温度和盐度对扇贝“渤海红”稚贝存活和生长的互作效应
2022-01-26朱战豪姚高友苏晓盈刘晓坤刘付少梅方家熙陈楠生王春德刘志刚
张 元,朱战豪,姚高友,苏晓盈,刘晓坤,刘付少梅,方家熙,陈楠生,王春德,刘志刚
( 1.广东海洋大学 水产学院,广东 湛江 524088; 2.广东省南海经济无脊椎动物健康养殖工程研究中心,广东 湛江 524088; 3.湛江银浪海洋生物技术有限公司,广东 湛江 524022; 4.香港理工大学 应用生物及化学科技学系,香港 999077; 5.西蒙·弗雷泽大学 理学院分子生物和生物化学系,加拿大 不列颠哥伦比亚省 本那比市 V5A 1S6; 6.青岛农业大学 海洋科学与工程学院,山东 青岛 266109 )
紫扇贝(Argopectenpurpuratus)与海湾扇贝北部亚种(A.irradiansirradians)杂交经6代选育后得到扇贝“渤海红”,在第五届全国水产原种和良种审定委员会上通过新品种审定。其壳较薄且颜色鲜艳,为紫红色,具有明显的生长性状优势[1-3],相同养殖条件下,9月龄贝相比于普通海湾扇贝,其体质量与闭壳肌质量增长超过37.0%和49.0%[4]。2013年开始,扇贝“渤海红”的商业养殖辐射到山东省、河北省和辽宁省大部分地区[5],目前为北方扇贝养殖品种之一[6],经济效益显著,推动扇贝养殖业健康持续发展。扇贝“渤海红”的半致死温度为29 ℃[5],2015年广东海洋大学将其与耐高温的粤西养殖品种墨西哥湾扇贝(A.irradiansconcentricus)进行杂交,以期突破高温障碍,在南方进行养殖推广。众所周知,单一因子的海水温度或盐度的变化直接影响养殖对象的存活与生长,也有研究表明,不同试验条件下,其试验结果会反映出各种环境因子的重要程度[7-8],存在协同和互作效应[9-11];其他研究亦指出,各种不同时期的贝类对温度与盐度均有最佳的适应性[11-14]。扇贝“渤海红”经6代选育后,子一代的温度耐受性变宽,可适应我国大部分养殖海域[5],但有关该品种的最适温度、盐度及温度、盐度互作效应目前尚未见报道。
