中国野生毛葡萄转录因子VqWRKY6与VqbZIP1互作调控抗白粉病功能分析
2022-02-02张洁姜长岳王跃进
张洁,姜长岳,王跃进
中国野生毛葡萄转录因子VqWRKY6与VqbZIP1互作调控抗白粉病功能分析
张洁,姜长岳,王跃进
西北农林科技大学园艺学院/旱区作物逆境生物学国家重点实验室/农业农村部西北地区园艺作物生物与种质创制重点实验室,陕西杨凌 712100
【目的】欧洲葡萄作为世界葡萄主栽品种,具有产量高、品质佳的优点,但对病害抵抗能力差。白粉病是严重危害葡萄栽培的一种真菌性病害,中国野生葡萄资源丰富,可为抗病育种提供充足种质资源。论文旨在筛选调控抗白粉病的葡萄转录因子基因,探究转录因子基因调控抗白粉病的作用机理,为选育葡萄抗病品种提供优质的基因资源。【方法】从中国野生毛葡萄‘商-24’中克隆得到转录因子基因,使用DANMAN和MEGA-X软件对序列进行分析。利用PEG介导转化拟南芥原生质体进行亚细胞定位分析发挥转录调控作用的位置,利用酵母双杂交和双分子荧光互补试验证明VqWRKY6能够和转录因子VqbZIP1互作形成转录复合体。以感病葡萄‘赤霞珠’叶片为试材,通过农杆菌介导法瞬时转化到‘赤霞珠’葡萄叶片,叶片进行白粉菌()接种后,观察发病症状,用台盼蓝染色在显微镜下观察菌丝发育进程,使用DAB染色观察活性氧积累,比较共同过表达和的葡萄叶片、单独过表达的葡萄叶片、单独过表达的葡萄叶片和对照组叶片的差异。使用实时荧光定量PCR技术对抗病基因在白粉菌诱导下的表达水平进行分析。【结果】定位于葡萄2号染色体,编码342个氨基酸,属于WRKY家族的group Ⅲ亚家族。亚细胞定位和酵母转录激活试验证明,VqWRKY6在细胞核内发挥转录调控功能且在酵母中有转录激活活性。‘赤霞珠’叶片共同过表达和后,叶片表面白粉菌菌丝扩繁速率显著慢于单独过表达和单独过表达的叶片,共同过表达和的叶片组织中活性氧含量显著高于单独过表达和单独过表达的叶片;此外,VqWRKY6和VqbZIP1的协同调控能够激活茉莉酸(JA)途径的,基因表达量显著上调。【结论】VqWRKY6和VqbZIP1协同作用可能通过激活JA抗病途径,促进活性氧产生,增强抗病基因表达,抑制白粉菌的生长,从而提高葡萄对白粉病的抗性。因此,中国野生毛葡萄‘商-24’是重要的抗病种质资源,而与可作为重要的抗病基因资源。
中国野生毛葡萄;WRKY转录因子;白粉病;转录调控
0 引言
【研究意义】葡萄是世界上栽培历史悠久且重要的果树之一。根据联合国粮食及农业组织(FAO)的统计数据,2020年我国葡萄栽培面积和产量分别为76.75万公顷和1 484.31万吨,分别占世界葡萄栽培总面积和总产量的11%和19%。目前世界范围内广泛栽培的葡萄品种为欧洲葡萄,其具有品质优良、产量高的特点,但抵抗真菌病害的能力较差[1]。白粉病是葡萄栽培生产上危害较大的一种典型真菌病害,主要危害绿色幼嫩组织,受害组织表面生成白色粉状物,影响葡萄果实的正常发育并造成减产,严重损害葡萄的品质和产量[2]。在葡萄的栽培过程中普遍使用化学防治作为病害防治的手段。然而,喷施农药不仅增加葡萄栽培的成本,也会导致农药及其副产品的积累,对环境和人畜造成一定影响[3-5]。因此,选育抗病葡萄品种是控制病害产生的一种有效措施。中国是世界葡萄起源中心之一,野生抗病葡萄资源丰富[6]。这些野生资源蕴含丰富的抗病调控基因,为揭示葡萄与白粉菌()的互作机理提供了丰富的材料[7]。明确中国野生葡萄中存在的抗病调控基因,阐明其调控抗病性的作用机理,对抗病葡萄选育具有重要意义。【前人研究进展】WRKY转录因子在植物抗病信号途径中发挥重要的转录调控作用[8-11]。目前,在模式植物拟南芥中鉴定到WRKY家族成员75个[12],小麦中鉴定到270个[13],小豆和绿豆中分别有84个和85个[14],水稻中有83个[15],葡萄中有59个[16]。在拟南芥中异源表达毛葡萄‘丹凤-2’的可激活水杨酸(salicylic acid,SA)途径,增强植物的抗病性,VqWRKY53与MYB转录因子MYB14和MYB15的协同作用能够调控芪合酶基因的表达[17-18]。在转和葡萄植株中,‘无核白’对于白粉病抗性显著提高[19]。拟南芥中、和能够增强对丁香假单胞菌()抗性[20],增强对灰霉病抗性[21]。在大麦中,已发现有12个能够响应大麦白粉菌(f. sp.)的诱导,其中10个显著上调表达,2个明显下调[22]。水稻中过表达会增强对稻瘟病的抗性[17]。菊花中过表达能够增强植株对白色锈病的抗性[23]。此外,在葡萄中过表达能够增强对白粉病的抗性[24]。植物受到病原微生物的诱导会产生病程相关蛋白(pathogenesis-related protein,PR),植物抗病性的标志之一即PR蛋白的产生[25-26]。在易感葡萄叶片中过表达可以增强对霜霉病的抗性[27]。拟南芥中过表达,可以正向调控对白粉病的防御反应[28]。在葡萄中,转录因子WRKY40和WRKY75与启动子结合参与葡萄抵抗霜霉病入侵的抗病进程[29]。【本研究切入点】本团队前期通过人工接种葡萄白粉菌后的诱导表达分析发现,中国野生毛葡萄()中能够响应白粉菌的诱导上调表达,本研究以不同中国野生葡萄为供试材料,在人工接种白粉菌诱导条件下研究转录因子VqWRKY6和VqbZIP1互作调控葡萄抗白粉病的机理。【拟解决的关键问题】研究中国野生葡萄中抗病相关转录因子基因调控葡萄抗病性的作用,为利用中国野生葡萄资源进行葡萄抗病育种提供理论依据。
