基于TMT定量蛋白质组学揭示纳米包装双孢蘑菇采后冷藏生理代谢规律
2022-02-02王朝方东路张攀容姜雯裴斐胡秋辉马宁
王朝,方东路,张攀容,姜雯,裴斐,胡秋辉,马宁
基于TMT定量蛋白质组学揭示纳米包装双孢蘑菇采后冷藏生理代谢规律

1南京财经大学食品科学与工程学院/江苏省现代粮食流通与安全协同创新中心/江苏高校粮油质量安全控制及深加工重点实验室,南京 210023;2南京林业大学轻工与食品学院,南京 210037
【背景】双孢蘑菇采后极易发生开伞、失水及褐变等品质劣变现象,极大地影响了其贮藏品质和商业价值。前期研究已证实纳米包装可有效延缓双孢蘑菇采后的品质劣变,但其保鲜机制仍不清晰。【目的】本研究通过串联质谱标记(TMT)定量蛋白质组学技术,对纳米包装和普通聚乙烯包装的双孢蘑菇贮藏期间的差异表达蛋白进行分析,进一步探究纳米包装保鲜双孢蘑菇的作用机制。【方法】以双孢蘑菇为研究对象,用纳米包装对其进行保鲜,并以普通聚乙烯包装作为对照。对贮藏期间双孢蘑菇进行蛋白提取和胰蛋白酶解,并通过TMT标记及液相色谱串联质谱检测,筛选出差异表达蛋白,结合生物信息学分析,研究差异蛋白所参与的主要代谢途径,同时利用实时荧光定量聚合酶链式反应(qPCR)技术,在基因层面验证差异蛋白的表达水平。【结果】纳米包装有效维持了双孢蘑菇的外观品质,并且延缓了细胞膜透性的增加。随着贮藏时间的增加,两组包装的差异蛋白数目增多,在贮藏中期(6 d)和贮藏末期(10 d),差异蛋白分别达到62个和148个,其中纳米包装和普通包装有共同差异蛋白22个。结合生物信息学分析,发现这些差异蛋白主要与能量代谢和脂代谢等功能途径相关。对脂代谢途径进行重点分析,结果显示纳米包装对双孢蘑菇的膜脂代谢具有调控作用,相较于普通包装组,纳米包装组的脂肪酸合成酶、磷酸胆碱孢苷酰转移酶和磷脂酸磷酸酯酶呈上调趋势,同时下调了膜脂降解关键酶如磷脂酶D和脂肪酶的活性;从基因水平上来看,编码这些蛋白的基因表达与组学结果相一致。【结论】利用TMT定量蛋白质组学技术,能对不同包装双孢蘑菇贮藏期间的差异蛋白进行筛选和分析。纳米包装调节了双孢蘑菇的膜脂代谢,抑制了膜脂降解相关酶的表达,有效延缓了细胞膜透性的增加,维持了细胞膜结构和功能,进而延缓双孢蘑菇贮藏期间的品质劣变。
纳米包装;双孢蘑菇;贮藏保鲜;蛋白质组学
0 引言
【研究意义】双孢蘑菇(),也称为白蘑菇,是世界上种植和消费最广泛的食用蘑菇之一,不仅味道鲜美,而且营养价值丰富[1]。但由于双孢蘑菇采后代谢旺盛,容易导致褐变、腐烂等现象[2]。传统的食用菌保鲜方式有低温冷藏、气调包装、涂膜保鲜等[3],但由于成本、安全和保鲜效果不理想等原因,存在着一定的局限性。而纳米包装作为一种新型包装材料,在防雾、抑菌、清除乙烯等方面具有明显优势[4],笔者团队前期研制了一种纳米材料包装袋,相比于普通商业包装袋,纳米包装能抑制食用菌的呼吸速率,减少活性氧积累,有效地缓解了食用菌在贮藏期间的品质劣变[5-7]。然而,对于纳米包装保鲜双孢蘑菇的作用机制,尚未有更深入的研究,因此,利用蛋白质组学技术进一步探究双孢蘑菇采后的代谢规律,对丰富食用菌保鲜体系研究具有重要意义。【前人研究进展】目前,高通量蛋白质组学分析技术作为一种高效、准确的蛋白质分离鉴定手段[8],已成功应用于对果蔬、农作物、肉制品等的分子过程调控机制研究[9],串联质谱标记(tandem mass tag,TMT)定量蛋白质分析技术便是其中被广泛使用的一种。对于被分离和鉴定后的蛋白质,可通过生物信息学分析对其功能和代谢途径进行研究[10]。从生物学角度来看,基因的表达状态对其相应的蛋白质表达具有调控作用,而通过实时荧光定量聚合酶链式反应(quantitative real-time polymerase chain reaction,qPCR)技术,能够对不同物种的基因表达水平进行有效分析[11]。Fang等[12]利用TMT标记结合液相色谱串联质谱法对不同包装金针菇贮藏期间细胞蛋白组进行分离鉴定,研究其保鲜机制;杨波若等[13]通过TMT蛋白质组学,分析了影响猪肉持水性能的差异蛋白;韩冉等[14]采用TMT蛋白质组学结合生物信息学分析,研究了小麦淀粉糯性变异的机制。【本研究切入点】笔者前期已经证实纳米包装能改善双孢蘑菇的贮藏品质,但大多只停留在单一物质或酶面层,尚未有更深入的机理研究。【拟解决的关键问题】本研究以双孢蘑菇为试验对象,利用TMT定量蛋白质组学技术,对纳米包装保鲜双孢蘑菇的代谢规律进行研究;同时通过生物信息学分析,定位差异蛋白的功能和代谢途径,并从基因水平对差异表达蛋白进行检测,为进一步研究纳米包装对食用菌保鲜的作用机制提供参考。
1 材料与方法
试验于2020年11月至2021年6月在南京财经大学食品科学与工程学院进行。
1.1 试验材料
纳米包装袋的制备参照FANG等[4]的方法。30%纳米银、35%纳米二氧化钛、10%纳米二氧化硅及25%纳米凹凸棒土混合制成纳米粉体;然后将其与低密度聚乙烯、低密度线性聚乙烯、分散剂、润滑剂、偶联剂按15﹕46﹕22﹕10﹕5﹕2的比例混匀制成纳米复合母粒;最后将7.5%纳米复合母粒、8%防雾剂和84.5%聚乙烯塑料颗粒混合加工成长、宽、厚分别为25 cm、25 cm和40 μm的纳米包装薄膜。对照组为不添加纳米复合母粒的普通聚乙烯塑料薄膜。