响应曲面分析法(RSM)是利用合理的试验设计并通过试验得到相应数据,采用二次多元回归方程拟合因素与响应值之间的函数关系,通过分析此方程来寻求最优试验参数并解决多变量问题的一种统计方法[15-16]。试验数据经软件分析得到相应的响应曲面,其可直观地反映因素对响应值的影响[17]。此方法试验次数少,在短期内能得到最优化试验条件[18-19],结合中心复合设计方式(CCD)建立响应曲面模型,并对模型进行分析与评价。笔者通过探讨温度和盐度两个重要的环境因子间的互作效应及其对稚贝生长和存活的影响,以确定稚贝的存活与生长的最适温度、盐度组合,为该贝的人工养殖对温度、盐度的需求奠定试验依据,并对扇贝“渤海红”作为亲本培育新品种提供一个合适的暂养温度,有利于对其与墨西哥湾扇贝杂交后代的温度、盐度适应性进行评价。
1 材料与方法
1.1 材料
试验所用“渤海红”稚贝取自湛江银浪海洋生物技术有限公司长洪育苗场,2019年4月1日运回广东海洋大学广东省南海经济无脊椎动物健康养殖工程研究中心实验室暂养3 d。暂养期间海水温度22~26 ℃,盐度27~31,并且持续充气,每日投喂浓缩小球藻(Chlorellasp.)3~4次,每次投喂饵料10~12 mL,保持试验水体饵料密度约为1×104个/mL,每日全量换水1次。
1.2 方法
1.2.1 试验设计
根据预试验结果,采用中心复合设计法和响应曲面分析法,以稚贝的存活率、体质量增长率和壳长增长率这3个为响应值,以温度(16~28 ℃)和盐度(22~34)为试验因子,每个因子取3个水平,各水平的编码值分别为-1、0、1,中心点重复3次,共设计11个组合,整个试验重复3次(表1)。
