靶标基因突变导致入侵性杂草长芒苋对咪唑乙烟酸产生抗性
2022-02-10吉美静黄兆峰崔海兰陈景超于海燕李香菊
吉美静 黄兆峰 崔海兰 陈景超 于海燕 李香菊





摘要
長芒苋Amaranthus palmeri生长迅速,适应性广,繁殖系数高,具有很强的竞争性,已在我国多地定植,对作物产量及生态环境构成潜在威胁。一旦其对除草剂产生抗性,将大大增加治理难度。本试验研究了采自不同地点的长芒苋种群对除草剂咪唑乙烟酸的抗性水平和抗性机理。整株生物测定得出,长芒苋疑似抗性种群和敏感种群对咪唑乙烟酸的GR50分别为6 584.18 g/hm2和19.38 g/hm2, 抗性指数(RI)高达339.78。对抗性突变位点的分子检测发现,疑似抗性种群部分植株ALS基因574位色氨酸突变为亮氨酸(Trp574Leu),部分植株653位丝氨酸突变为天冬氨酸(Ser653Asp),除此之外,35%的植株还发生了Trp574Leu和Ser653Asp双位点突变。表明我国已存在对咪唑乙烟酸产生高水平抗性的长芒苋种群,主要抗性机理为ALS靶标位点突变。本研究首次在我国发现了长芒苋种群对除草剂咪唑乙烟酸的靶标抗性。而入侵性杂草对除草剂产生抗性也对其治理提出了新的挑战。
关键词
长芒苋;入侵性杂草;咪唑乙烟酸;抗性水平;ALS基因突变位点
中图分类号:
S482.4
文献标识码:A
DOI:10.16688/j.zwbh.2020639
Targetsite mutation in the ALS gene endowing imazethapyr resistance in invasive weed Amaranthus palmeri in China
JI Meijing#,HUANG Zhaofeng#,CUI Hailan,CHEN Jingchao,YU Haiyan,LI Xiangju*
(Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing100193, China)
Abstract
Palmer amaranth Amaranthus palmeri, an alien invasive species, has colonized in many sites all across China. It has rapid growing speed, wide adaptability and high reproduction coefficient with strong competitiveness, which poses a potential threat to crop yield and ecological environment. Once endowing resistance to herbicide, it will be more difficult for the weed control. The objective of this study was to determine whether A.palmeri populations collected from the two sites of China was imazethapyr resistant and to reveal the resistant mechanism to the herbicide. Results of wholeplant doseresponse experiments showed that the RI of the suspected resistant population was found to be 339.78fold more resistant to imazethapyr than a susceptible population based on biomass reduction (GR50values of 6 584.18 g/hm2 and 19.38 g/hm2 in suspected resistant population and the susceptible population, respectively). The ALS gene sequence analysis showed that in the suspected resistant populations, an amino acid substitution of position 574 (Trp574Leu) in some resistant plants and position 653 (Ser653Asp) in other resistant plants. Additionally, double mutation (both Trp574Leu and Ser653Asp) accounted for 35% in all tested plants. This research confirms that high level resistance of A.palmeri to ALSinhibiting herbicide imazethapyr with Trp574 to Leu and Ser653 to Asp amino acid substitution is the most likely mode of resistance. It is for the first report that A.palmeri population evolved target resistance to imazethapyr in China, and the resistance of invasive weeds to herbicides also posed a new challenge to the management of invasive weeds.