双孢蘑菇采购自五河众兴菌业科技有限公司。样品运至实验室后,将其放入4℃冰箱预冷12 h。挑选菇体完整、颜色洁白、未开伞、无病虫害和机械损伤、菌盖直径30—40 mm的双孢蘑菇,用利刀削去菇柄泥渍部分,用纳米包装袋和普通聚乙烯膜包装袋进行分装。每袋装入双孢菇(200±5)g,每个处理组和对照组各3袋,置于温度为4℃、相对湿度为90%条件下贮藏。贮藏期为10 d,每隔2 d取样一次,将样品用液氮研磨至粉末状,保存于-80℃超低温冰箱待测,其中,选取第0、6和10天的样品进行TMT定量蛋白质组学分析,每个取样时间点的样品进行3次生物学重复。
1.2 仪器与试剂
仪器:FreeZone 2.5冷冻干燥机,美国Labconco公司;Agilent 1100 series高pH分离液相色谱仪,美国Agilent公司;Q Exactive HF质谱仪,美国Thermo Scientific公司;GL-21M型高速冷冻离心机,上海市离心机械研究所有限公司;DDS-11A型电导率仪,上海盛磁仪器有限公司;HH-4数显恒温水浴锅,常州国华电器有限公司;Chemidocxrs+凝胶成像系统,美国Bio-Rad公司;ELX800酶标仪,美国Bio-Tek公司;7500实时荧光定量PCR系统,美国Thermo Scientific公司。
试剂:甲醇(HPLC级)、乙腈(HPLC级)、蔗糖、氯化钠购自南京化学试剂有限公司;二硫苏糖醇(DTT)、甲酸(HPLC级)、-巯基乙醇、醋酸铵、苯基甲磺酰氟(PMSF)购自上海麦克林生化科技有限公司;胰蛋白酶购于北京华利世科技有限公司;碘乙酰胺、四乙基硼氢化铵(TEAB)、Tris-HCl、EDTA·2Na、柱式真菌总RNA抽提纯化试剂盒购于上海生工生物工程股份有限公司;TMTpro 16标记试剂盒购于美国Thermo Scientific公司;HiScript III RT SuperMix for qPCR(+gDNA wiper)逆转录试剂盒、ChamQ™ SYBR qPCR Master Mix(Low ROX Premixed)qPCR试剂盒购于南京诺唯赞生物科技股份有限公司。
1.3 试验设计
1.3.1 外观品质及相对电导率测定 对第0、2、4、6、8和10天的双孢蘑菇进行拍照观察,记录其贮藏期间的表观变化情况。相对电导率参考Barman等[15]的方法并稍作修改。将双孢蘑菇竖切成厚度为3 mm的薄片,后用打孔器将薄片处理成20块圆片。将圆片放入40 mL蒸馏水中,立即测定悬浮液的电导值,记为P0,静置10 min后,测量电导值P1,最后煮沸10 min后测定电导值P2。相对电导率根据以下公式进行计算:
相对电导率(%)=[(P1-P0)/(P2-P0)]×100
1.3.2 蛋白提取 蛋白的提取参考FANG等[12]的方法并稍作修改。取5 g样品加入10 mL蛋白提取液(0.25 mol·L-1Tris-HCl,pH 8.0,含24% w/v蔗糖、0.1 mol·L-1NaCl、0.04 mol·L-1EDTA·2Na、0.01 mol·L-1DTT)充分混匀。加入10 mL酚-Tris-HCl(0.5 mol·L-1,pH 7.8),4℃静置30 min。样品于4℃,7 100×离心10 min,收集上层。加入5倍体积的预冷0.1 mol·L-1醋酸铵-甲醇溶液,-20℃静置12 h。然后4℃,12 000×离心10 min,收集沉淀。沉淀用5倍体积的预冷甲醇清洗,并于4℃、12 000×离心10 min,收集沉淀后用5倍体积丙酮清洗两遍。将沉淀常温干燥后溶于10 mL裂解缓冲液(50 mmol·L-1tris-HCl,pH 7.2,2% v/v-巯基乙醇和1 mmol·L-1苯基甲磺酰氟),溶解3 h。最后将溶液12 000×离心20 min,收集上清液待用。
1.3.3 胰蛋白酶解 胰蛋白酶解参照Wang等[16]的方法并稍作修改。将100 µg蛋白溶于20 µL TEAB(pH 8.5)中,加入0.5 µL 10 mmol·L-1DTT并在55℃条件下孵育0.5 h。加入碘乙酰胺,使其终浓度为10 mmol·L-1,避光静置15 min。溶液中加入6倍体积的丙酮沉淀蛋白,-20℃静置12 h。溶液于4℃、8 000×离心10 min收集沉淀并使丙酮挥发。沉淀中加入100 µL TEAB(100 mmol·L-1)。最后加入胰蛋白酶(蛋白质﹕胰蛋白酶=50﹕1,w/w)于37℃下消化12 h。酶解后的样品冻干后于-80℃保存备用。
1.3.4 TMT标记 取冻干样品用50 µL TEAB(100 mmol·L-1,pH 8.5)重悬。标记反应根据TMTpro(ThermoScientific)试剂盒说明书进行操作,流程见图1。取TMTpro试剂,平衡至室温,加入20 µL无水乙腈,充分振荡后离心。取10 µL TMTpro到样品中,混匀,静置1 h。每个样品加入5 μL 5%羟胺,孵育15 min。收集样品,冻干后于-80℃保存。
1.3.5 液相色谱串联质谱检测 液相色谱串联质谱检测参考FANG等[17]的方法,并稍作修改。流动相A为0.1%的甲酸溶液,流动相B为含0.1%甲酸的乙腈溶液。流速为0.35 µL∙min-1,梯度洗脱方法:0 min,2% B;1 min,6% B;52 min,35% B;54 min,90% B;60 min,90% B。质谱条件:一级MS质量分辨率设为120 000,质谱扫描设定为全扫描荷质比m/z为350—1 650;所有MS/MS图谱采集使用数据依赖型的正离子模式下的高能碰撞裂解完成,碰撞能量设为32;MS/MS的分辨率设为60 000,动态排除时间设为40 s。