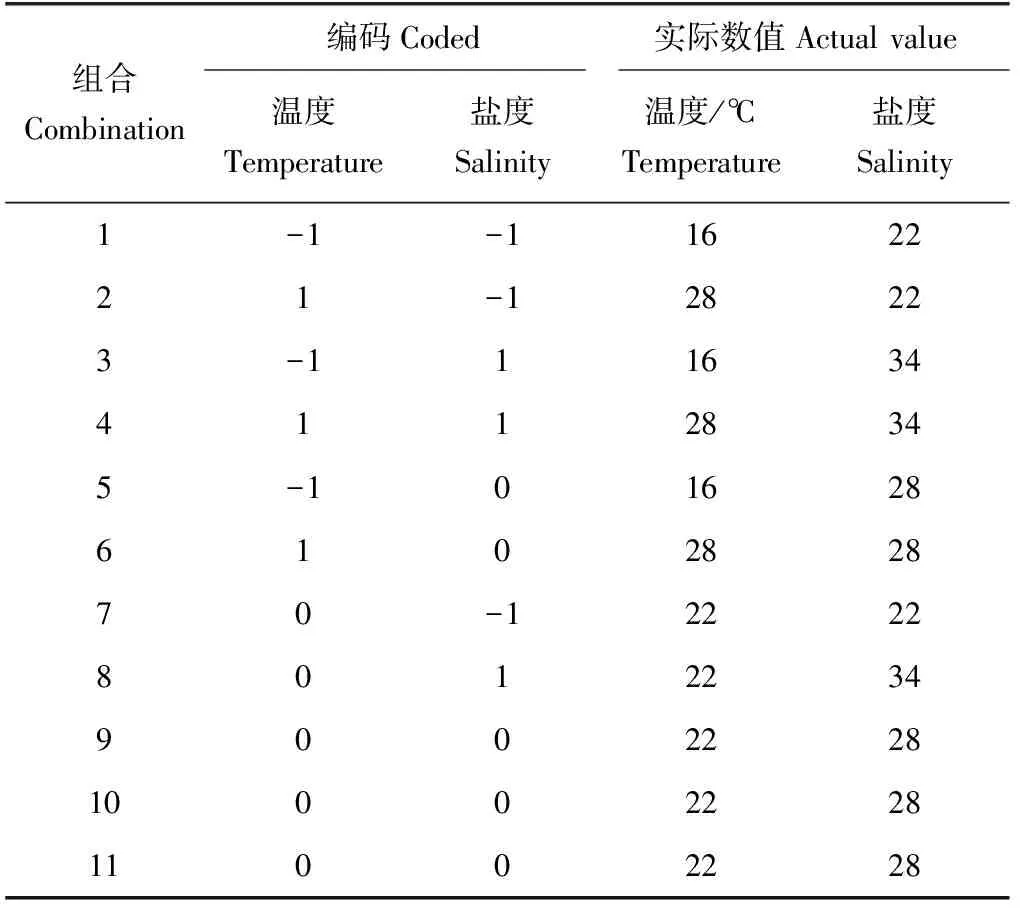
表1 扇贝“渤海红”温度与盐度组合的试验设计Tab.1 Experimental design of combination of temperature and salinity of scallop “Bohai Red”
1.2.2 试验方法与管理
1.2.2.1 试验装置
试验装置由1000 mL烧杯、100 W可控温电子加热棒(±0.5 ℃)、电子温度计(±0.1 ℃)、泡沫箱(30 L)、冰袋、散气石、气管、气泵等组成。
1.2.2.2 试验组别设置
试验分11个组,每组设置3个平行,试验容器为1000 mL烧杯,每个烧杯放入水温20 ℃的海水600~700 mL,置于装有淡水的30 L泡沫箱中进行水浴控温,每个烧杯随机放入经暂养3 d后的健康、活力好且规格一致的稚贝30只。经测量,试验稚贝壳长(8.67±0.022) mm,体质量(0.16±0.002) g。
1.2.2.3 试验水温控制
试验期间,以水浴加热的方式控温,电子温度计用于校正各组温度,室温用空调控制为22 ℃。在淡水里加冰袋和安装可控温加热棒(±0.5 ℃),通过降温和升温达到温度点的动态平衡,并通过循环水泵使水浴温度均匀,泡沫箱口用黑色薄膜覆盖以保温。水温从常温开始,以1 ℃/2 h的速率升温或降温达到各组合预设温度后开始试验。
1.2.2.4 试验盐度控制
提前准备相应盐度的海水,烧杯内的盐度按±3/h的速率添加海水晶溶液或经过曝气的淡水调节至各试验组设置的盐度,达到预设盐度后进行试验。
1.2.2.5 试验饵料投喂