Key words
Amaranthus palmeri;invasive weed;imazethapyr;resistant level;ALS gene mutation site
长芒苋Amaranthus palmeri是一年生苋科Amaranthaceae苋属Amaranthus异株苋亚属Subgen Acnida L.杂草。原产于墨西哥西北部和美国的南加州至新墨西哥州到得克萨斯一带,后入侵至世界各地,现欧洲、大洋洲、亚洲等均有分布[1]。其植株生长迅速,适应性广泛,繁殖系数高,具有很强的竞争性。长芒苋种子萌发速度快,萌发对环境条件要求不严[2];其根系发达,株高可达2~3 m[3 -4],单株结实量达20万~60万粒[2,4];作为C4植物,它具有CO2补偿点低,蒸腾效率高的特点;且植株具有他感作用。因此,长芒苋能有效利用光能、水分、营养和空间[5 6],对生境中的其他植物产生生物胁迫影响[4]。目前长芒苋被美国列为棉田和大豆田10大难治杂草,也是最具破坏性和降低经济效益的抗草甘膦杂草之一。
1985年,我国首次在北京市丰台区南苑乡发现长芒苋[7-8]。目前,北京、天津、山东、江苏等地有长芒苋入侵报道,在耕地、荒地、河滩、沟渠边、村边、铁路与公路边、仓库周围、加工厂、工地、港口、垃圾场和饲养场周围等地长芒苋均可生长[7],并且有多地蔓延的趋势[9 -10]。中东部地区及华北地区,如北京、天津、河北、河南、山东中西部、山西南部、安徽北部和陕西中部等地为长芒苋潜在适生区[11]。与入侵地的反枝苋A.retroflexus、绿穗苋A.hybridus等其他苋属杂草相比,长芒苋植株长势和可塑性更高、对环境的适应能力更强,对水分、光照、营养的竞争也更加激烈[6],并易与本地植物种间杂交,把抗除草剂基因转移到近缘杂草[12]。对入侵地生境、农业生产及生物安全带来潜在隐患。
我们在前期的农田杂草抗性监测工作中发现,我国部分长芒苋种群对监测的靶标除草剂咪唑乙烟酸产生抗性。咪唑乙烟酸为咪唑啉酮类除草剂,具有除草效果好、使用剂量低、毒性小、使用方便等特点[13],一般用于大豆播前土壤处理或苗后茎叶处理,对长芒苋有较好的防治效果,但长期使用会导致长芒苋对该药剂产生抗性。国外有多例长芒苋对咪唑乙烟酸产生抗性的报道。1993年,美国的紫花苜蓿田、大豆田、玉米田、高粱田均发现对咪唑乙烟酸产生抗性的长芒苋[14]。近几年,阿根廷、巴西、以色列等国家也报道了抗ALS抑制剂的长芒苋种群[14 -15]。目前我国对长芒苋的研究主要集中在扩散范围[9-10]、分类鉴定[16]及适生性分析[17]等方面,尚无长芒苋对咪唑乙烟酸产生抗性研究的相关报道。
基于此,我们以咪唑乙烟酸为靶标除草剂,采用生物测定与分子生物学相结合的手段,对监测区域采集的长芒苋疑似抗性种群进行研究,以明确长芒苋对咪唑乙烟酸的抗性水平和抗性机理,为入侵性杂草长芒苋的有效治理提供支撑。
1材料与方法
1.1材料
杂草种子:长芒苋疑似抗性种群(种群1)采自山东省潍坊市,长芒苋敏感种群(种群2)采自北京市昌平区。
供试药剂及试剂:5%咪唑乙烟酸水剂(AS),山东先达农化股份有限公司;植物基因组DNA提取试剂盒、2x Pfu PCR Master Mix,天根生化科技(北京)有限公司;Trans2K DNA marker,北京全式金生物技术有限公司。