1.3.6 生物信息学分析 将质谱检测数据通过uniprot-database伞菌目数据库进行检索,并运用SEQUEST®搜索引擎的Proteome Discoverer 2.4(美国Thermofisher公司)处理。将数据归一化处理后,利用Gene ontology数据库和KEGG 数据库对差异蛋白的功能和其参与代谢的通路进行分析。
1.3.7 qPCR分析 按照ChamQ SYBR qPCR Master Mix试剂盒操作说明进行qPCR分析。反应体系包括10 µL 2×ChamQ SYBR qPCR Master Mix、2 µL cDNA、0.8 µL引物和7.2 µL Nuclease-free H2O。用于qPCR的引物由上海生工生物有限公司合成,以为内参(表1)。qPCR程序设置见表2。

表1 qPCR引物序列

表2 qPCR条件设置
1.3.8 结果统计与分析 采用2−ΔΔCT法计算基因的相对表达量。使用SPSS 26.0软件进行统计分析,显著性分析采用Duncan test,当<0.05时,差异显著。

图1 TMT定量蛋白质组学分析不同包装双孢蘑菇代谢规律流程图
2 结果
2.1 不同包装双孢蘑菇贮藏期间的外观质量及细胞膜透性变化
双孢蘑菇的表观品质是影响消费者购买的重要因素,也是评判双孢蘑菇质量优劣与否的基础指标,能在一定程度上反映出蘑菇的新鲜程度。如图2-A所示,新鲜双孢蘑菇洁白、菌盖表面无破损,但随着贮藏时间的增加,双孢蘑菇的外观品质逐渐降低,其中,普通包装组从4 d开始,逐渐表现出较为明显的褐变现象,当贮藏10 d时,已经呈现出较为严重的表观衰败,并伴有菌盖破膜和水渍状斑块。而纳米包装组则保持了较好的外观品质,尤其到了贮藏末期,纳米包装组表现出的褐变程度要明显好于普通包装组。
细胞膜透性是评价膜系统完整性的一个重要指标,一般而言,细胞膜透性越大,细胞的损伤程度就越严重;此外,它也在一定程度上反映了果蔬的老化程度[18]。细胞膜透性通常用相对电导率(%)表示,如图2-B所示,两组包装双孢蘑菇在贮藏期间的细胞膜透性不断增大,而相较于普通包装组,纳米包装组的增长趋势较为平缓。其中,纳米包装组第6、8和10天的细胞膜透性分别为15.3、16.9和19.3,显著低于对照组的18.5、22.0和26.9。
2.2 差异表达蛋白筛选
使用伞菌目数据库uniprot-database进行蛋白数据检索,Proteome Discover 2.4进行数据处理。根据数据库检索得到的蛋白数据,按照Score Sequest HT>0、Unique Peptides≥1的标准筛选可信蛋白,共鉴定出可信蛋白3 099个。当差异倍数FC>1.5或FC<0.8,且<0.05时,蛋白被认为有显著差异。由图3可知,不同包装处理的双孢蘑菇在贮藏期间的蛋白表达具有差异性。由图3-A可知,在贮藏6 d时,有差异表达蛋白62个,其中,与普通包装组相比,纳米包装组上调蛋白39个,下调蛋白23个;在贮藏10 d时,差异表达蛋白148个,其中纳米包装相较于普通包装组分别上调和下调蛋白91个和57个(图3-B)。双孢蘑菇生理代谢旺盛,采摘后蒸腾作用和呼吸作用不断增强,极易引发其品质的劣变[19-20]。因此,由于双孢蘑菇旺盛的生理代谢,随着贮藏时间的延长,两组包装差异蛋白数不断增加,说明不同包装对双孢蘑菇采后贮藏的影响不断增大。

*表示处理间差异显著(P<0.05)。下同 *indicate sinificant difference (P<0.05). The same as below

A:贮藏6 d时纳米包装组和普通包装组差异表达蛋白;B:贮藏10 d时纳米包装组和普通包装组差异表达蛋白
2.3 差异表达蛋白统计分析
火山图可更直观地展示样本间差异表达物的分布情况,常被用于蛋白质组学和转录组学等的测序结果研究[21]。如图4所示,每个点即为一个蛋白,蓝色的点表示下调表达蛋白,红色的点为上调表达蛋白,灰色点则为非显著差异表达蛋白。其横坐标log2(FC)距0点越远则表示蛋白差异越大。因此,由图4-A与图4-B可知,随着贮藏时间的延长,在差异表达蛋白数量增加的同时,其显著性也随之增加,这可能是由于纳米包装组与普通包装组双孢蘑菇间的代谢差异性不断增大所致。另外,通过对同一包装组的不同贮藏期与CK组新鲜双孢蘑菇的分析,如图4-C韦恩图所示,6 d与10 d的纳米包装组相较于新鲜样品有101个相同的差异表达蛋白,而6 d与10 d的普通包装组与新鲜样品比有145个相同的差异表达蛋白,这表明不同包装处理组对双孢蘑菇贮藏期间的蛋白表达有不同的影响,从而导致不同的保鲜效果。

A:贮藏6 d时纳米包装组和普通包装组差异表达蛋白火山图;B:贮藏10 d时纳米包装组和普通包装组差异表达蛋白火山图;C:贮藏期间纳米包装组和普通包装组差异表达蛋白韦恩图。N:纳米包装组;P:普通包装组;CK:新鲜组
2.4 GO富集与KEGG富集分析
利用GO数据库对纳米包装组和普通包装组的差异表达蛋白进行GO功能富集分析,将差异表达蛋白划分为生物过程(BP)、细胞组分(CC)和分子功能(MF)3大类,分别挑选出每类最为富集的功能节点,以探究不同包装差异蛋白所属的功能区域。如图5-A所示,两组包装差异表达蛋白在生物过程中最为富集的节点分别为:糖代谢过程、翻译过程、蛋白质成熟、细胞内蛋白质转运、三羧酸循环、囊泡介导转运、蛋白质折叠和脂肪酸代谢过程;差异蛋白在细胞组分中最为富集的节点分别为:胞外区、细胞膜、核小体、核仁和内质网膜;差异蛋白在分子功能类别中最为富集的节点分别为:金属离子结合、RNA结合、水解酶活性、氧化还原酶活性、FAD绑定、NAD绑定、核糖体结构成分、翻译起始因子活性、蛋白激酶调节活性和GTP绑定。