试验期间每日24 h微波充气,各组于每日7:00、15:00和23:00投喂浓缩小球藻,每次投喂饵料10~12 mL,日投喂密度约为1×104个/mL,每次投喂前要检测各平行组饵料密度是否一致,先观察水色再通过血球计数板检测水中的饵料密度,从而调整投喂量,既保证稚贝摄食充足,又不留残饵。
1.2.2.6 试验日常管理
试验期间24 h监控温度,每日各组等温等盐换水50%,及时清理污物与死亡个体,记录死亡时间。判断扇贝死亡的标准为贝壳在反复触碰后没有开闭壳反应。试验共进行14 d,试验结束时统计各组存活率,并测量记录各组稚贝壳长和体质量的生长数据。
1.2.3 测定指标
RS=(n1/n0)×100%
RLG=(L1-L0)/t×1000
RBWG=(m1-m0)/t×1000
式中,RS为稚贝存活率(%),n1和n0分别为试验结束和开始时稚贝的存活数,t为试验时间(d);RLG为壳长增长率(μm/d),L1和L0分别为试验结束和开始时稚贝的平均壳长(cm);RBWG为体质量增长率(mg/d),m1和m0分别为试验结束和开始时稚贝的平均体质量(g)。计算L1和m1时,死亡个体的壳长和体质量按初始数值列入计算,其增长率为0。
1.2.4 数据处理
试验数据采用平均值±标准差表示,通过最小二乘法对温度与盐度的两因子与响应值之间的定量关系进行二次回归拟合,建立回归模型:
y=b0+b1T+b2S+b3TS+b4T2+b5S2
式中,T为温度,S为盐度,y为测定的响应值,b0为回归常数,b1、b2分别为温度和盐度的一次效应,b3为温度和盐度的互作效应,b4、b5为温度和盐度的二次效应。
使用Excel 2016和Design Expert 8.0软件对数据进行处理与作图。显著水平设定为0.05,极显著水平设定为0.01。
2 结果与分析
2.1 不同温度、盐度组合稚贝存活率和壳长及体质量增长率