1 材料与方法
1.1 试材及取样
供试葡萄材料:中国野生毛葡萄‘商-24’、中国野生华东葡萄()‘白河-35-1’、欧洲葡萄()‘赤霞珠’和‘无核白’,所需葡萄材料均取自西北农林科技大学园艺学院北校区葡萄种质资源圃。在2021年5—6月晴朗天气对健康且长势一致的葡萄叶片进行白粉菌接种处理作为供试材料。
供试模式植物为拟南芥(,Col-0),拟南芥植株在植物光照培养箱中培养,环境温度设置为22℃,光周期L﹕D=16 h﹕8 h。
供试白粉菌孢子采集自西北农林科技大学温室中感病葡萄叶片,植物过表达载体pCAMBIA2300由本实验室马福利博士[30]改造并保存,农杆菌菌株GV3101由本实验室保存。
1.2 基因克隆与载体构建
在液氮速冻的条件下,使用组织研磨仪将‘商-24’和‘赤霞珠’叶片充分研磨,用购自Omega公司的RNA提取试剂盒进行RNA提取,使用购自天根公司的商品化试剂盒进行反转录得到cDNA,根据葡萄基因组网站Grape Genome Browser(https://www.genoscope. cns.fr/externe/GenomeBrowser/Vitis)提供的‘黑比诺’同源基因和序列设计引物VqWRKY6-F/VqWRKY6-R、VqbZIP1-F/VqbZIP1-R(表1),从‘商-24’中克隆和编码序列(coding sequence,CDS),用TaKaRa PrimeSTAR Max DNA Polymerase R045A高保真酶进行PCR扩增,对条带大小正确的扩增产物进行纯化回收,使用无缝克隆技术连接至目标载体。经检测正确的阳性克隆菌液送至北京奥科生物有限公司测序,用DANMAN分析软件进行序列比对,生物信息学分析软件MEGA-X和FigTree v1.4.4用于WRKY转录因子家族成员聚类分析。试验所用引物见表1。

表1 本研究所用引物
1.3 酵母双杂交试验
酵母双杂交验证方法参照文献[31]。将BD- VqWRKY6/AD-VqbZIP1及对照组BD-VqWRKY6/AD分别共转化酵母菌株Y2H Gold,在SD/-Ade/-His/-Leu/ -Trp+AbA+X--Gal四缺培养基平板验证,观察结果并拍照。
1.4 亚细胞定位与双分子荧光互补验证
亚细胞定位:取长势健壮的拟南芥叶片用于制备拟南芥原生质体与转化[32]。亚细胞定位试验中将pCAMBIA2300-VqWRKY6融合质粒与35S-AtHY5- mCherry共转化拟南芥原生质体后进行观察,pCAMBIA2300载体作为对照。利用激光扫描共聚焦显微镜检测GFP和mCherry信号,叶绿体信号显示蓝色和mCherry信号区域区分。
双分子荧光互补:拟南芥原生质体制备和显微镜观察方法同上。将pSPYNE-VqWRKY6质粒与pSPYCE-VqbZIP1质粒共转化拟南芥原生质体后进行观察,以pSPYNE-VqWRKY6和pSPYCE作为对照。叶绿体信号显示红色,利用激光扫描共聚焦显微镜检测YFP信号。
1.5 葡萄瞬时转化和抗病性分析
1.5.1‘赤霞珠’叶片瞬时转化 将融合过表达载体35S-VqWRKY6-GFP与35S-VqbZIP1-GFP转入农杆菌菌株GV3101,将携带重组载体的农杆菌菌液收集菌体重悬至OD600在0.3—0.5左右,使用真空泵通过真空渗透的方式使农杆菌菌液导入葡萄叶片,置于温度为22℃的光照培养箱中进行培养。
1.5.2 人工接种白粉菌 参考王跃进等[33]的接菌方法,采取感白粉病的葡萄叶片,配制孢子悬浮液,用血球计数板调节孢子浓度为2×105个/ml时进行接菌,使用喷壶将配好的孢子悬浮液向瞬时转化24 h后生长状况良好的‘赤霞珠’叶片斜上方喷雾,使叶片表面均匀覆盖细小雾滴。接种后的葡萄叶片在光照培养箱中培养。
1.5.3 葡萄叶片上白粉菌菌丝发育进程观察 采用台盼蓝染色[34]观察接种白粉菌后1—5 d‘赤霞珠’叶片上菌丝的生长发育状况。将叶片放置于罐头瓶中,加入台盼蓝染色液直至叶片完全浸没,沸水浴加热15 min后,去除染色液,用无菌水清洗两次洗净废液后,加入水合氯醛浸没叶片,常温放置过夜脱色。脱色后,用于后续显微镜观察。
1.5.4 活性氧含量检测 对接种白粉菌第5天的葡萄叶片进行3,3′-二氨基联苯胺(DAB)染色,观察过氧化氢(H2O2)的积累,染色后去除DAB染液,加入无水乙醇于沸水浴中孵育5—10 min进行脱色,脱色后的叶片用于后续显微观察。
1.5.5 实时荧光定量PCR(qRT-PCR) 参照OMEGA 公司 RNA 提取试剂盒说明书对接种白粉菌后0—5 d的葡萄叶片提取RNA,对提取的RNA使用天根试剂盒反转录为cDNA,作为qRT-PCR检测模板,以葡萄为内参基因,试验结果取3次生物学重复的平均值,分析抗病基因的表达水平变化。qRT-PCR分析使用TaKaRa商品化定量试剂盒,样品总体积20 μL,反应混合液包括7.4 μL无菌水,0.8 μL引物(上游及下游引物),1 μL模板cDNA及10 μL SYBR Taq酶。