图5 不同包装组双孢蘑菇贮藏期间差异表达蛋白的GO(A)与KEGG(B)分析
KEGG是系统分析蛋白质在细胞中代谢途径的主要公共数据库,参与同一途径的蛋白质共同执行它们的生物学功能[22-23],利用KEGG数据库对两组包装差异表达蛋白进行通路分析,从而进一步探究差异蛋白的生物学功能。选取两个包装组中差异表达蛋白富集最为显著的20条KEGG通路,结果如图5-B所示,差异蛋白的主要代谢途径有柠檬酸循环、丙酮酸代谢、酮体合成与降解、脂肪酸生物合成和脂肪酸降解等。
对两组包装差异表达蛋白质进一步分析,共筛选出31个差异蛋白(表3)。在这些蛋白中,XP_ 006456433.1、XP_006457863.1在贮藏6 d时下调,10 d时无显著变化,其中,XP_006457863.1尚未表征;而XP_006456433.1主要参与催化葡萄糖生成过氧化氢,最后使木质素发生降解的过程[24],6 d时纳米包装下调了XP_006456433.1的表达进而延缓了双孢蘑菇的木质化过程,10 d时可能由于两组包装木质素累积进程相近而变化不显著。此外,XP_006453957.1、XP_006458170.1在6 d时无显著变化,10 d时出现下调,以及XP_006456371.1、XP_006463403.1等5个蛋白在6 d时变化不显著,在10 d时发生上调。另外,有XP_006462684.1、XP_006453790.1等22个蛋白是两组包装在贮藏6 d与10 d时的共同差异蛋白,其中,有上调蛋白12个,下调蛋白10个。
2.5 生物信息学与通路分析
通过对差异蛋白的生物信息学分析,发现纳米包装对双孢蘑菇的能量代谢具有调控作用,在贮藏期间,相较于普通包装,纳米包装组的双孢蘑菇中ATP激酶(XP_006454339.1)与ATP结合蛋白(XP_006455684.1)均得到上调,延缓了纳米组蘑菇的能量亏损。此外,线粒体作为真核生物产生能量的主要场所[25],纳米包装组中的线粒体膜转运蛋白(XP_006454789.1)也呈上调趋势,从而维持线粒体电子传递链的稳态,延缓了双孢蘑菇的衰老过程。另外,纳米包装对双孢蘑菇细胞膜水平同样具有调控作用,纳米包装组中的XP_006456371.1、XP_006454643.1和XP_006454671.1等膜组分蛋白均为上调。具体的膜脂代谢调控途径如图6所示,相较于普通包装组,纳米组双孢蘑菇中的XP_006459152.1柠檬酸合酶(citrate synthase,CS)、XP_006453790.1脂肪酶(lipase,LPS)和XP_006462684.1磷脂酶D(phospholipase D,PLD)呈现下调,并且随着贮藏时间的延长,它们在10 d时相比于6 d时下调更为显著;而XP_006459063.1脂肪酸合酶(fatty acid synthase,FAS)、XP_006453826.1磷酸胆碱孢苷酰转移酶(phosphocholine cytidylyltransferase,CTP)和XP_006456787.1磷脂酸磷酸酯酶(phosphatidate phosphatase,PPase)则呈现上调趋势,相较于6 d,它们在贮藏末期(10 d)上调趋势更显著。

红色表示上调,绿色表示下调 Red means up-regulation, and green means down-regulation
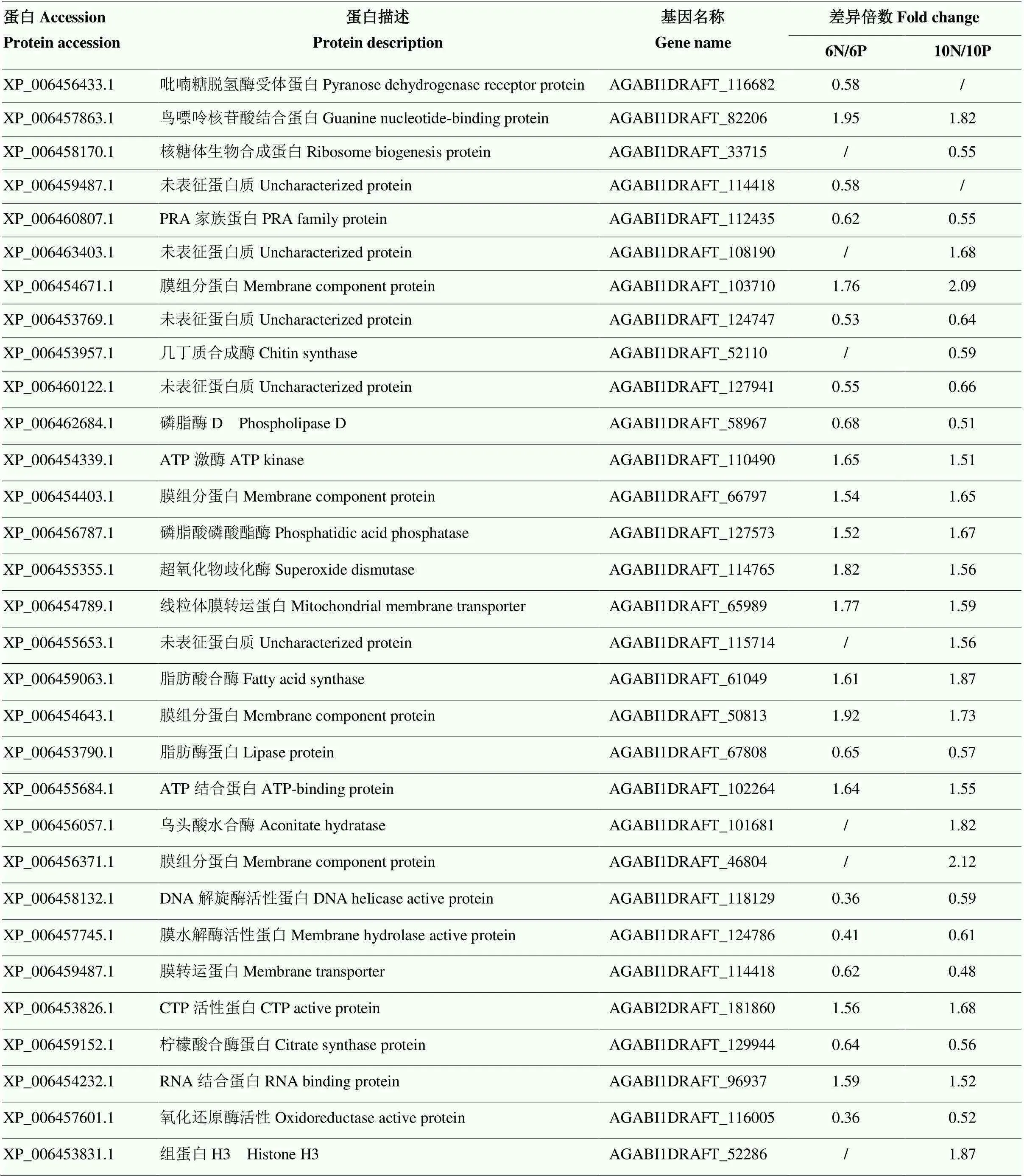
表3 不同包装组双孢蘑菇贮藏期间主要差异表达蛋白
“/”表示该蛋白质无显著性变化 “/” means that the protein has no significant change
2.6 膜脂代谢差异表达蛋白的基因验证
通过对以上通路的分析,为了进一步研究双孢蘑菇贮藏期间膜脂代谢差异蛋白在基因水平的表达变化,对编码CS、FAS、CTP、PLD、PPase和LPS的基因进行qPCR验证。如图7所示,的相对表达呈现出先降低后上升,又再降低的趋势,这种变化可能是由于双孢蘑菇在贮藏过程中脂代谢波动所致[26],而纳米包装组的表达始终显著低于普通包装组,这有利于维持贮藏过程中的脂类平衡;对于,纳米包装组的表达为先下降后略有上升,而普通包装组总体上为下降趋势,这表明纳米包装组可通过维持脂肪酸的合成能力来延缓双孢蘑菇在贮藏过程中的脂肪酸降解;的表达总体呈下降趋势,其中纳米包装组在6、8和10 d的表达要显著高于普通包装组,这有助于维持细胞膜的结构;两组包装双孢蘑菇的表达为先上升后下降趋势,并且2、4、6和10 d时纳米包装组的表达水平要显著低于普通包装组,表明纳米包装抑制了的表达,进而延缓了细胞膜的水解;的表达也呈现出先上升后下降的趋势,双孢蘑菇通过提高的表达来延缓脂类的损失程度,其中纳米包装组的表达要高于普通包装组,且在2、6和10 d达到显著水平;在普通包装组中为先上升后下降的趋势,在6 d达到表达高峰,而纳米包装组则在2 d时有最高表达,而后降低,且在4、6、8和10 d显著低于普通包装组,表明纳米包装抑制了的表达,进而缓解膜脂的降解。以上差异蛋白在基因水平的表达与组学结果相符,这再次表明,纳米包装可以通过调控以上蛋白的基因表达,进而影响双孢蘑菇的膜脂代谢途径,以实现延缓其衰老和品质劣变的目的。