当温度一定时,随着盐度的增长,存活率、壳长增长率和体质量增长率呈先升后降的趋势(表2);当盐度一定时,随着温度的升高,存活率、壳长增长率和体质量增长率也呈先升后降的趋势,存活率、壳长增长率和体质量增长率最大分别可达97%、200.80 μm/d和21.83 mg/d。从存活率角度看,组合9、10和11之间差异不显著(P>0.05),组合1与组合5之间差异显著(P<0.05);壳长增长率与体质量增长率的规律一致,组合9、10和11之间差异不显著(P>0.05),组合7与8之间、组合4与5之间和组合1与3之间差异不显著(P>0.05)。
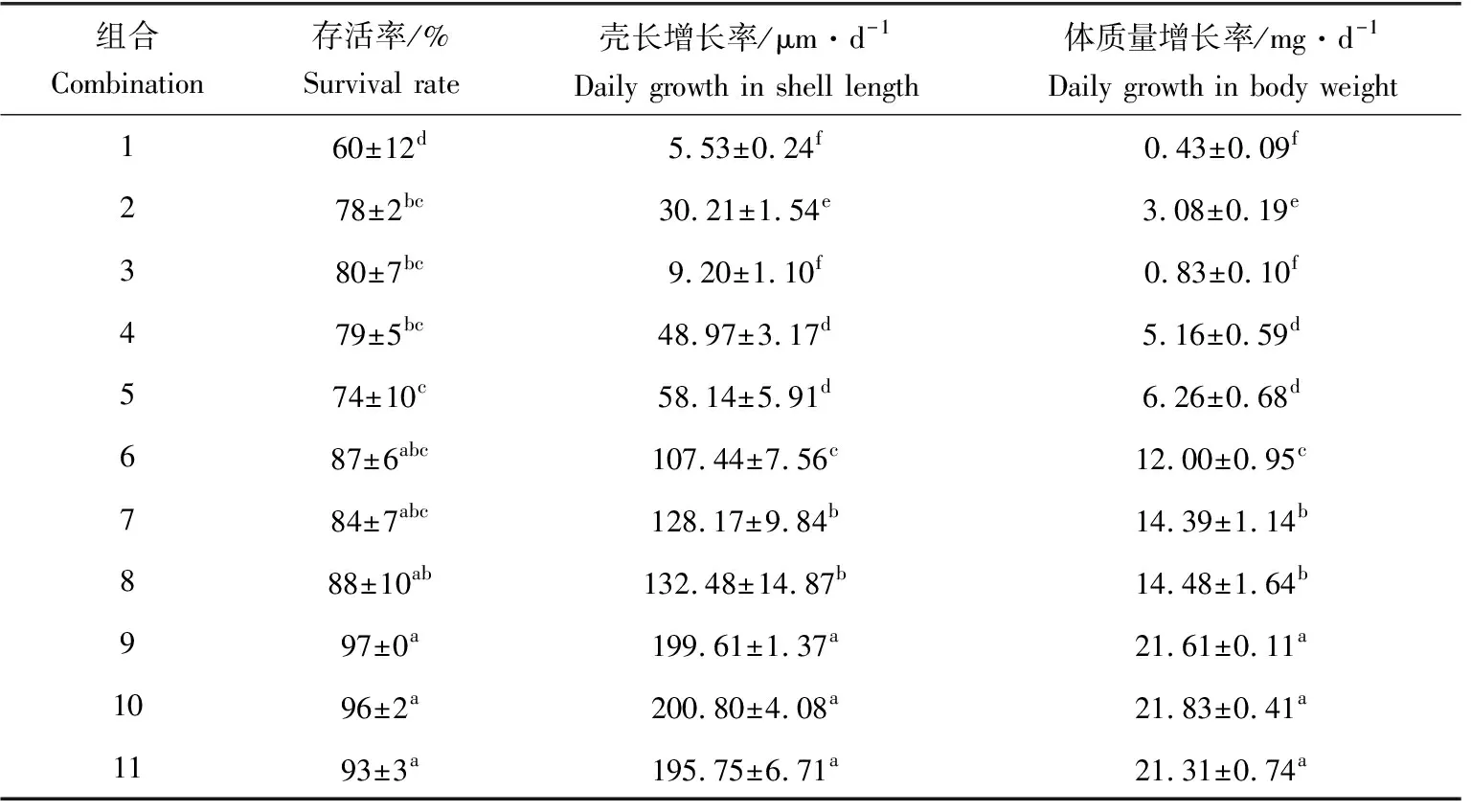
表2 不同温度、盐度组合下稚贝存活率和壳长及体质量增长率Tab.2 Survival rate and daily growth in shell length and body weight of juveniles under different temperature and salinity combinations
2.2 温度与盐度对稚贝存活率的多元回归拟合
对稚贝存活率进行多元回归拟合,得到存活率(y)与温度(T)和盐度(S)的二次多项回归模型y=-3.69898+0.20674T+0.15591S-1.31944×10-3TS-3.66959×10-3T2-2.14181×10-3S2。模型的方差分析结果(表3)表明,所建立的回归模型极显著(P<0.01),而失拟项不显著(P>0.05),表明模型的拟合效果较好。模型的多元相关系数r2=0.9696,校正系数r2=0.9391,表明此模型能解释96.96%的响应值变化,仅有总变异的3.04%不能用此模型解释,模型的拟合效果较好而且预测系数r2=0.7622在校正系数合理范围内,表明可用来预测响应值。