仪器設备:ASS4型行走式喷雾塔,Tee Jet8003扇形喷头,由国家农业信息化工程技术研究中心研制;PCR仪,型号AG22331,德国Eppendorf公司;低温离心机,型号Centrifuge 5417R,德国Eppendorf公司;全自动数码凝胶图像分析系统,型号Tanon1600,上海天能科技有限公司;电泳槽,型号JYC600C,北京君意东方电泳设备有限公司。
1.2试验方法
1.2.1长芒苋抗性水平研究
试验于2020年8月-10月进行,采用盆栽法培养长芒苋幼苗,自然光照,温度25~35℃。于直径9 cm的盆钵中分别种植疑似抗性种群(种群1)和敏感种群(种群2),每盆撒播20粒饱满一致的长芒苋种子,覆土1~2 cm。将盆钵放入装有水的托盘中,以盆钵底部渗灌方式供水,每隔3 d浇水一次,保持土壤湿度。幼苗长至2~3叶期间苗,每盆保留6株分布均匀、长势基本一致的长芒苋植株。于长芒苋3~4叶期,以ASS4型行走式喷雾塔喷施除草剂咪唑乙烟酸,喷雾压力0.275 MPa,喷液量367.5 L/hm2。根据前期预试验结果,种群1设8个处理,分别为75 g/hm2(1×,田间推荐剂量,按有效剂量计,下同)、150 g/hm2(2×)、300 g/hm2(4×)、600 g/hm2(8×)、 1 200 g/hm2(16×)、2 400 g/hm2(32×)、4 800 g/hm2(64×)和9 600 g/hm2(128×)。种群2设置6个处理,分别为9.375 g/hm2(1/8 ×)、18.75 g/hm2(1/4×)、37.5 g/hm2(1/2×)、75 g/hm2(1×)、150 g/hm2(2×)和300 g/hm2(4×)。每个处理设置3次重复。施药后21 d,剪取长芒苋地上部分,称量鲜重,计算相对鲜重。
1.2.2长芒苋抗性机理研究
根据长芒苋ALS基因序列(KY781923.1)设计引物(表1),扩增片段包括ALS基因保守区已报道的8个突变位点。其中反ALS1199f/r扩增片段长度为1 199 bp,包括574、653、654等3个位点;反ALS921f/r扩增片段长度为921 bp,包括122、197、205、376、377等5个位点,引物由北京六合华大基因科技有限公司合成。参照1.2.1幼苗培养方法种植长芒苋,待植株长到3~4叶期,分别剪取种群1和种群2单株的幼嫩叶片(每个种群20株样本),置于 80℃冰箱内保存。
采用植物基因组提取试剂盒提取长芒苋全基因组DNA。PCR扩增反应体系(25 μL):2× Pfu PCR Master Mix 12.5 μL,ddH2O 10.5 μL,上下游引物各0.5 μL,DNA模板1 μL。PCR程序:95℃预变性3 min;94℃变性30 s,53℃(ALS921f/r)/55℃(ALS1199f/r)退火30 s,72℃延伸2 min,35个循环;72℃延伸10 min。扩增产物于1%琼脂糖凝胶中进行电泳检测,并送北京擎科生物技术有限公司进行序列测定。利用Seq Man软件将测序后获得序列与GenBank长芒苋ALS基因序列(KY781923.1)进行比对,比较序列中8个被报道位点的差异性。
1.3数据处理与分析
采用SigmaPlot 12.5软件进行双逻辑非线性回归分析,得到剂量反应曲线、相关系数以及GR50,并求得抗性指数RI,具体公式如下。
双逻辑非线性回归模型:
Y=C+ D C 1+(X/GR50)b 。
Y为不同除草剂剂量处理下长芒苋的地上部分鲜重与对照地上部分鲜重的百分比;C为待测指标下限,D为待测指标上限;X为除草剂剂量;GR50为鲜重抑制中量,b为斜率。
相对鲜重计算公式:
相对鲜重= 处理杂草鲜重 对照杂草鲜重 ×100%;