2 结果
2.1 白粉菌诱导下不同葡萄材料WRKY6的表达
接种白粉菌后,中国野生毛葡萄‘商-24’6响应白粉菌诱导表达变化最为明显,且在12 h达到峰值,表达量是接菌前的17.7倍,远高于其他3个材料,推测‘商-24’中在抗病途径中发挥转录调控作用。在‘白河-35-1’中,接菌24 h开始响应白粉菌诱导,后续响应不明显。‘赤霞珠’中在12—72 h有微弱响应。在‘无核白’叶片中接种后12—120 h持续响应白粉菌诱导,72 h表达量达到峰值,为对照组的6.0倍(图1)。
2.2 中国野生毛葡萄‘商-24’中VqWRKY6的克隆与序列分析
利用同源克隆技术从‘商-24’中分离获得编码区序列长度为1 029 bp,编码342个氨基酸(图2-a)。在葡萄基因组网站比对分析发现,定位于葡萄第2号染色体,具体位置为1 228 115—1 229 951位点(图2-b)。
与欧洲葡萄中对应的VvWRKY6进行氨基酸序列对比,发现有3处氨基酸位点发生突变(图2-c),分别为蛋白序列第55处的异亮氨酸突变为蛋氨酸,第213处的组氨酸突变为天冬酰胺,第300处的组氨酸突变为酪氨酸。
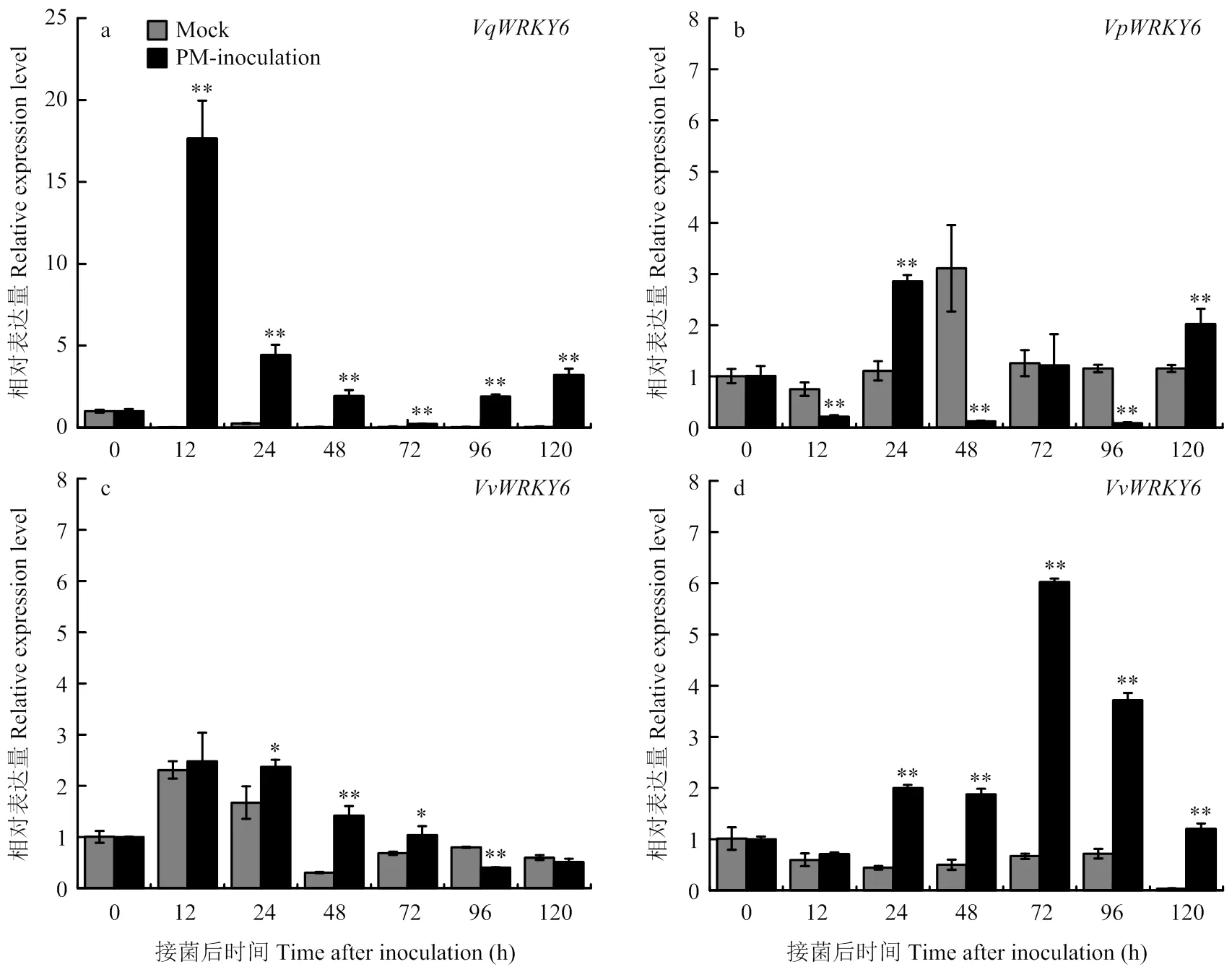
a:商-24 Shang-24;b:白河-35-1 Baihe-35-1;c:赤霞珠Cabernet Sauvignon;d:无核白Thompson Seedless。Mock:无菌水处理葡萄叶片grape leavestreated with sterile water;PM-inoculation:白粉菌接种后葡萄叶片grape leaves inoculated with U. necator
将克隆得到的氨基酸序列和拟南芥及欧洲葡萄中WRKY基因家族成员进行聚类分析,结果表明中国野生毛葡萄中的VqWRKY6属于WRKY家族的group Ⅲ亚家族(图2-d)。
2.3 中国野生毛葡萄VqWRKY6转录因子转录自激活及亚细胞定位分析
将的编码序列构建到酵母表达载体pGBKT7(BD),获得pGBKT7-VqWRKY6融合载体。将融合载体转化至酵母菌株Y2H在SD/-Trp+AbA+ X--Gal培养基上验证。结果发现,在SD/-Trp+AbA+ X--Gal培养基上菌斑全部变蓝(图3-a)。因此,VqWRKY6具有自激活活性。
对VqWRKY6保留保守结构域从N端短截124个氨基酸(图3-b),构建短截后的VqWRKY6-BD载体,进行转录自激活分析,发现自激活活性缺失,可用于后续蛋白互作验证。
将克隆得到的VqWRKY6的编码序列去除终止密码子,构建到pCAMBIA2300-35S-GFP植物表达载体,得到融合载体pCAMBIA2300-VqWRKY6。利用实验室前期构建的35S-AtHY5-mCherry融合载体,与拟南芥中核定位蛋白AtHY5共定位分析。结果表明,VqWRKY6在细胞核发挥转录调控功能(图3-c)。
2.4 酵母双杂交筛选验证中国野生毛葡萄转录因子互作
为了探究转录因子间是否可以相互作用,将构建的pGBKT7-VqWRKY6、pGBKT7-VqMYB153、pGBKT7-VqERF110分别与pGADT7-VqbZIP1、pGADT7- VqWRKY6、pGADT7-VqWRKY53、pGADT7-VqMYB14进行互作验证。将VqMYB153-BD/VqWRKY6-AD、VqMYB153-BD/VqWRKY53-AD、VqMYB153-BD/ VqbZIP1-AD、VqERF110-BD/VqWRKY6-AD、VqERF110- BD/VqMYB14-AD、VqWRKY6-BD/VqMYB14-AD、VqWRKY6-BD/VqMYB15-AD、VqWRKY6-BD/VqbZIP1- AD组合涂在酵母板子SD/-Leu/-Trp上培养,之后取共转化基因的单克隆,以及阴性和阳性对照在无菌蒸馏水中稀释后取2.5 μl滴于SD/-Ade/-His/-Leu/-Trp+ AbA+X--gal培养基上生长,观察发现VqWRKY6- BD/VqbZIP1-AD共转菌斑生长并且显蓝(图4)。
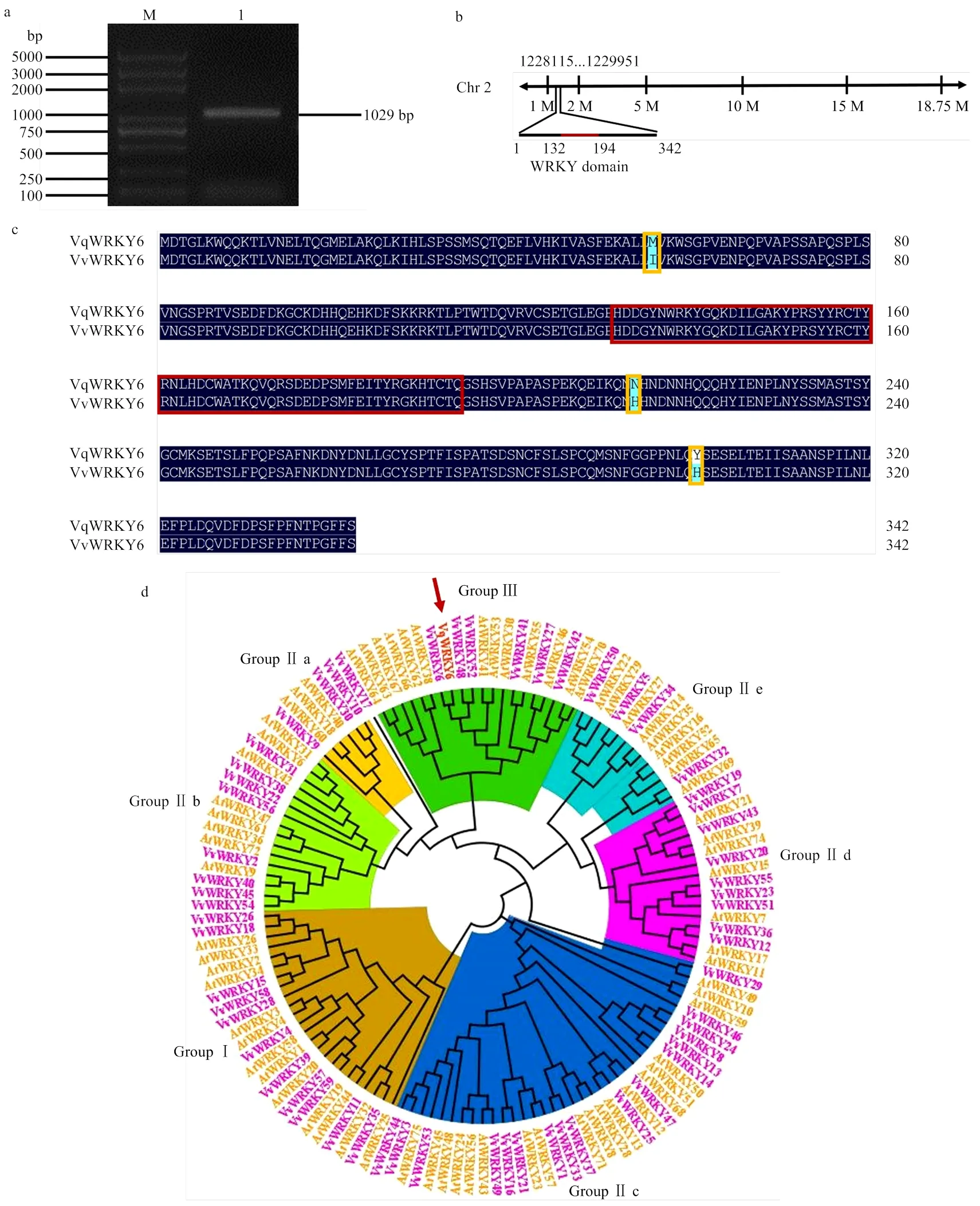
a:M:DNA Marker;1:VqWRKY6克隆VqWRKY6 clone (1029 bp)。b:VqWRKY6定位于葡萄2号染色体,红色标注区域为WRKY结构域VqWRKY6 is located on the grape chromosome 2, and the red labeled area is the WRKY domain。c:中国野生毛葡萄VqWRKY6和欧洲葡萄VvWRKY6氨基酸序列对比,深蓝色部分表示序列一致,黄色方框标注处表示氨基酸序列突变位置,红色方框标注处为WRKY结构域Comparison of the amino acid sequences of Chinese wild V. quinquangularis VqWRKY6 and V. vinifera VvWRKY6, the dark blue part indicates the same sequence, the yellow box indicates the mutation position of the amino acid sequence, and the red box indicates the WRKY domain。