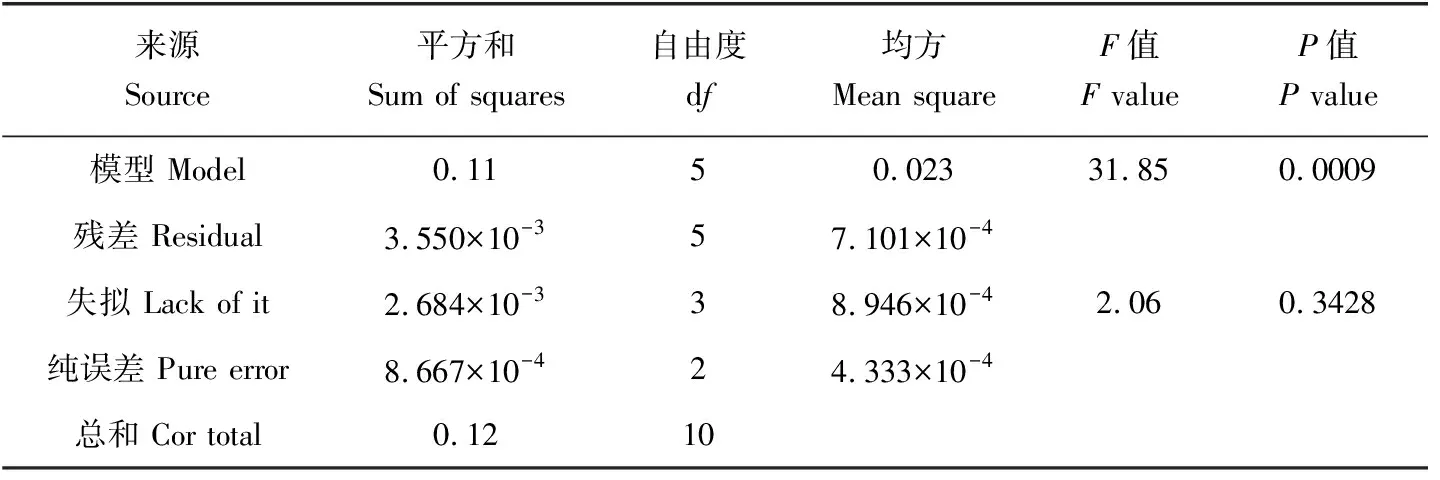
表3 存活率回归模型的方差分析Tab.3 Variance analysis of regression models of survival rate
回归模型系数的显著性检验结果(表4)表明,温度的一次效应、温度的二次效应和盐度的二次效应对稚贝的存活达到极显著水平(P<0.01);盐度的一次效应和温度与盐度的互作效应对稚贝存活率的影响达到显著水平(P<0.05)。
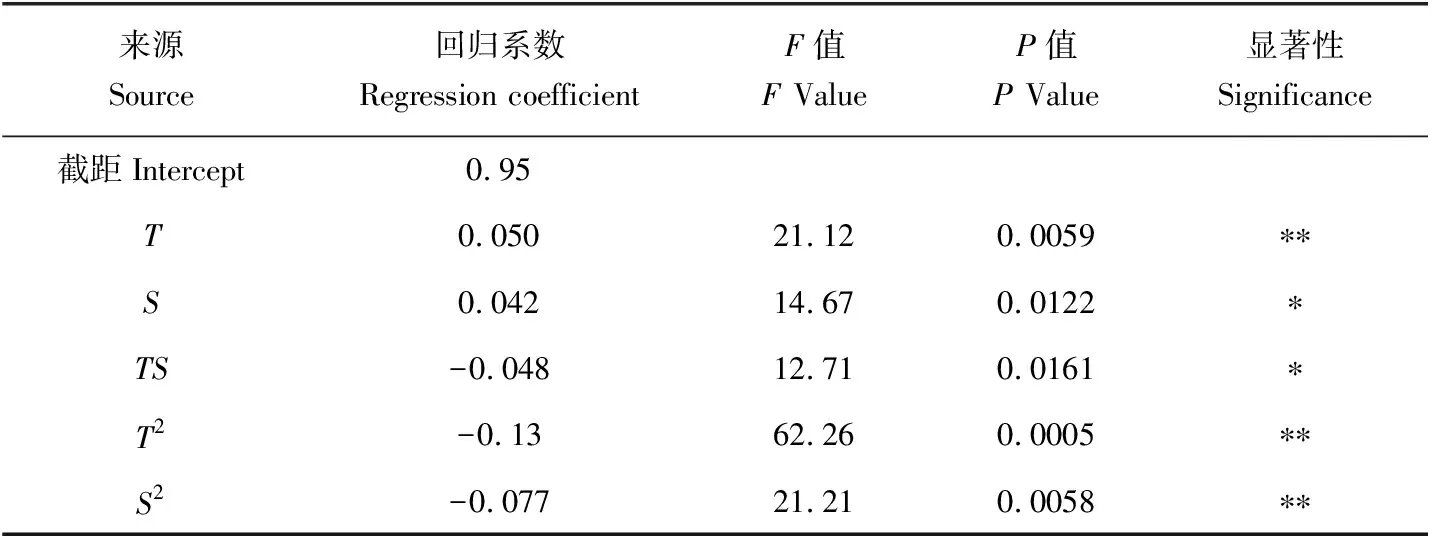
表4 存活率模型回归系数的显著性检验Tab.4 The significance test for the regression coefficient of survival rate model
根据软件得到稚贝存活率的响应曲面和等高线图(图1)。温度对存活率的影响曲线较陡,影响显著(P<0.01),而盐度对存活率的影响曲线较为平缓(P<0.05)(图1a);等高线图为椭圆形,表明温度和盐度的交互作用显著(P<0.05)(图1b)。随着温度升高和盐度增大,存活率呈先升后降的趋势。对存活率的模型进行优化,结果显示,温度22.89 ℃、盐度29.35是存活的最优温度、盐度组合,此时存活率最大达95.52%,可靠性为0.960。
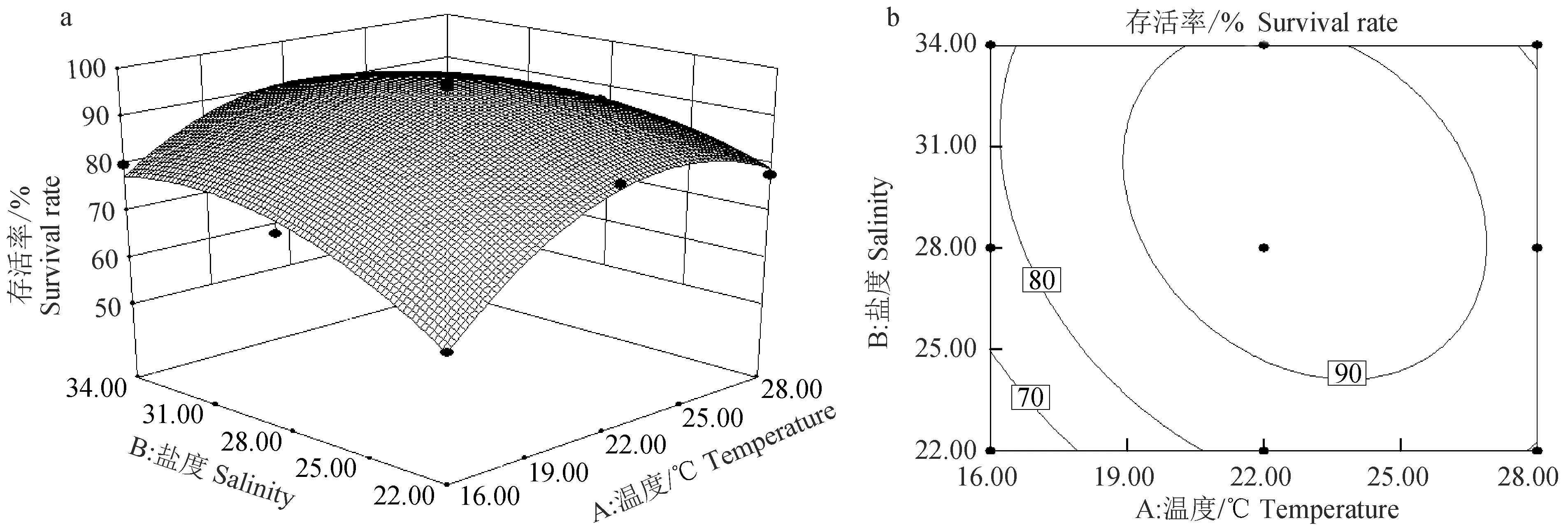
图1 温度、盐度及其互作效应对扇贝“渤海红”稚贝存活率影响的响应曲面(a)和等高线图(b)Fig.1 Response surface plot (a) and its contour plot (b) of the effects of temperature, salinity and their mutual interactions on the survival rate of juvenile scallop “Bohai Red”