抗性指数RI的计算公式:
RI= 抗性种群GR50 敏感种群GR50 。
2结果与分析
2.1长芒苋对咪唑乙烟酸的抗性水平
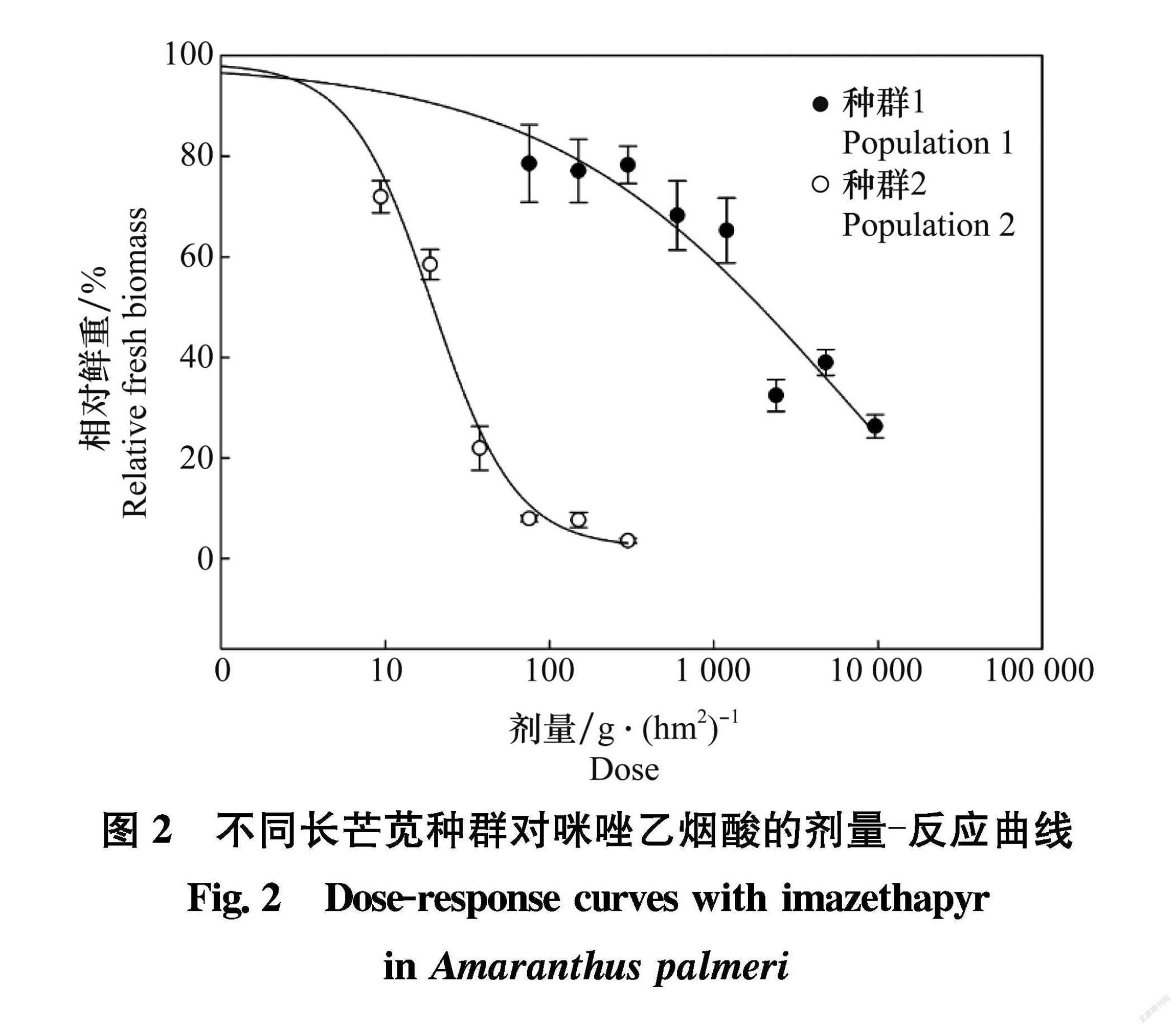
施用咪唑乙烟酸后,随着施药量逐渐增加,两个长芒苋种群的生长均受到不同程度的抑制(图1),主要表现为植株生长停滞,叶片黄化、植株逐渐枯死。施药后7 d,种群2在1/2×下,大部分植株死亡;而种群1在64×无植株死亡,128×处理,虽然抑制了植株生长,但仅有1~2株植株死亡。将施药剂量与药后21 d两个长芒苋种群地上部鲜重利用双逻辑非线性回归模型进行拟合,得到剂量 反应曲线(图2),相关系数分别为0.96、0.99。由此计算出种群1和种群2的GR50分别为6 584.18 g/hm2和19.38 g/hm,抗性指数(RI)为339.78。初步得出,长芒苋种群1对咪唑乙烟酸产生了高水平抗性,而长芒苋种群2对咪唑乙烟酸敏感。
2.2长芒苋对咪唑乙烟酸的抗性机理
上述2对引物扩增长芒苋ALS序列的部分片段结果如图3所示。通过片段长度可看出,图中条带为2对引物扩增的目标条带。将测序后长芒苋种群1、2的序列与GenBank中长芒苋ALS序列(KY781923.1)进行比对,结果发现种群1的574和653位点均发生突变(图4)。进一步分析得出,该种群基因突变类型为574位色氨酸(Trp)突变为亮氨酸(Leu),653位丝氨酸(Ser)突变为天冬氨酸(Asp);另外,在我们检测的20株长芒苋植株中,还发现有7株为Trp574Leu和Ser653Asp的双突变。种群2的ALS序列未发生突变。可见,靶标突变是长芒苋抗咪唑乙烟酸机制之一。
3讨论
研究结果表明,目前我国已经存在对咪唑乙烟酸产生抗性的长芒苋种群,并且为抗性水平较高的靶标抗性。这也是我国首次发现入侵性杂草对除草剂产生抗药性。推测种群1产生高水平抗性的原因有三种可能。第一种可能,长芒苋对除草剂的抗性基因系由我国本土归化苋属植物(如反枝苋)与长芒苋杂交引入。长芒苋为雌雄异株植物,易进行种间杂交。Molin等[12]通过杂交得到了长芒苋和刺苋Amaranthus spinosus的杂交种,并发现抗ALS除草剂基因的转移。陈金奕等[18]报道反枝苋对咪唑乙烟酸产生抗性,ALS基因存在3种突变类型,分别为205位丙氨酸被缬氨酸取代,653位丝氨酸被苏氨酸取代,574位色氨酸被亮氨酸取代。长芒苋种群1的突变位点与我国前期报道的对咪唑乙烟酸产生抗性的反枝苋突变有2个位点相同,这是否由反枝苋的抗ALS基因通过杂交转移到了长芒苋种群中需进一步验证。第二种可能,长芒苋入侵我国时便携带抗ALS除草剂基因。我国是大豆进口国,2019年,我国大豆进口8 850万t以上,主要来源于美国、巴西、阿根廷等国家。目前,美国、阿根廷、巴西等均发现抗ALS抑制剂长芒苋种群。Larran等[15]2017年在阿根廷首次发现了ALS基因发生突变的长芒苋,突变类型包括122位、574位和653位三种类型,其中对咪唑乙烟酸的抗性指数达206。Kupper等[19]报道了W574L和S653N两种突变类型的巴西长芒苋种群。2020年西班牙报道了抗ALS抑制劑的长芒苋生物型[20]。由于长芒苋入侵我国山东时间短,长期使用ALS抑制剂使其进化产生抗性的几率小,而进口大豆等粮食、货物夹带可能是抗性长芒苋的来源。第三种可能,长芒苋入侵我国后自然产生了靶基因突变,在ALS抑制剂的选择压下形成了抗性种群。
