由表1可以看出,在Cd胁迫下接种GBR32菌株能够降低Cd的转运系数,说明GBR32菌株能够抑制重金属Cd从根部向地上部的运输.综上,说明GBR32菌株能够大幅度提高番茄植株对Cd的吸收和富集能力,并且将Cd大量集中在根部.因此可以成为具有潜在重金属修复能力的菌株,进一步用于实际重金属污染修复[35].某些根际细菌细胞壁含有的羧基、羟基、氨基、磷酸基团等官能团能与金属离子配位络合,从而将其固定于细胞壁上[36],因此,细菌细胞壁组成与Cd累积之间的关系有待深入研究.
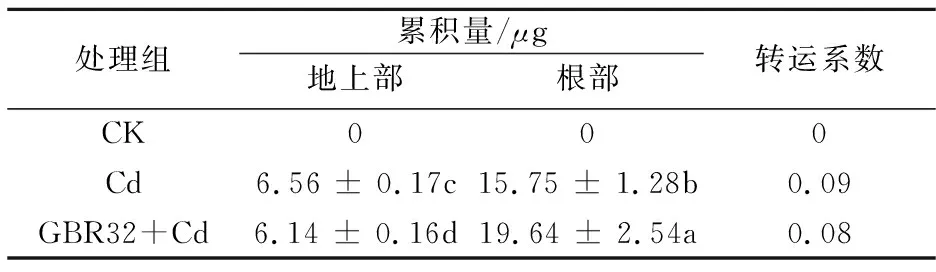
表1 不同处理下的Cd累积量
3 结论
本研究通过对番茄幼苗接种从被重金属污染的稻田土壤样本中分离得到的GBR32菌株,研究发现:(1)接种GBR32菌株可显著地提高Cd胁迫下番茄幼苗的生物量;(2)降低番茄MDA、SOD和POD活性,提高番茄叶片中的叶绿素含量,提高番茄内源茉莉酸和乙烯含量,缓解了Cd胁迫;(3)大幅度提高番茄根部对Cd的富集能力并将Cd固定在根部,降低Cd转运系数.该研究结果为根际细菌在调控作物重金属吸收转运,缓解重金属毒害,开展受污染农田安全利用提供科学依据.