d:中国野生毛葡萄转录因子VqWRKY6与欧洲葡萄和拟南芥WRKY家族成员聚类分析。VqWRKY6用红色箭头和红色字体标注,欧洲葡萄WRKY成员用紫色字体标注,拟南芥WRKY成员用黄色字体标注Cluster analysis of VqWRKY6 with WRKY family members from V. vinifera and A. thaliana. VqWRKY6 is marked with red arrow and red font, V. vinifera WRKY members are marked in purple font, and the WRKY members in A. thaliana are marked in yellow font
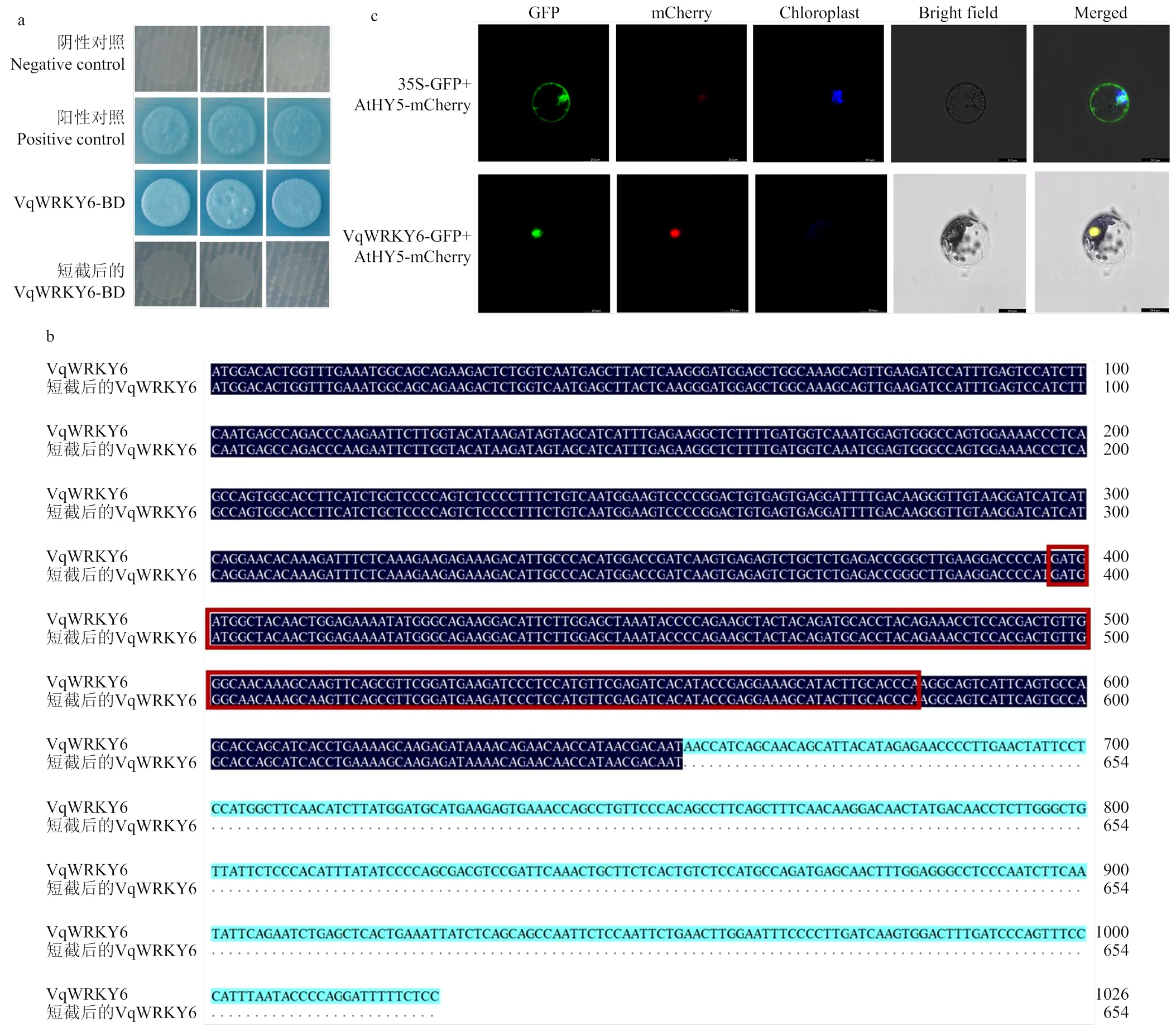
a:VqWRKY6在酵母中转录激活功能分析,序列短截后的VqWRKY6转录自激活分析function analysis of transcriptional activation on VqWRKY6 in yeast, and transcription activation analysis on VqWRKY6 after sequence short-cutting;b:短截后的VqWRKY6与全长VqWRKY6的序列比对分析alignment of VqWRKY6 sequence in short-cutting and full-length;c:VqWRKY6在拟南芥原生质体中的亚细胞定位subcellular localization of VqWRKY6 in A. thaliana protoplasts。GFP:绿色荧光信号green fluorescence signal;mCherry:红色荧光信号red fluorescence signal;chloroplast:叶绿体自发荧光chloroplast autofluorescence;Bright field:明场图bright field plot;Merged:组合图combined graph
2.5 中国野生毛葡萄VqWRKY6与VqbZIP1互作
以欧亚种葡萄‘黑比诺’品系‘PN40024’基因组信息为参考。克隆出长度为900 bp的,编码299个氨基酸(图5-a)。将转录因子VqbZIP1连接AD载体得到pGADT7-VqbZIP1载体,与pGBKT7- VqWRKY6共转化酵母细胞进行酵母双杂交验证。发现融合载体pGADT7-VqWRKY6与pGBKT7-VqbZIP1共转化的酵母菌株在四缺培养基SD/-Ade/-His/-Leu/ -Trp+X--Gal+AbA上能够正常生长,形成菌斑且变蓝(图5-b)。说明在酵母系统中,VqWRKY6与VqbZIP1能够发生相互作用形成转录调控复合体,推测这种互作现象可能存在于植物体中。为了进一步在植物细胞中验证这种相互作用,将VqWRKY6与VqbZIP1的CDS全长序列去除终止密码子,并构建到pSPYNE和pSPYCE上得到融合载体,转入拟南芥原生质体进行双分子荧光互补验证。结果表明,在拟南芥原生质体的细胞核内检测到YFP的黄色荧光信号(图5-c)。

图4 酵母双杂交筛选中国野生毛葡萄转录因子间互作
2.6 转录因子VqWRKY6和VqbZIP1瞬时表达与增强葡萄对白粉菌的抗性
2.6.1 白粉菌接种观察及抗病性分析 在单独过表达的叶片,以及共同过表达和的叶片,相对于单独过表达以及对照叶片白粉病症不明显,而在单独过表达以及对照叶片上白粉菌菌丝体的生长则较为明显,可以观察到肉眼可见的白粉菌菌落(图6-a)。根据观察结果推测,单独过表达、共同过表达与可以增强葡萄对白粉病的抗性。
为了进一步探究VqWRKY6提高葡萄对白粉病抗性的机理,对瞬时转化后的‘赤霞珠’叶片接种白粉菌,利用台盼蓝进行葡萄组织化学染色,观察0—5 d内白粉菌孢子萌发和菌丝发育进程。显微观察结果表明,共同过表达和的叶片组织菌丝生长发育进程较单独过表达的叶片迟缓,而单独过表达的叶片上菌丝生长发育状况与对照组相比差异较小。在叶片接种白粉菌12 h后,单独过表达与共同过表达和的叶片表面并未观察到萌发的孢子,表明白粉菌孢子已附着于叶片表面;对照组和单独过表达的叶片表面白粉菌孢子开始萌发伸出菌丝。在24—120 h观察到单独过表达和共同过表达和的叶片上白粉菌孢子陆续萌发,但菌丝生长速率较对照组和单独过表达的叶片显著迟缓,且次生菌丝体较为稀疏(图6-b)。
上述结果表明,在葡萄中过表达能够有效减缓白粉菌孢子的萌发,减慢菌丝的生长,在葡萄中表达和可减缓白粉菌的菌丝发育进程,而VqWRKY6和VqbZIP1的协同互作能够进一步增强葡萄对白粉病的抗性。