2.3 温度与盐度对稚贝壳长增长率的多元回归拟合
对稚贝壳长增长率进行多元回归拟合,得到壳长增长率(y)与温度(T)和盐度(S)的二次多项回归模型y=-2708.68617+136.07523T+97.39362S+0.10479TS-3.08749T2-1.76708S2。模型的方差分析结果(表5)表明,所建立的回归模型极显著(P<0.01),而失拟项不显著(P>0.05),表明模型的拟合效果较好。模型的多元相关系数r2=0.9969,校正系数r2=0.9938,表明此模型能解释99.69%的响应值变化,仅有总变异的0.31%不能用此模型解释,模型的拟合效果很好,且预测系数r2=0.9726接近校正系数,可用来预测响应值。
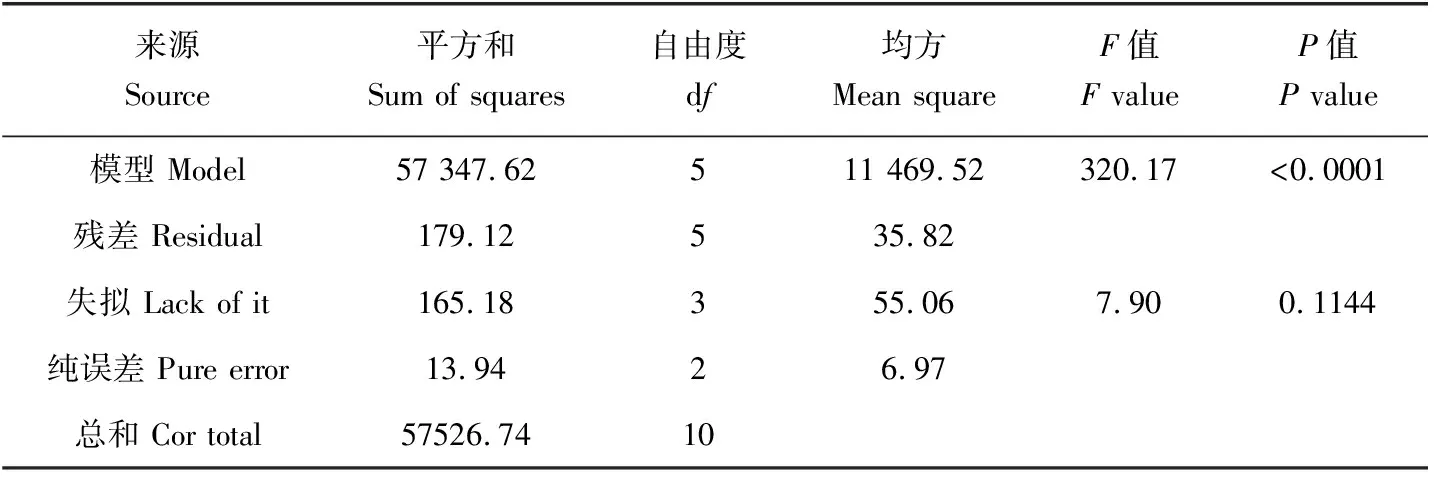
表5 壳长增长率回归模型的方差分析Tab.5 Variance analysis of regression model of shell length growth model
回归模型系数的显著性检验结果(表6)表明,温度的一次效应、二次效应和盐度的二次效应对稚贝的壳长增长率达到极显著水平(P<0.01);而盐度的一次效应、温度和盐度的互作效应对稚贝的壳长增长率不显著(P>0.05)。
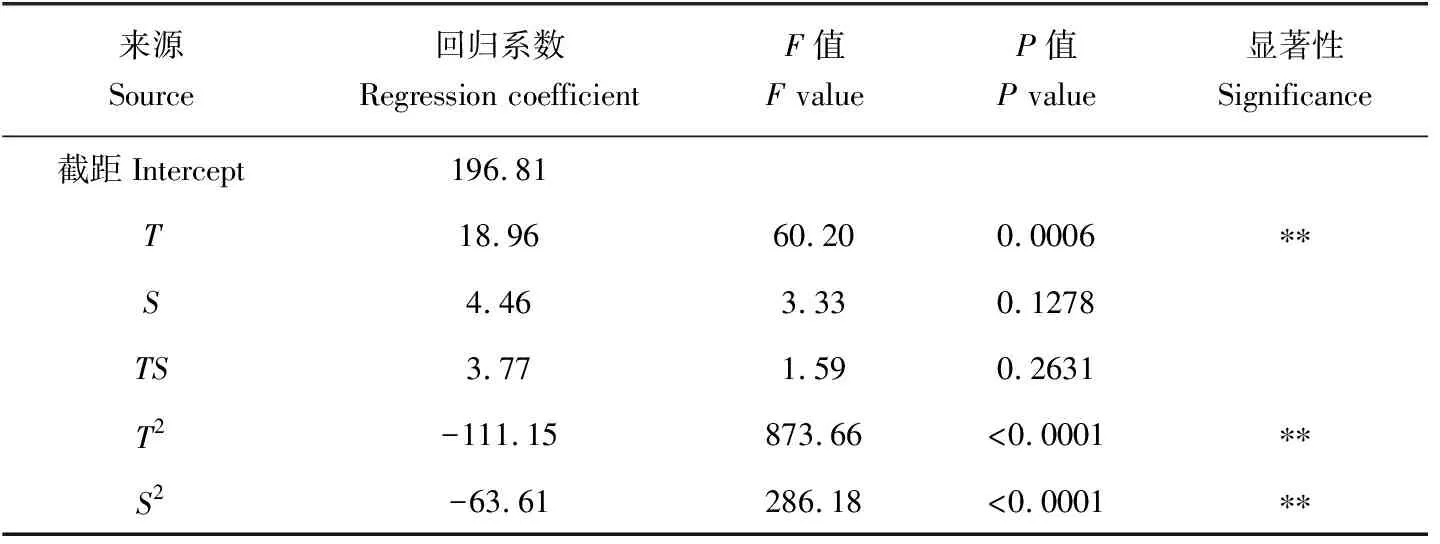
表6 壳长增长率回归模型系数的显著性检验Tab.6 The significance test for the regression coefficient of the daily growth model of shell length