图7 不同包装组双孢蘑菇贮藏期间膜脂代谢途径差异蛋白的qPCR分析
3 讨论
3.1 纳米包装提升双孢蘑菇采后的贮藏品质
新鲜的双孢蘑菇水分含量高、组织细嫩,且生理代谢旺盛[19],尤其是采收后,蒸腾作用与呼吸代谢增强,使子实体老化加剧,极易发生褐变、失水萎蔫和破膜开伞等现象,最终导致品质劣变[20]。本研究结果表明,与普通包装相比,纳米包装有效维持了双孢蘑菇采后的表观品质,在整个贮藏期间未发生明显的褐变现象,笔者课题组前期的研究表明,纳米包装还能保持双孢蘑菇的营养品质,例如维持总糖和蛋白质的含量[27],从而延缓其采后衰老进程。双孢蘑菇在贮藏过程中,膜脂过氧化程度不断加剧,导致细胞膜透性增大,严重破坏了细胞膜的结构与功能,但纳米包装能延缓细胞膜透性的增加,丙二醛(MDA)是反映膜脂过氧化水平的重要指标,在前期工作中,已经对两组包装双孢蘑菇的MDA含量变化进行了研究,发现MDA含量随贮藏时间的增加而升高,纳米包装组的MDA含量始终低于普通包装组[27],这也进一步解释了本研究中纳米组细胞膜透性较低的原因,说明纳米包装有效缓解了双孢蘑菇贮藏期间的膜脂过氧化,进而延缓了细胞膜透性的增加,减少了细胞膜损伤,更有利于提升双孢蘑菇的贮藏品质。
3.2 纳米包装调控双孢蘑菇的能量代谢和膜脂代谢
在贮藏过程中,两组包装的差异蛋白数量不断增加,它们主要与能量代谢和脂代谢等途径相关。ATP作为细胞最直接的能量供应来源,对果蔬采后的正常生理代谢功能起着重要作用,而贮藏期间的能量不足则会加速新鲜蘑菇质量的恶化[28]。在笔者前期的研究中,已经证明纳米包装对食用菌贮藏期间能量代谢的调控作用[4,6,27,29-30],采后4 d时纳米包装组双孢蘑菇的呼吸速率比普通包装组低9%;此外,纳米包装还缓解了其能荷的下降,在贮藏末期,纳米组的双孢蘑菇仍保持了较高的能荷水平。因此,本研究也再一次表明,纳米包装可以调控双孢蘑菇的能量代谢,维持了其贮藏过程中的能量状态,有助于保持双孢蘑菇的正常生理功能和缓解采后衰老。
细胞膜将酶与底物隔开使细胞形成区室化分布,这对保持细胞正常的生理功能至关重要,研究表明,细胞膜损伤是导致果蔬品质劣变的主要因素之一[31-32]。YAO等[33]通过甘氨酸甜菜碱处理西葫芦,减少了膜脂过氧化程度,减轻了其冷害的发生;LIU等[34]发现,低温贮藏荔枝果实,可降低其膜透性和膜磷脂的降解以延长贮藏时间。本研究发现纳米包装对双孢蘑菇的膜脂代谢途径同样具有调控作用(图6)。三羧酸循环是细胞内蛋白质、脂肪和糖相互关联及转化的枢纽,而乙酰辅酶A在这些代谢过程中起着关键作用[35]。一方面,CS作为三羧酸循环限速酶,可以催化乙酰辅酶A与草酰乙酸的缩合产物生成柠檬酸,后者再进入三羧酸循环[36];另一方面,乙酰辅酶A和丙二酰辅酶A又在FAS系统的催化下生成脂肪酸进入脂代谢[37]。而纳米包装组抑制了的表达,提高了的表达,减少双孢蘑菇能量损耗,同时也有助于维持双孢蘑菇体内脂质的平衡[38]。磷脂是细胞膜系统的主要成分,在细胞结构、分子信号转导和膜流动性中起关键作用,其中,磷脂酰胆碱(phosphatidylcholine,PC)和磷脂酰乙醇胺(phosphatidylethanolamine,PE)是维持细胞膜结构的主要磷脂[39],PC通过CDP-胆碱途径合成,而CTP是其主要的限速酶[40]。在贮藏期间,相较于普通组,纳米组的呈不断上调趋势,更有利于PC的合成,从而维持膜结构;PLD作为磷脂代谢的起始酶,可以水解PC、PE等膜磷脂组分[41],LPS也是参与磷脂水解的关键酶,可将甘油二酯转化为游离脂肪酸[34],这种由细胞膜磷脂水解所造成的膜结构破坏,会最终导致膜功能的丧失,而纳米包装抑制了及的表达,从而减少了双孢蘑菇的膜损伤。根据前期的研究,这种抑制作用可能是由于纳米包装调控了贮藏环境的气体组成,加上纳米材料具有抑菌和清除乙烯的作用,进而改善双孢蘑菇的内源代谢和减少外部侵染所致[42]。另外,PLD水解过程会产生磷脂酸(phosphatidic acid,PA),PA的积累则会加剧膜脂过氧化程度[43],而PPase作为磷酸酯酶的一种,能催化PA的水解生成二酰甘油,二酰甘油则是三酰甘油合成的关键前体物质,有助于脂肪的合成[44]。因此,在纳米包装组双孢蘑菇的高表达有效地维持了细胞膜结构和功能。从基因层面上看,这些差异蛋白的表达趋势与组学结果相符,表明纳米包装能调控双孢蘑菇的膜脂代谢,延缓细胞膜损伤,进而维持细胞膜结构与功能,最终保持双孢蘑菇的贮藏品质。
4 结论
随着贮藏时间的增加,两组包装的差异表达蛋白数目不断增大,到贮藏10 d时,共鉴定差异表达蛋白148个。这些蛋白主要参与到能量代谢及脂代谢等代谢途径。其中,纳米包装通过抑制、和的相对表达,以及提高、和的表达量,可延缓双孢蘑菇在贮藏过程中的细胞膜损伤,有效缓解了其采后品质劣变。
[1] BAN Z J, LI L, GUAN J F, FENG J H, WU M Y, XU X M, LI J. Modified atmosphere packaging (MAP) and coating for improving preservation of whole and sliced. Journal of Food Science and Technology, 2014, 51(12): 3894-3901. doi: 10.1007/ s13197-013-0935-9.