在病原菌入侵过程中,活性氧作为抗病相关信号传导途径的重要组分发挥作用[35]。为了探究转录因子VqWRKY6及其互作蛋白VqbZIP1是否参与调控病原诱导下的活性氧产生和积累,对瞬时转化后的‘赤霞珠’叶片进行白粉菌接种,取接种后5 d的叶片进行DAB染色,结果发现,在过表达叶片组织接种区域均有不同程度的红褐色加深状况,单独过表达的叶片红褐色积累程度高于单独过表达的叶片,说明单独过表达的叶片活性氧含量高于单独过表达的叶片。其中在共同过表达和的叶片上红褐色程度最深,说明共同过表达和的叶片组织的活性氧含量高于单独过表达和单独过表达叶片(图6-c)。
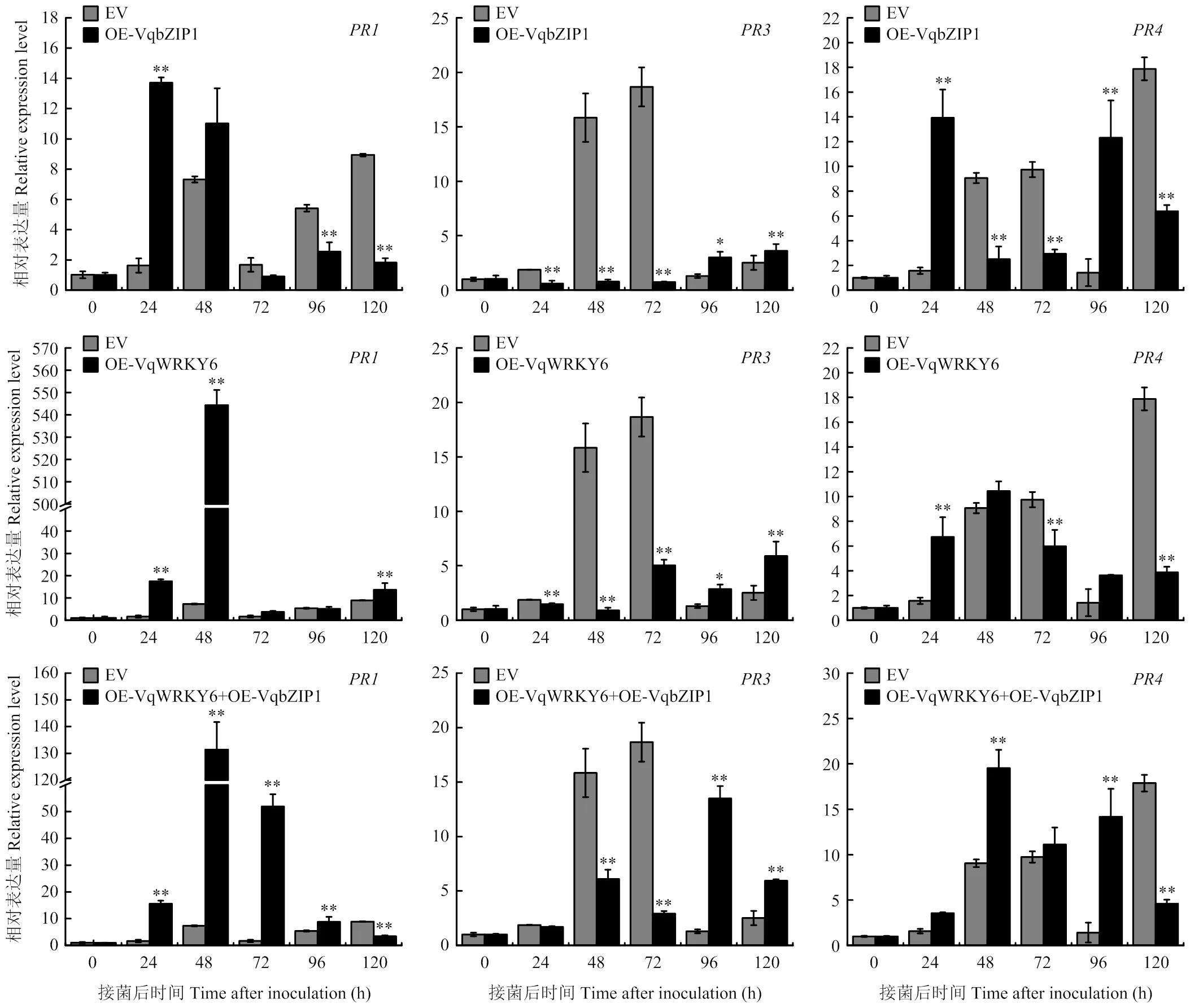
2.6.2 过表达葡萄叶片在白粉菌诱导下抗病基因的表达情况 为了进一步探究VqWRKY6参与调控葡萄抗病性的分子机理,以及VqWRKY6与葡萄抗病基因在调控抗病进程中的关系,通过qRT-PCR分析瞬时转化后的‘赤霞珠’叶片在接种白粉菌前后病程相关蛋白基因表达量的动态变化。结果表明,与对照组相比,在共同过表达和的葡萄叶片中,接种白粉菌72 h后的表达水平显著上调,是同时期对照组的30.9倍,而单独过表达和单独过表达的葡萄叶片中仅在24—48 h上调表达,而在接种后72 h表达水平恢复至常态;接种白粉菌96 h后表达量明显上调,其中共同过表达和叶片中的表达量在96 h达到峰值,是同时期对照组的10.4倍,而单独过表达和单独过表达的葡萄叶片表达水平在96 h和120 h有微弱响应(图7)。
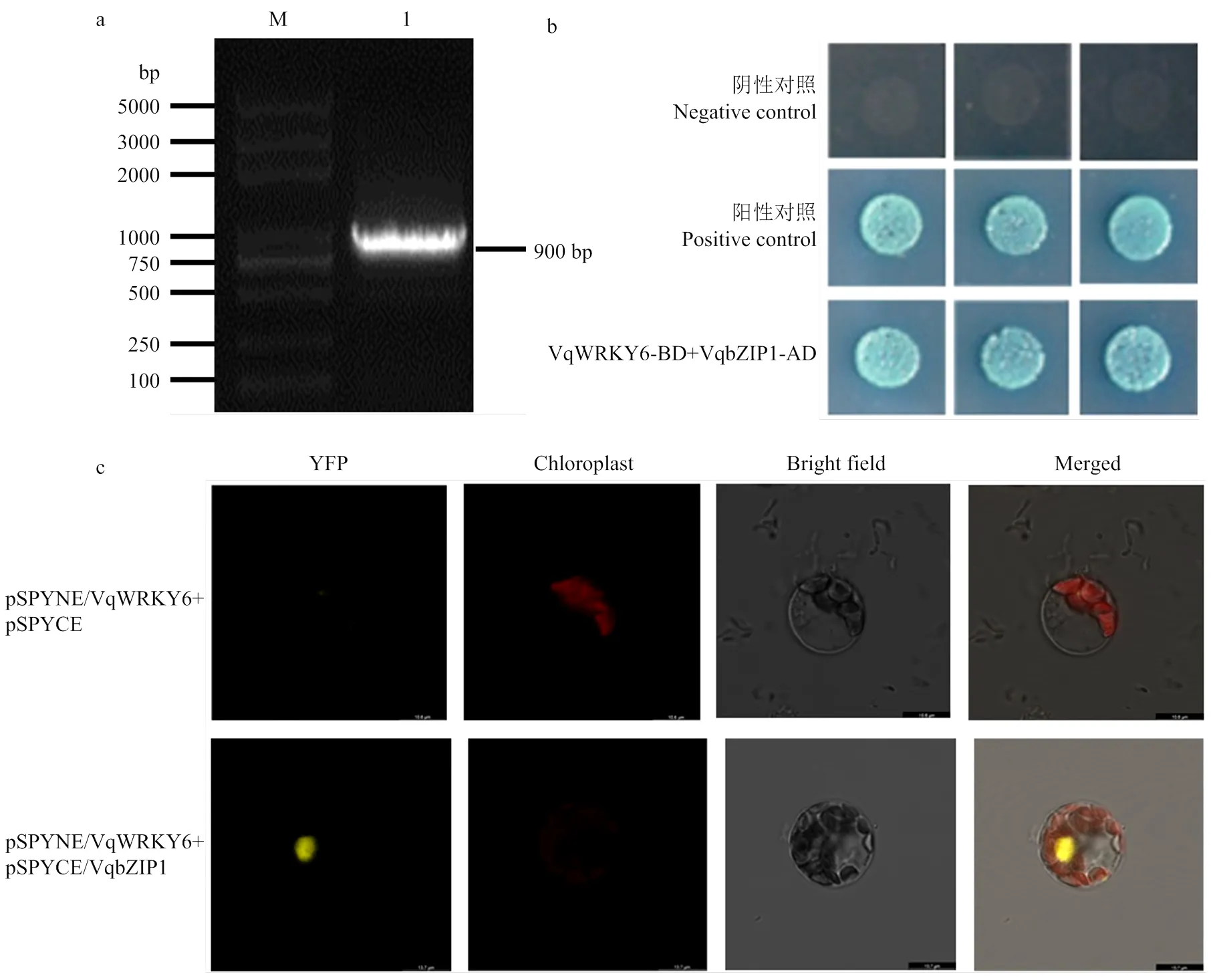
a:M:DNA Marker;1:VqbZIP1克隆VqbZIP1 clone (900 bp)。b:酵母双杂交验证VqWRKY6与VqbZIP1的互作The yeast two-hybrid was carried out to verify the interaction between VqWRKY6 and VqbZIP1。c:双分子荧光互补技术验证VqWRKY6与VqbZIP1的互作。pSPYNE-VqWRKY6和pSPYCE-VqbZIP1共转入拟南芥原生质体,以pSPYNE-VqWRKY6和pSPYCE共转入拟南芥原生质体作为对照To confirm the interaction between VqbZIP1 and VqWRKY6, the assay of bimolecular fluorescence complementation was performed. pSPYNE-VqWRKY6 and pSPYCE-VqbZIP1 were co-transferred toA. thalianaprotoplasts, and pSPYNE-VqWRKY6 and pSPYCE were co-transferred to A. thaliana protoplasts as control
3 讨论
3.1 WRKY转录因子增强抗病性
在高等植物中WRKY转录因子是最大的转录因子家族之一,是植物先天免疫系统和后天系统获得性抗性中的核心组件[10,36]。在众多共调控拟南芥防御基因的启动子中存在WRKY转录因子基因的结合位点(C/TTGACC/T,W box),这为证明锌指型WRKY转录因子在调节防御中有广泛而关键的作用提供了间接证据[37]。在过表达苹果‘GL-3’株系中调控抗病性显著增强[38];在苹果愈伤组织过表达,提高了苹果愈伤组织对轮纹病的抗性[39];拟南芥中异源表达,也显著提高其对轮纹病的抗性[40]。PANDEY等[41]研究发现白粉病入侵时,拟南芥AtWRKY18和AtWRKY40的突变体中与植物抗毒素合成相关的关键基因和的表达量有显著升高趋势;LI等[42]研究表明,中国野生华东葡萄‘白河-35-1’中和响应SA途径和白粉菌诱导表达,在拟南芥中异源表达和转基因株系对白粉病的抗性显著提高;乔恒波[43]研究表明,在拟南芥中异源表达基因可提高对白粉病及DC3000的抗性。
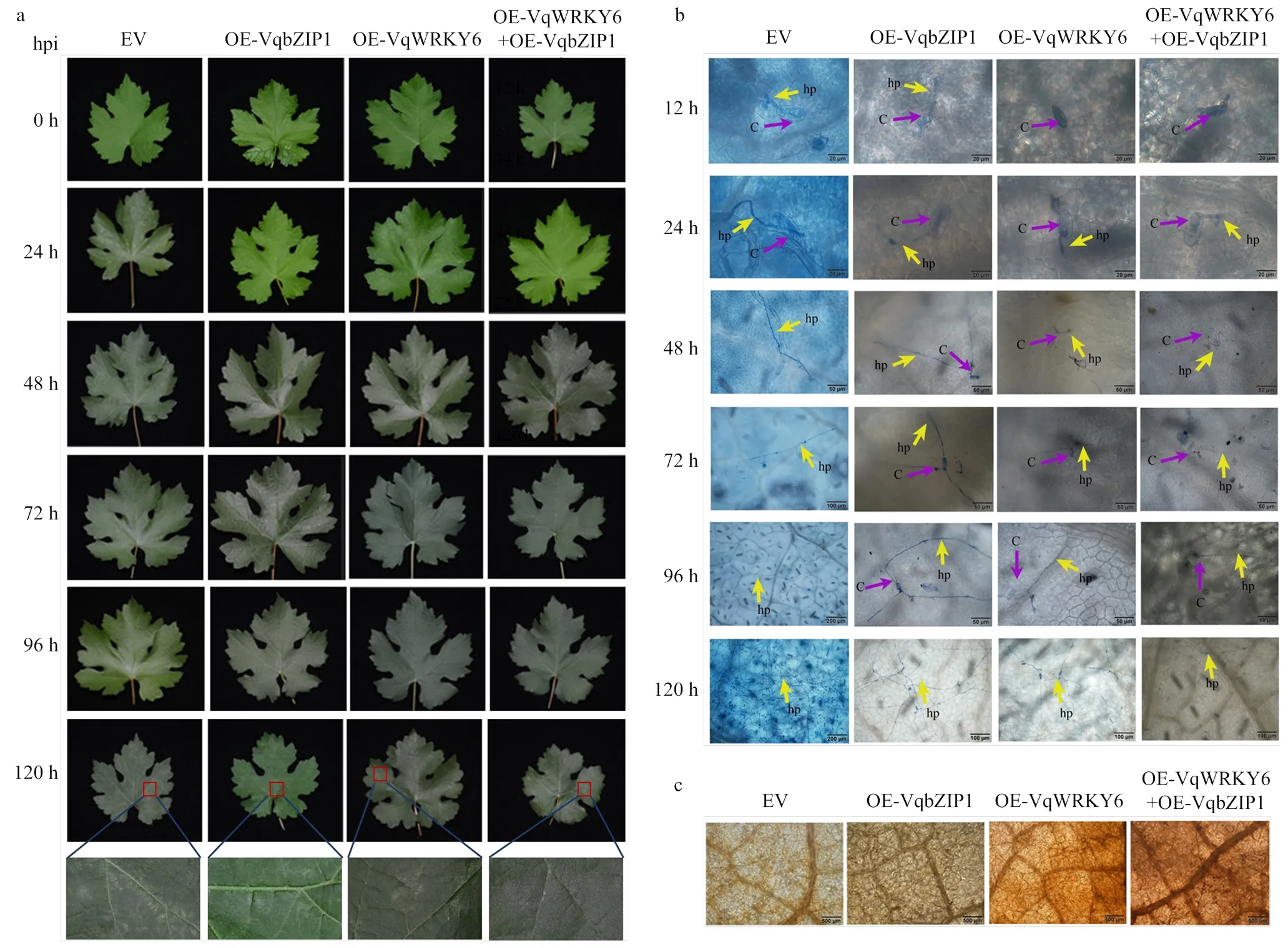
a:取瞬时转化后24 h的葡萄叶片进行人工接种白粉菌,对接种后0—120 h的叶片进行观察,红色方框标示白粉菌菌丝体Grape leaves were inoculated with U. necator at 24 h after transient transformation, the observation on leaves was performed at 0-120 h after inoculation; Red box indicates the mycelium of U. necator;EV:空载体对照empty vector control;OE-VqbZIP1:单独过表达VqbZIP1overexpressing of VqbZIP1 alone;OE-VqWRKY6:单独过表达VqWRKY6 overexpressing of VqWRKY6alone;OE-VqWRKY6+OE-VqbZIP1:共同过表达VqWRKY6和VqbZIP1 Co-overexpressing of VqWRKY6 and VqbZIP1;hpi:白粉菌接种后时间Hours post inoculation。b:对瞬时转化后24 h的葡萄叶片进行人工接种白粉菌,显微观察菌丝发育生长进程Grape leaves were inoculated with U. necator at 24 h after transient transformation, and hyphae development and growth process were microscopically observed;c:分生孢子conidium;hp:初级菌丝primary hypha。c:DAB染色检测葡萄叶片组织中H2O2含量DAB staining is used to detect H2O2 content in grape tissues