长芒苋作为外来入侵杂草,产生抗药性无疑会使我国生态环境和农业生产受到严重影响。资料报道,美国入侵性杂草造成的作物损失每年约为0.27亿美元[21]。与其他入侵杂草不同的是,长芒苋作为雌雄异株的植物,可以将抗除草剂基因转移到其他近缘杂草上。除此以外,长芒苋自身可以进行基因交换,也是对除草剂抗性增强的因素之一[22]。国外由于连续多年使用化学除草剂,使长芒苋产生了多种抗性生物型。如美国内布拉斯加州报道长芒苋对HPPD抑制剂(硝磺草酮、苯唑草酮、环磺酮)和光系统Ⅱ抑制剂(莠去津)产生抗性[23]。堪萨斯州发现了全球首例对草甘膦、氯磺隆、硝磺草酮、莠去津产生多抗性的长芒苋种群[24]。阿根廷报道了长芒苋对三类ALS抑制剂(氯嘧磺隆、双氟磺草胺、咪唑乙烟酸)产生了交互抗性[15]。巴西报道了对EPSPS抑制剂(草甘膦)、ALS抑制剂(氯磺隆、甲嘧磺隆、咪唑乙烟酸)产生多抗性的长芒苋[19]。因此,为了降低长芒苋在我国抗性遗传进化以及扩散的风险,在其治理技术上,应于幼苗期至植株结实期前进行人工拔除或机械铲除,防止种子成熟后扩散蔓延;加强境内外植物检疫,杜绝长芒苋种子随农副产品调运传入新的区域;除此之外,在使用化学除草剂治理长芒苋时,应坚持多种除草剂科学混用及轮换使用的原则。
杂草抗性机理主要包括两类,一类为靶标突变,一类为非靶标突变。其中靶标突变由于靶标基因位点发生突变或是靶标酶过量表达[25]。 非靶标突变则是由于杂草对除草剂的吸收代谢能力增强,使到达作用靶点的除草剂剂量降低,最终导致杂草抗药性的产生。本研究明确了我国入侵性杂草长芒苋对咪唑乙烟酸的抗性水平及主要抗性机理,发现我国长芒苋对咪唑乙烟酸产生高水平抗性原因之一是ALS基因发生靶标突变,至于上述种群是否存在ALS抑制剂非靶标抗性有待进一步探讨。
参考文献
[1]WARD S M, WEBSTER T M, STECKEL L E, et al. Palmer amaranth (Amaranthus palmeri): a review [J]. Weed Technology, 2013, 27(1): 12- 27.
[2]KEELEY P E, CARTER C H, THULLEN R J. Influence of planting date on growth of palmer amaranth (Amaranthus palmeri) [J]. Weed Science, 1987, 35(2): 199 -204.
[3]BRYSON C T, DE FELICE M S. Weeds of the south [M]. Athens: The University of Georgia Press, 2009.
[4]KILNGAMAN T E, OLIVER L R. Palmer amaranth (Amaranthus palmeri) interference in soybeans (Glycine max) [J]. Weed Science, 1994, 42(4): 523- 527.
[5]EHLERINGER J. Ecophysiology of Amaranthus palmeri, a sonoran desert summer annual [J]. Oecologia, 1983, 57: 107- 112.
[6]HORAK M J, LOUGHIN T M. Growth analysis of four amaranthus species [J]. Weed Science, 2000, 48(3): 347 -355.