[2] DOKHANIEH A Y, AGHDAM M S. Postharvest browning alleviation ofusing salicylic acid treatment. Scientia Horticulturae, 2016, 207: 146-151. doi: 10.1016/j.scienta.2016.05. 025.
[3] 徐静, 谷艳杰. 不同包装方式对金针菇冷藏保鲜效果的影响. 山东农业工程学院学报, 2016, 33(12): 122-124.
XU J, GU Y J. The effect of different packaging methods onrefrigerated preservation. Journal of Shandong Agriculture and Engineering University, 2016, 33(12): 122-124. (in Chinese)
[4] FANG D L, YANG W J, KIMATU B M, AN X X, HU Q H, ZHAO L Y. Effect of nanocomposite packaging on postharvest quality and reactive oxygen species metabolism of mushrooms (). Postharvest Biology and Technology, 2016, 119: 49-57. doi: 10.1016/j.postharvbio.2016.04.012.
[5] YANG W J, SHI C, HU Q H, WU Y Y, FANG D L, PEI F, MARIGA A M. Nanocomposite packaging regulate respiration and energy metabolism in. Postharvest Biology and Technology, 2019, 151: 119-126. doi: 10.1016/j.postharvbio.2019.02. 003.
[6] 李志啸, 杨文建, 方东路, 裴斐, 马宁, 方勇, 赵立艳, 安辛欣, 胡秋辉. 纳米包装材料对双孢菇细胞壁代谢及品质的影响. 食品科学, 2016, 37(6): 248-253.
LI Z X, YANG W J, FANG D L, PEI F, MA N, FANG Y, ZHAO L Y, AN X X, HU Q H. Effects of nano-packaging on cell wall metabolism and postharvest qualities of. Food Science, 2016, 37(6): 248-253. (in Chinese)
[7] ZUO C Z, HU Q H, SU A X, XU H, LI X T, MARIGA A M, YANG W J. Nanocomposite packaging delays lignification ofby regulating phenylpropanoid pathway and mitochondrial reactive oxygen species metabolisms. Postharvest Biology and Technology, 2021, 171: 111360. doi: 10.1016/j.postharvbio.2020. 111360.
[8] 剧柠, 苟萌, 张彤彤. 蛋白质组学技术及其在乳及乳制品中的应用研究进展. 食品与发酵工业, 2021, 47(3): 245-251.
JU N, GOU M, ZHANG T T. Advances in proteomics and its application in milk and dairy products. Food and Fermentation Industries, 2021, 47(3): 245-251. (in Chinese)
[9] WU X Q, JIANG L, YU M L, AN X J, MA R J, YU Z F. Proteomic analysis of changes in mitochondrial protein expression during peach fruit ripening and senescence. Journal of Proteomics, 2016, 147: 197-211. doi: 10.1016/j.jprot.2016.06.005.
[10] 陈鹏, 周瑞阳, 蒋利和. 线粒体蛋白质组学技术及其在植物细胞质雄性不育机理研究中的应用. 南方农业学报, 2011, 42(4): 353-357. doi: 10.3969/j.issn.2095-1191.2011.04.002.
CEHN P, ZHOU R Y, JIANG L H. Progress in mitochondrial proteomics technology and its applications in studying plant cytoplasmic male sterility. Journal of Southern Agriculture, 2011, 42(4): 353-357. doi: 10.3969/j.issn.2095-1191.2011.04.002. (in Chinese)
[11] 赵巧灵. 金枪鱼在冷藏过程中鲜度变化及差异蛋白质组学研究[D]. 杭州: 浙江工商大学, 2015.
ZHAO Q L. Freshness variation and differential proteome analysis of tuna () during refrigerated storage[D]. Hangzhou: Zhejiang Gongshang University, 2015. (in Chinese)
[12] FANG D L, YANG W J, DENG Z L, AN X X, ZHAO L Y, HU Q H. Proteomic investigation of metabolic changes of mushroom () packaged with nanocomposite material during cold storage. Journal of Agricultural and Food Chemistry, 2017, 65(47): 10368-10381. doi: 10.1021/acs.jafc.7b04393.
[13] 杨波若, 李华健, 苏娅宁, 李霞, 瞿静, 陈韬. 基于微观结构和蛋白质组学分析影响猪肉持水性的差异蛋白. 食品工业科技, 2021, 42(7): 136-144.
YANG B R, LI H J, SU Y N, LI X, QU J, CHEN T. Analysis of different proteins affecting water holding capacity of pork based on microstructure and proteomics. Science and Technology of Food Industry, 2021, 42(7): 136-144. (in Chinese)
[14] 韩冉, 毛凤鑫, 程敦公, 李豪圣, 刘建军, 曹新有, 宋健民, 辛明明, 郭军, 刘成, 刘爱峰. 全糯性小麦济糯116的籽粒蛋白质组学分析. 山东农业科学, 2021, 53(5): 1-7.
HAN R, MAO F X, CHENG D G, LI H S, LIU J J, CAO X Y, SONG J M, XIN M M, GUO J, LIU C, LIU A F. Proteomic analysis of waxy wheat Jinuo 116. Shandong Agricultural Sciences, 2021, 53(5): 1-7. (in Chinese)
[15] BARMAN K, ASREY R, PAL R K, JHA S K, BHATIA K. Post-harvest nitric oxide treatment reduces chilling injury and enhances the shelf-life of mango (L.) fruit during low-temperature storage. The Journal of Horticultural Science and Biotechnology, 2014, 89(3): 253-260. doi: 10.1080/14620316.2014. 11513076.