葡萄基因组共有59个WRKY转录因子成员,依据序列特征和系谱进化分Ⅰ、Ⅱ、Ⅲ 3个组。Ⅲ组基因包含C2HC锌指结构(C-X7-X23-H-X1-C),有6个属于此组,分别为、、、、、[16]。将同源克隆获得的中国野生毛葡萄VqWRKY6蛋白序列与拟南芥和欧洲葡萄的WRKY家族成员进行聚类分析,发现VqWRKY6属于group Ⅲ族。group Ⅲ族的WRKY转录因子在植物抗逆和抗病上起重要作用。在棉花中WRKY group Ⅲ族基因广泛参与应对非生物胁迫[44];在潘那利番茄中,WRKY41会迅速响应应对不利环境[45];在拟南芥植株中异源表达WRKY group Ⅲ成员提高了对白粉病的抗性[46]。
EV:空载体对照empty vector control;OE-VqbZIP1:单独过表达VqbZIP1overexpressing of VqbZIP1 alone;OE-VqWRKY6:单独过表达VqWRKY6 overexpressing of VqWRKY6alone;OE-VqWRKY6+OE-VqbZIP1:共同过表达VqWRKY6和VqbZIP1 Co-overexpressing of VqWRKY6 and VqbZIP1
中国野生毛葡萄‘商-24’对白粉病具有显著抗性[6]。对抗病和感病葡萄进行人工接种白粉菌,观察的表达模式发现,‘商-24’中迅速响应白粉菌诱导表达,在12 h达到峰值,且后续也较对照组持续上调表达。在欧洲葡萄中,响应白粉菌诱导表达弱于‘商-24’。且通过观察接种白粉菌后葡萄叶片发病表征,发现过表达葡萄叶片白粉菌菌丝生长状况不明显。推测VqWRKY6与葡萄白粉病抗性调控相关。
3.2 VqWRKY6与VqbZIP1互作促进活性氧积累
活性氧作为信号分子参与应对植物抵抗病原物入侵,对病原体攻击部位防御最快的反应是氧化爆发,主要是超氧化物和过氧化氢[47]。积累的过氧化氢可能通过参与细胞壁强化,诱导防御基因表达等增强抗病性[35,48]。DAB染色结果发现在接种白粉菌位置,葡萄叶片显色加深,共同过表达和的叶片红褐色程度最深,表明其活性氧积累水平最高,推测和互作可以刺激葡萄组织活性氧的产生从而增强对白粉病抗性。
3.3 VqWRKY6与VqbZIP1互作促进抗病基因表达
WRKY转录因子可参与多种植物对不同胁迫响应的激素信号传导途径[49]。拟南芥中转录因子WRKY70作为WRKY group Ⅲ亚族成员表达受SA诱导,过表达的拟南芥对白粉病抗性增强[50-51]。VqWRKY52通过作用于SA介导信号传导途径增强抗病性[46]。葡萄中过表达可以上调JA途径抗病相关基因表达,提高转基因株系的抗病性[52]。受SA特异性诱导,JA途径诱导、[53-54]。定量结果发现,WRKY6不仅能够增强SA途径的表达,也能增强JA途径和的表达,说明WRKY6能够通过参与SA和JA两种抗病途径,调控增强葡萄的抗病性。本研究中共同过表达和葡萄叶片中和的表达量显著上升,推测VqWRKY6与VqbZIP1互作可能促进JA途径表达,从而提高对白粉病抗性。
4 结论
中国野生毛葡萄‘商-24’中转录因子VqWRKY6与VqbZIP1互作调控叶片组织活性氧产生和积累,使菌丝发育进程变缓,抗病基因、显著上调表达。‘商-24’VqWRKY6与VqbZIP1互作调控增强了抗病性,‘商-24’可作为葡萄抗病育种的种质资源与材料。
[1] 贺普超, 王跃进, 王国英, 任治邦, 和纯成. 中国葡萄属野生种抗病性的研究. 中国农业科学, 1991, 24(3): 50-56.
HE P C, WANG Y J, WANG G Y, REN Z B, HE C C. The studies on disease-resistance ofwild species originated in China. Scientia Agricultura Sinica, 1991, 24(3): 50-56. (in Chinese)
[2] ARMIJO G, SCHLECHTER R, AGURTO M, MUNOZ D, NUNEZ C, ARCE-JOHNSON P. Grapevine pathogenic microorganisms: understanding infection strategies and host response scenarios. Frontiers in Plant Science, 2016, 7: 382.