[7]李振宇. 长芒苋—中国苋属一新归化种[J]. 植物学通报, 2003, 20(6): 734- 735.
[8]车晋滇. 外来入侵杂草长芒苋[J]. 杂草科学, 2008, 26(1): 58- 60.
[9]马金双. 中国入侵植物名录[M]. 北京: 高等教育出版社, 2013.
[10] 莫训强, 孟伟庆, 李洪远. 天津3种外来植物新记录—长芒苋、瘤梗甘薯和钻叶紫菀[J]. 天津师范大学学报(自然科学版), 2017, 37(2): 36 -38.
[11] 李慧琪, 赵力, 祝培文,等. 入侵植物长芒苋在中国的潜在分布[J]. 天津师范大学学报(自然科学版), 2015, 35(4): 57 -61.
[12] MOLIN W T, NANDULA V K, WRIGHT A A, et al. Transfer and expression of ALS inhibitor resistance from palmer amaranth (Amaranthus palmeri) to an A.spinosus×A.palmeri hybrid [J]. Weed Science, 2016, 64(2): 240 -247.
[13] 宋慧, 李海峰, 付楠,等. 咪唑啉酮类除草剂及其抗性作物的发展现状[J]. 河北农业科学, 2018, 22(3): 38- 41.
[14] HEAP I. International survey of herbicide resistant weeds [EB/OL]. [2020- 11-29]. http:∥www.weedscience.org∥Home.aspx.
[15] LARRAN A S, PALMIER V E, PEROTTI V E. Targetsite resistance to acetolactate synthase (ALS)inhibiting herbicides in Amaranthus palmeri from Argentina [J]. Pest Management Science, 2017, 73(12): 2578 -2584.
[16] 徐晗, 趙彩云, 刘勇波,等. ITS序列及其SNP位点在外来入侵杂草长芒苋、西部苋和糙果苋物种鉴定中的应用[J]. 植物保护, 2017, 43(5): 128- 133.
[17] 曹晶晶, 王瑞, 李永革,等. 外来入侵植物长芒苋在中国不同地区的表型变异与环境适应性[J]. 植物检疫, 2020, 34(3): 25- 31.
[18] 陈金奕, 张朝贤, 黄红娟,等. 反枝苋对咪唑乙烟酸抗性水平及分子机制[J]. 植物保护, 2015, 41(2): 130- 134.
[19] KUPPER A, BORGATO E A, PATTERSON E L, et al. Multiple resistance to glyphosate and acetolactate synthase inhibitors in palmer amaranth (Amaranthus palmeri) identified in Brazil [J]. Weed Science, 2017, 65(3): 317- 326.
[20] TORRA J, ROYOESNAL A, ROMANO Y, et al. Amaranthus palmeri a new invasive weed in Spain with herbicide resistant biotypes [J]. Agronomy. 2020, 10(7): 933.
[21] PIMENTEL D, ZUNIGA R, MORRISON D. Update on the environmental and economic costs associated with alieninvasive species in the United States [J]. Ecological Economics, 2005, 52(3): 273 -288.
[22] CHAUDHARI S, JORDAN D L, YORK A C, et al. Biology and management of glyphosateresistant and glyphosatesusceptible palmer amaranth (Amaranthus palmeri) phenotypes from a segregating population [J]. Weed Science, 2017, 65(6):755- 768.
[23] CHAHAL P S, GANIE Z A, JHALA A J. Overlapping residual herbicides for control of photosystem (PS) Ⅱand 4hydroxyphenylpyruvate dioxygenase (HPPD)inhibitorresistant palmer amaranth (Amaranthus palmeri S. Watson) in glyphosateresistant maize [J/OL]. Frontiers in Plant Science, 2018, 8: 2231. DOI:10.3389/fpls.2017.02231.
[24] KUMAR V, LIU Rui, BOYER G, et al. Confirmation of 2,4D resistance and identification of multiple resistance in a Kansas palmer amaranth (Amaranthus palmeri) population [J]. Pest Management Science, 2019, 75(11): 2925 -2933.
[25] POWLES S B, YU Qin. Evolution in action: plants resistant to herbicides [J]. Annual Review of Plant Biology, 2010, 61: 317- 347.
(責任编辑: 田喆)
收稿日期:2020 -11- 29修订日期:2020- 12- 07
基金项目:
国家现代农业产业技术体系(CARS-25)
* 通信作者
Email:xjli@ippcaas.cn