[16] WANG C, CHU J J, FU L L, WANG Y B, ZHAO F, ZHOU D Q. iTRAQ-based quantitative proteomics reveals the biochemical mechanism of cold stress adaption of razor clam during controlled freezing-point storage. Food Chemistry, 2018, 247: 73-80. doi: 10.1016/j.foodchem.2017.12.004.
[17] FANG D L, ZHENG Z, MA N, YANG W J, DAI C, ZHAO M W, DENG Z L, HU Q H, ZHAO L Y. Label-free proteomic quantification of packagedduring commercial storage. Postharvest Biology and Technology, 2020, 169: 111312. doi: 10.1016/j.postharvbio.2020.111312.
[18] HU Y H, CHEN C M, XU L, CUI Y, YU X Y, GAO H J, WANG Q, LIU K, SHI Y, CHEN Q X. Postharvest application of 4-methoxy cinnamic acid for extending the shelf life of mushroom (). Postharvest Biology and Technology, 2015, 104: 33-41. doi: 10.1016/j.postharvbio.2015.03.007.
[19] ZHANG K X, PU Y Y, SUN D W. Recent advances in quality preservation of postharvest mushrooms (): A review. Trends in Food Science & Technology, 2018, 78: 72-82. doi: 10.1016/j.tifs.2018.05.012.
[20] KAMAL A S M, KHAIR A, BEGUM F, CHOWDHURY K, KARIM R. Effect of respiratory gases (O2; CO2) on shelf-life of fresh oyster mushrooms packaged with different sealable polymeric materials. Bangladesh Journal of Scientific and Industrial Research, 2015, 50(3): 205-210. doi: 10.3329/bjsir.v50i3.25587.
[21] LI W T, FREUDENBERG J, SUH Y J, YANG Y N. Using volcano plots and regularized-Chi statistics in genetic association studies. Computational Biology and Chemistry, 2014, 48: 77-83. doi: 10.1016/j.compbiolchem.2013.02.003.
[22] KANEHISA M, ARAKI M, GOTO S, HATTORI M, HIRAKAWA M, ITOH M, KATAYAMA T, KAWASHIMA S, OKUDA S, TOKIMATSU T, YAMANISHI Y. KEGG for linking genomes to life and the environment. Nucleic Acids Research, 2007, 36(suppl_1): D480-D484. doi: 10.1093/nar/gkm882.
[23] ZHU Z, ZHANG Y B, LIU J, CHEN Y L, ZHANG X J. Exploring the effects of selenium treatment on the nutritional quality of tomato fruit. Food Chemistry, 2018, 252: 9-15. doi: 10.1016/j.foodchem. 2018.01.064.
[24] ZHU N, LIU J W, YANG J S, LIN Y J, YANG Y, JI L, LI M, YUAN H L. Comparative analysis of the secretomes ofand other wood-decay basidiomycetes during solid-state fermentation reveals its unique lignocellulose-degrading enzyme system. Biotechnology for Biofuels, 2016, 9: 42. doi: 10.1186/ s13068-016-0461-x.
[25] MARTEZ J, ROLAND S, ANDREAS S, PETER K, RONALD E. Perfil de eficacia y seguridad de Echinacea purpurea en la prevención de episodios de resfriado común: Estudio clínico aleatorizado, doble ciego y controlado con placebo. Revista de Fitoterapia, 2013, 13(24): 125-135. doi: 10.1002/jsfa.
[26] 葛亚东, 潘蔚, 汪劼, 朱国萍. 柠檬酸合酶的分子生物学研究进展. 生物学杂志, 2010, 27(3): 59-62.
GE Y D, PAN W, WANG J, ZHU G P. Advances in molecular biology of citrate synthase. Journal of Biology, 2010, 27(3): 59-62. (in Chinese)
[27] 杨文建, 单楠, 杨芹, 方勇, 赵立艳, 安辛欣, 马宁, 刘音宏, 辛志宏, 胡秋辉. 纳米包装材料延长双孢蘑菇贮藏品质的作用. 中国农业科学, 2012, 45(24): 5065-5072.
YANG W J, SHAN N, YANG Q, FANG Y, ZHAO L Y, AN X X, MA N, LIU Y H, XIN Z H, HU Q H. Effect of nano-packing material on improvement of preservation quality of. Scientia Agricultura Sinica, 2012, 45(24): 5065-5072. (in Chinese)
[28] LI P Y, YIN F, SONG L J, ZHENG X L. Alleviation of chilling injury in tomato fruit by exogenous application of oxalic acid. Food Chemistry, 2016, 202: 125-132. doi: 10.1016/j.foodchem.2016.01.142.
[29] 马宁, 王鹤潼, 方东路, 赵立艳, 杨文建, 裴斐, 胡秋辉. 基于线粒体能量代谢途径的金针菇采后纳米包装保鲜机制. 中国农业科学, 2020, 53(16): 3356-3371.
MA N, WANG H T, FANG D L, ZHAO L Y, YANG W J, PEI F, HU Q H. Nano-packaging preservative mechanism ofafter harvest based on mitochondrial energy status pathways. Scientia Agricultura Sinica, 2020, 53(16): 3356-3371. (in Chinese)
[30] WU Y Y, HU Q H, LI Z X, PEI F, MUGAMBI M A, YANG W J. Effect of nanocomposite-based packaging on microstructure and energy metabolism of. Food Chemistry, 2019, 276: 790-796. doi: 10.1016/j.foodchem.2018.10.088.
[31] GAO H, CHAI H K, CHENG N, CAO W. Effects of 24- epibrassinolide on enzymatic browning and antioxidant activity of fresh-cut lotus root slices. Food Chemistry, 2017, 217: 45-51. doi: 10.1016/j.foodchem.2016.08.063.
[32] CHEN Y H, SUN J Z, LIN H T, LIN M S, LIN Y F, WANG H, HUNG Y C. Salicylic acid treatment suppresses Phomopsis longanae Chi-induced disease development of postharvest longan fruit by modulating membrane lipid metabolism. Postharvest Biology and Technology, 2020, 164: 111168. doi: 10.1016/j.postharvbio.2020. 111168.
[33] YAO W S, XU T T, FAROOQ S U, JIN P, ZHENG Y H. Glycine betaine treatment alleviates chilling injury in zucchini fruit () by modulating antioxidant enzymes and membrane fatty acid metabolism. Postharvest Biology and Technology, 2018, 144: 20-28. doi: 10.1016/j.postharvbio.2018.05.007.
[34] LIU H, SONG L L, YOU Y L, LI Y B, DUAN X W, JIANG Y M, JOYCE D C, ASHRAF M, LU W J. Cold storage duration affects litchi fruit quality, membrane permeability, enzyme activities and energy charge during shelf time at ambient temperature. Postharvest Biology and Technology, 2011, 60: 24-30. doi: 10.1016/j.postharvbio. 2010.11.008.