[3] DÉLYE C, LAIGRET F, CORIO-COSTET M F. A mutation in the 14-demethylase gene ofthat correlates with resistance to a sterol biosynthesis inhibitor. Applied and Environmental Microbiology, 1997, 63(8): 2966-2970.
[4] DONALD T M, PELLERONE F, ADAM-BLONDON A F, BOUQUET A, THOMAS M R, DRY I B. Identification of resistance gene analogs linked to a powdery mildew resistance locus in grapevine. Theoretical and Applied Genetics, 2002, 104(4): 610-618.
[5] TAKSONYI P, KOCSIS L, MÁTYAS K K, TALLER J. The effect of quinone outside inhibitor fungicides on powdery mildew in a grape vineyard in hungary. Scientia Horticulturae, 2013, 161: 233-238.
[6] WANG Y, LIU Y, P H. E, CHEN J, LAMIKANRA O, LU J. Evaluation of foliar resistance toin Chinese wildspecies. Vitis, 1995, 34(3): 159-164.
[7] XU W, MA F, LI R, ZHOU Q, YAO W, JIAO Y, ZHANG C, ZHANG J, WANG X, XU Y, WANG Y. VpSTS29/STS2 enhances fungal tolerance in grapevine through a positive feedback loop. Plant, Cell and Environment, 2019, 42(11): 2979-2998.
[8] EULGEM T. Regulation of thedefense transcriptome. Trends in Plant Science, 2005, 10(2): 71-78.
[9] RYU H S, HAN M, LEE S K, CHO J I, RYOO N, HEU S, LEE Y H, BHOO S H, WANG G L, HAHN T R, JEON J S. A comprehensive expression analysis of the WRKY gene superfamily in rice plants during defense response. Plant Cell Reports, 2006, 25(8): 836-847.
[10] EULGEM T, SOMSSICH I E. Networks of WRKY transcription factors in defense signaling. Current Opinion in Plant Biology, 2007, 10(4): 366-371.
[11] NAOUMKINA M A, HE X Z, DIXON R A. Elicitor-induced transcription factors for metabolic reprogramming of secondary metabolism in. BMC Plant Biology, 2008, 8: 132.
[12] RIECHMANN J L, RATCLIFFE O J. A genomic perspective on plant transcription factors. Current Opinion in Plant Biology, 2000, 3(5): 423-434.
[13] 楚宗丽, 张睿男, 李亮杰, 孙君艳, 王付娟, 周强, 仝胜利. 小麦WRKY转录因子的鉴定及其在胚性愈伤组织形成中的表达分析. 麦类作物学报, 2021, 41(12): 1469-1478.
CHU Z L, ZHANG R N, LI L J, SUN J Y, WANG F J, ZHOU Q, TONG S L. Identification of wheat WRKY transcription factor and its expression analysis in embryonic callus formation. Journal of Triticeae Crops, 2021, 41(12): 1469-1478. (in Chinese)
[14] SRIVASTAVA R, KUMAR S, KOBAYASHI Y, KUSUNOKI K, Tripathi P, Kobayashi Y, Koyama H,SAHOO L. Comparative genome-wide analysis of WRKY transcription factors in two Asian legume crops: Adzuki bean and mung bean. Scientific Reports, 2018, 8(1): 16971.
[15] GOFF S A, RICKE D O, LAN T H, PRESTING G G, WANG R, DUNN M, GLAZEBROOK J, SESSIONS A, OELLER P, VARMA H,. A draft sequence of the rice genome (L. ssp.). Science, 2002, 296(5565): 92-100.
[16] GUO C L, GUO R R, XU X Z, GAO M, LI X Q, SONG J Y, ZHENG Y, WANG X P. Evolution and expression analysis of the grape (L.) WRKY gene family. Journal of Experimental Botany, 2014, 65(6): 1513-1528.
[17] CHUJO T, MIYAMOTO K, OGAWA S, MASUDA Y, SHIMIZU T, KISHI-KABOSHI M, TAKAHASHI A, NISHIZAWA Y, MINAMI E, NOJIRI H, YAMANE H, OKADA K. Overexpression of phosphomimic mutatedleads to enhanced blast resistance in rice. Plos One, 2014, 9(6): e98737.
[18] WANG D, JIANG C Y, LIU W D, WANG Y J. The WRKY53 transcription factor enhances stilbene synthesis and disease resistance by interacting with MYB14 and MYB15 in Chinese wild grape. Journal of experimental botany, 2020, 71(10): 3211-3226.
[19] 吴凤颖, 刘梦琦, 王跃进. 中国野生毛葡萄芪合酶基因抗白粉病功能分析. 园艺学报, 2020, 47(2): 205-219.
WU F Y, LIU M Q, WANG Y J. Functional analysis of thestilbene synthase genesandof the resistance to powdery mildew in. Acta Horticulture Sinica, 2020, 47(2): 205-219. (in Chinese)
[20] HU Y R, DONG Q Y, YU D Q.WRKY46 coordinates with WRKY70 and WRKY53 in basal resistance against pathogen. Plant Science, 2012, 185/186: 288-297.
[21] BIRKENBIHL R P, DIEZEL C, SOMSSICH I E.WRKY33 is a key transcriptional regulator of hormonal and metabolic responses towardinfection. Plant Physiology, 2012, 159(1): 266-285.
[22] MENG Y, WISE R P. HvWRKY10, HvWRKY19, and HvWRKY28 regulate Mla-triggered immunity and basal defense to barely powdery mildew. Molecular Plant-microbe Interactions, 2012, 25(11): 1492-1505.
[23] BI M M, LI X Y, YAN X, LIU D, GAO G, ZHU P F, MAO H Y. Chrysanthemum WRKY15-1 promotes resistance toHenn. via the salicylic acid signaling pathway. Horticulture Research, 2021, 8: 6.
[24] YIN W C, WANG X H, LIU H, WANG Y, NOCKER S, TU M X, FANG J H, GUO J Q, LI Z, WANG X P. Overexpression ofenhances powdery mildew resistance in grapevine by promoting salicylic acid signaling and specific metabolite synthesis. Horticulture Research, 2022, 9: uhab064.
[25] CHRISTENSEN A B, CHO B H, NæSBY M, GREGERSEN P L, BRANDT J, MADRIZ-ORDEÑANA K, COLLINGE D B, THORDAL- CHRISTENSEN H. The molecular characterization of two barley proteins establishes the novel PR-17 family of pathogenesis-related proteins. Molecular plant pathology, 2002, 3(3): 135-144.
[26] VAN LOON L C, REP M, PIETERSE C M J. Significance of inducible defense-related proteins in infected plants. Annual Review of Phytopathology, 2006, 44: 135-162.
[27] HE M Y, XU Y, CAO J L, ZHU Z G, JIAO Y T, WANG Y J, GUAN X, YANG Y Z, XU W R, FU Z F. Subcellular localization and functional analyses of a PR10 protein gene fromin response toinfection.Protoplasma, 2013, 250(1): 129-140.
[28] 马辉. 中国野生华东葡萄基因的克隆与功能分析[D]. 杨凌: 西北农林科技大学, 2014.
MA H. Molecular cloning and functional analysis ofgene in Chinese wild[D]. Yangling: Northwest A&F University, 2014. (in Chinese)
[29] 刘兵, 李梦媛, 张娜, 尚博兴, 刘国甜, 徐炎. 中国野生葡萄抗霜霉病相关基因及其启动子的克隆和功能分析. 园艺学报, 2021, 48(2): 265-275.
LIU B, LI M Y, ZHANG N, SHANG B X, LIU G T, XU Y. Cloning and functional analysis of the CDS and promoter ofgene response to downy mildew in Chinese wild grape. Acta Horticulture Sinica, 2021, 48(2): 265-275. (in Chinese)
[30] MA F L, Wang L, Wang Y J. Ectopic expression of, a stilbene synthase gene from, indicates STS presence in cytosolic oil bodies. Planta, 2018, 248(1): 89-103.
[31] 姚文孔. 中国野生华东葡萄泛素连接酶基因功能研究[D]. 杨凌: 西北农林科技大学, 2017.
YAO W K. Function analyses of E3 ubiquitin ligase genefrom Chinese wild grape[D]. Yangling: Northwest A&F University, 2017. (in Chinese)
[32] YOO S D, CHO Y H, SHEEN J.mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nature Protocols, 2007, 2(7): 1565-1572.
[33] 王跃进, 贺普超, 张剑侠. 葡萄抗白粉病鉴定方法的研究. 西北农林科技大学学报, 1999, 27(5): 6-10.
WANG Y J, HE P C, ZHANG J X. Studies on the methods of resistance toin. Journal of Northwest A&F University, 1999, 27(5): 6-10. (in Chinese)
[34] MICALI C, GÖLLNER K, HUMPHRY M, CONSONNI C, PANSTRUGA R. The powdery mildew disease of: A paradigm for the interaction between plants and biotrophic fungi//TheBook. The American Society of Plant Biologists, 2008, 6: e0115.
[35] LEVINE A, TENHAKEN R, DIXON R, LAMB C. H2O2from the oxidative burst orchestrates the plant hypersensitive disease resistance response. Cell, 1994, 79(4): 583-593.
[36] JIANG J J, MA S H, YE N H, JIANG M, CAO J S, ZHANG J H. WRKY transcription factors in plant responses to stresses. Journal of Integrative Plant Biology, 2017, 59(2): 86-101.
[37] SOMSSICH I E. Networks of transcriptional regulation underlying plant defense responses towards phytopathogens//Grasser K D. Regulation of Transcription in Plants. Blackwell Publishing, 2006: 266-284.
[38] 张远嬿. 苹果基因的克隆与功能分析[D]. 沈阳: 沈阳农业大学, 2018.
ZHANG Y Y. Cloning and functional analysis ofgene in apple[D]. Shenyang: Shenyang Agricultural University, 2018. (in Chinese)
[39] 周茜茜. 苹果轮纹病激发的SA特异性诱导表达基因的抗病功能鉴定[D]. 泰安: 山东农业大学, 2019.
ZHOU Q Q. Identification of disease resistance of sa-specific inducible genestimulated by[D]. Taian: Shandong Agricultural University, 2019. (in Chinese)
[40] 周茜茜, 邱化荣, 何晓文, 王宪璞, 刘秀霞, 李保华, 吴树敬, 陈学森.介导提高苹果与拟南芥对轮纹病菌的免疫抗性. 中国农业科学, 2018, 51(21): 4052-4064.
ZHOU Q Q, QIU H R, HE X W, WANG X P, LIU X X, LI B H, WU S J, CHEN X S.mediated improvement of immune resistance of apple andto. Scientia Agricultura Sinica, 2018, 51(21): 4052-4064. (in Chinese)
[41] PANDEY S P, ROCCARO M, SCHON M, LOGEMANN E, SOMSSICH I E. Transcriptional reprogramming regulated by WRKY18 and WRKY40 facilitates powdery mildew infection of. The Plant Journal, 2010, 64(6): 912-923.
[42] LI H, XU Y, XIAO Y, ZHU Z G, XIE X Q, ZHAO H Q, WANG Y J. Expression and functional analysis of two genes encoding transcription factors,and, isolated from Chinese wild. Planta, 2010, 232(6): 1325-1337.
[43] 乔恒波. 中国野生毛葡萄转录因子基因克隆与功能研究[D]. 杨凌: 西北农林科技大学, 2016.
QIAO H B. Cloning and function analysis of atranscription factor in[D]. Yangling: Northwest A&F University, 2016. (in Chinese)
[44] DOU L L, GUO Y N, ONDATI E, PANG C Y, WEI H L, SONG M Z, FAN S L, YU S X. Identification and expression analysis of group Ⅲ WRKY transcription factors in cotton. Journal of Integrative Agriculture, 2016, 15(11): 2469-2480.
[45] 魏娟娟, 杨伟, 潘宇, 张兴国, 李金华. 番茄41基因的克隆、表达分析与转基因植株的获得. 西南大学学报(自然科学版), 2017, 39(1): 46-54.
WEI J J, YANG W, PAN Y, ZHANG X G, LI J H. Cloning and expression analysis of a41 gene in tomato and its transfer into a tomato cultivar. Journal of Southwest University (Natural Science Edition), 2017, 39(1): 46-54. (in Chinese)
[46] WANG X, GUO R, TU M, WANG D, GUO C, WAN R, LI Z, WANG X. Ectopic expression of the wild grape WRKY transcription factor VqWRKY52 inenhances resistance to the biotrophic pathogen powdery mildew but not to the necrotrophic pathogen. Frontiers in Plant Science, 2017, 8: 97.
[47] APEL K, HIRT H. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annual Review of Plant Biology, 2004, 55: 373-399.
[48] MADER M, FUSSL R. Role of peroxidase in lignification of tobacco cells.Ⅱ. Regulation by phenolic compounds. Plant Physiology, 1982, 70(4): 1132-1134.
[49] FINATTO T, VIANA V E, WOYANN L G, BUSANELLO C, MAIA L C, OLIVEIRA A. Can WRKY transcription factors help plants to overcome environmental challenges? Genetics and Molecular Biology, 2018, 41(3): 533-544.
[50] LI J, BRADER G, PALVA E T. The WRKY70 transcription factor: a node of convergence for jasmonate-mediated and salicylate-mediated signals in plant defense. The Plant Cell, 2004, 16(2): 319-331.
[51] LI J, BRADER G, KARIOLA T, PALVA E T. WRKY70 modulates the selection of signaling pathways in plant defense. The Plant Journal,2006, 46(3): 477-491.
[52] MARCHIVE C, MZID R, DELUC L, BARRIEU F, PIRRELLO J, GAUTHIER A, CORIO-COSTET M F, REGAD F, CAILLETEAU B, HAMDI S, Lauvergeat v. Isolation and characterization of atranscription factor, VvWRKY1, and its effect on responses to fungal pathogens in transgenic tobacco plants. Journal of Experimental Botany, 2007, 58(8): 1999-2010.
[53] MØLLER S G, CHUA N H. Interactions and intersections of plant signaling pathways. Journal of Molecular Biology, 1999, 293(2): 219-234.
[54] CHEONG Y H, CHANG H S, GUPTA R, WANG X, ZHU T, LUAN S. Transcriptional profiling reveals novel interactions between wounding, pathogen, abiotic stress, and hormonal responses in. Plant Physiology, 2002, 129(2): 661-677.
Functional analysis of the interaction between transcription factors VqWRKY6 and VqbZIP1 in regulating the resistance to powdery mildew in Chinese wild
Zhang Jie, Jiang ChangYue, WANG YueJin
College of Horticulture, Northwest Agriculture and Forestry University/State Key Laboratory of Crop Stress Biology in Arid Areas/Key Laboratory of Horticultural Plant Germplasm Resource Utilization in Northwest China, Ministry of Agriculture and Rural Affairs, Yangling 712100, Shaanxi
【Objective】As the world’smain cultured varieties,has the advantages of high yield and good quality, however, the resistance of.to disease is poor. Powdery mildew is a fungal disease that seriously endangers viticulture. The resources of Chinese wild grape are abundant, which can provide sufficient germplasm resources for disease-resistant breeding. This objective of this study is to screen grape transcription factor genes that regulate the resistance to powdery mildew, and to explore the mechanism of these genes regulating the resistance to powdery mildew, which can provide high-quality genetic resources for selection and breeding of grape disease-resistant varieties.【Method】was cloned from Chinese wild‘Shang-24’. The sequences were analyzed using DANMAN and MEGA-X software and subcellular localization analysis was adopted usingprotoplasts by PEG-mediated transformation, which was performed for the location of transcriptional regulation. The yeast two-hybrid and bimolecular fluorescence complementation were adopted to prove that VqWRKY6 can interact with the transcription factor VqbZIP1 to form a transcriptional complex. Taking the leaves of the disease-susceptible grape ‘Cabernet Sauvignon’ as the material, the transient transformation mediated bywas performed in the leaves of ‘Cabernet Sauvignon’. Afterinoculation on the leaves, the pathogenesis symptoms were observed, hyphal development progression was microscopically visualized with trypan blue staining, and reactive oxygen species (ROS) accumulation was visualized using DAB staining. Thedifferences between grape leaves co-overexpressing ofand, overexpressing ofalone, overexpressing ofalone, and the control group were compared. The qRT-PCR was used to analyze the expression level of anti-disease genes underinduction.【Result】is located on grape chromosome 2, encodes 342 amino acids and belongs to the group Ⅲ subfamily of the WRKY family. VqWRKY6 exerts a transcriptional regulatory function in nucleus. The propagation rate ofon the surface of ‘Cabernet Sauvignon’ leaves after co-overexpressing ofandwas significantly slower than that of leaves overexpressing ofaloneand overexpressing ofalone, and the ROS content in the leaf tissues co-overexpressing ofandwas significantly higher than that of leaves overexpressingofalone and overexpressingofalone. In addition, the synergistic regulation of VqWRKY6 and VqbZIP1 could activateandof the jasmonate (JA) pathway, and the gene expression levels were significantly upregulated.【Conclusion】The synergistic effect of VqWRKY6 and VqbZIP1 may improve the disease resistance of grape to powdery mildew by activating the JA pathway, promoting the production of ROS, enhancing the expression of disease-resistant genes to inhibit the growth of. VqWRKY6 and VqbZIP1 are important disease-resistant genetic resources, and ‘Shang-24’ is an important disease-resistant germplasm resource.
Chinese wild; WRKY transcription factor; powdery mildew; transcriptional regulation

10.3864/j.issn.0578-1752.2022.23.005
2022-06-28;
2022-08-08
国家自然科学基金面上项目(31872055)
张洁,E-mail:jiee@nwafu.edu.cn。通信作者王跃进,E-mail:wangyj@nwsuaf.edu.cn
(责任编辑 岳梅)