[35] 陈牧, 刘锐, 翁屹. 三羧酸循环的发现与启示. 医学与哲学, 2012, 33(1): 71-73.
CHEN M, LIU R, WENG Y. The discovery and revelation of tricarboxylic acid cycle. Medicine and Philosophy, 2012, 33(1): 71-73. (in Chinese)
[36] WIENKOOP S, SAALBACH G. Proteome analysis. Novel proteins identified at the peribacteroid membrane fromroot nodules. Plant Physiology, 2003, 131(3): 1080-1090. doi: 10.1104/pp. 102.015362.
[37] FERNANDEZ-MOYA R, LEBER C, CARDENAS J, DA SILVA N A. Functional replacement of thefatty acid synthase with a bacterial type II system allows flexible product profiles. Biotechnology and Bioengineering, 2015, 112(12): 2618-2623. doi: 10.1002/bit.25679.
[38] LIN Y X, LIN Y F, CHEN Y H, WANG H, SHI J, LIN H T. Hydrogen peroxide induced changes in energy status and respiration metabolism of harvested longan fruit in relation to pericarp browning. Journal of Agricultural and Food Chemistry, 2016, 64(22): 4627-4632. doi: 10.1021/acs.jafc.6b01430.
[39] ZHOU Y C, PAN X P, QU H X, UNDERHILL S J R. Low temperature alters plasma membrane lipid composition and ATPase activity of pineapple fruit during blackheart development. Journal of Bioenergetics and Biomembranes, 2014, 46(1): 59-69. doi: 10.1007/ s10863-013-9538-4.
[40] BANCHIO C, SCHANG L M, VANCE D E. Activation of CTP: phosphocholine cytidylyltransferase α expression during the S phase of the cell cycle is mediated by the transcription factor Sp1. The Journal of Biological Chemistry, 2003, 278(34): 32457-32464. doi: 10.1074/jbc.m304810200.
[41] HONG Y Y, ZHAO J, GUO L, KIM S C, DENG X J, WANG G L, ZHANG G Y, LI M Y, WANG X M. Plant phospholipases D and C and their diverse functions in stress responses. Progress in Lipid Research, 2016, 62: 55-74. doi: 10.1016/j.plipres.2016.01.002.
[42] 杨燕婷, 杨芹, 杨方美, 辛志宏, 赵立艳, 郁志芳, 胡秋辉. 纳米包装材料对金针菇的保鲜作用. 中国农业科学, 2009, 42(9): 3250-3258.
YANG Y T, YANG Q, YANG F M, XIN Z H, ZHAO L Y, YU Z F, HU Q H. Effect of nano-packaging material on quality of. Scientia Agricultura Sinica, 2009, 42(9): 3250-3258. (in Chinese)
[43] KARAKURT Y, HUBER D J. Ethylene-induced gene expression, enzyme activities, and water soaking in immature and ripe watermelon () fruit. Journal of Plant Physiology, 2004, 161(4): 381-388. doi: 10.1078/0176-1617-01221.
[44] 李一路, 张晴晴, 胡卫芹, 屈钢, 洪月云. 磷脂酸磷酸酶在脂质代谢和信号转导中的作用及其调控. 植物生理学报, 2017, 53(6): 897-904.
LI Y L, ZHANG Q Q, HU W Q, QU G, HONG Y Y. Roles and regulation of phosphatidic acid phosphatase in lipid metabolism and signaling. Plant Physiology Journal, 2017, 53(6): 897-904. (in Chinese)
Physiological Metabolic Role of Nanocomposite PackagedDuring Postharvest Cold Storage Analyzed by TMT-Based Quantitative Proteomics

1College of Food Science and Engineering, Nanjing University of Finance and Economics/Jiangsu Collaborative Innovation Center for Modern Grain Circulation and Safety/Key Laboratory of Grains and Oils Quality Control and Processing, Nanjing 210023;2College of Light Industry and Food Engineering, Nanjing Forestry University, Nanjing 210037
【Background】() is prone to quality deterioration, such as umbrella opening, water loss, and browning after harvest, which seriously affects the storage quality and commercial value. Our previous research has confirmed that the nanocomposite packaging material (Nano-PM) could effectively delay the postharvest quality deterioration of, but the preservation mechanism is still unclear. 【Objective】In this study, the differentially expressed proteins ofin Nano-PM and polyethylene packaging material (Normal-PM) during storage were analyzed by Tandem Mass Tag (TMT) quantitative proteomics technology. The preservation mechanism of Nano-PM onwas further explored. 【Method】was taken as the research object. The Nano-PM was used for the preservation of, and the Normal-PM was used as the control. The protein extraction and trypsin hydrolysis were performed onduring storage. The differentially expressed proteins were screened by TMT labeling and liquid chromatography-tandem mass spectrometry detection. Combined with bioinformatics analysis, the main metabolic pathways involved in differential proteins were studied. Quantitative real-time polymerase chain reaction (qPCR) technology was used to determine the gene expression levels of differential proteins. 【Result】The Nano-PM effectively maintained the appearance quality ofand delayed the increase of cell membrane permeability. The number of differential proteins in two groups increased during storage. In the middle (6 d) and late (10 d) stages of storage, the numbers of differential proteins were 62 and 148, respectively. Among them, 22 differential proteins were common. Combined with bioinformatics analysis, these differential proteins were mainly related to pathways, such as energy metabolism and lipid metabolism. The lipid metabolism pathway was mainly analyzed, and the results showed that the Nano-PM had a regulatory effect on the membrane lipid metabolism of. Compared with the Normal-PM, the protein expression of fatty acid synthase, phosphorylcholine cytidylyltransferase, and phosphatidic acid phosphatase under Nano-PM were up-regulated, while the protein expression of key enzymes in membrane lipid degradation, such as phospholipase D and lipase, were down-regulated. At the gene level, the expression of genes encoding these proteins were consistent with the proteomics results. 【Conclusion】The differential proteins of different packagedduring storage could be screened and analyzed by TMT-based quantitative proteomics technology. Nano-PM regulated the membrane lipid metabolism of, and inhibited the expression of membrane lipid degradation-related enzymes, which effectively delayed the increase in cell membrane permeability of, maintained the structure and function of the cell membrane, and delayed the quality deterioration ofduring storage.
nanocomposite packaging;; preservation; proteomics

10.3864/j.issn.0578-1752.2022.23.013
2022-03-09;
2022-05-25
江苏省自然科学基金(BK20201395)、江苏高校优势学科建设工程资助项目(PAPD)
王朝,E-mail:wangchaonufe@163.com。通信作者马宁,E-mail:maning@nufe.edu.cn
(责任编辑 赵伶俐)