根据软件得到稚贝壳长增长率的响应曲面和等高线图(图2)。温度对壳长增长率的影响曲线较陡,影响极显著(P<0.01),盐度对稚贝壳长增长率的影响曲线平缓,影响不显著(P>0.05)(图2a),温度效应大于盐度效应;等高线图为圆形,表明温度和盐度的交互作用不显著(P>0.05)(图2b)。随着温度升高和盐度的增大,壳长增长率呈先升后降的趋势。对壳长增长率的模型进行优化,结果显示,温度22.52 ℃、盐度28.23是壳长增长率的最优温度、盐度组合,此时壳长增长率最大达到197.71 μm/d,可靠性为0.984。
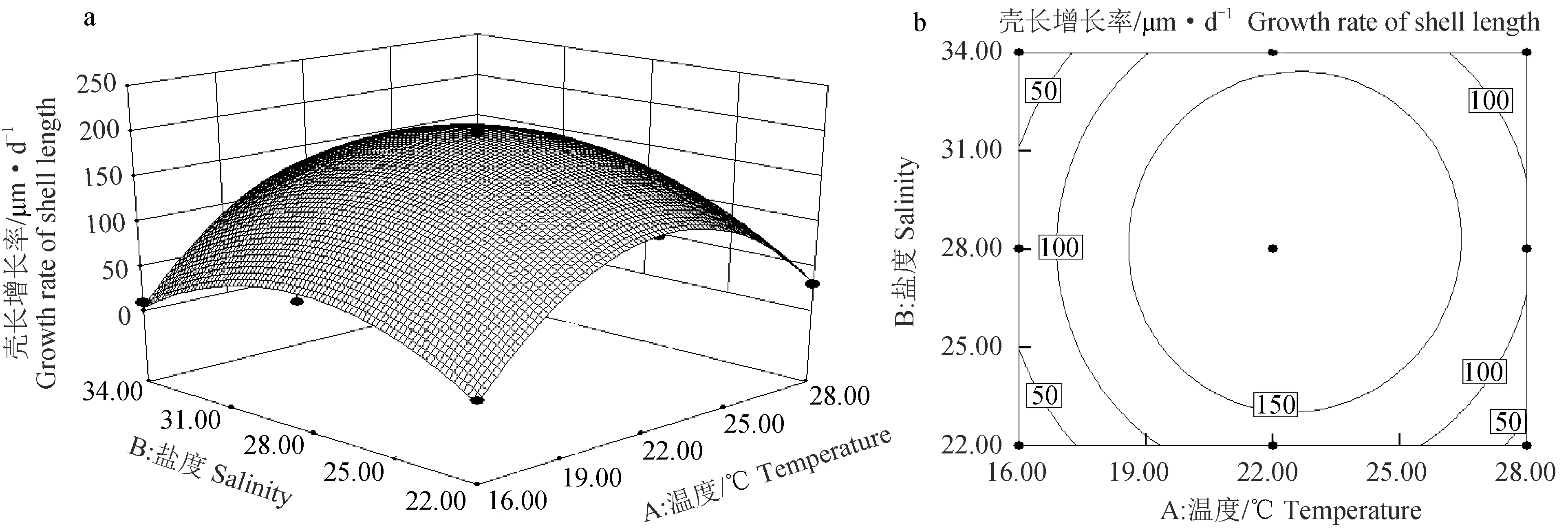
图2 温度、盐度及其互作效应对扇贝“渤海红”稚贝壳长增长率影响的响应曲面(a)和等高线图(b)Fig.2 Response surface plot (a) and its contour plot (b) of the effects of temperature, salinity and their mutual interactions on the daily growth of shell length of juvenile scallop “Bohai Red”
2.4 温度与盐度对稚贝体质量增长率的多元回归拟合
对稚贝体质量增长率进行多元回归拟合,得到稚贝体质量增长率(y)与温度(T)和盐度(S)的模型y=-296.89637+14.99439T+10.61232S+0.011667TS-0.34018T2-0.19281S2。模型的方差分析结果(表7)表明,所建立的回归模型极显著(P<0.01),而失拟项不显著(P>0.05),表明模型的拟合效果较好。模型的多元相关系数r2=0.9966,校正系数r2=0.9932,表明此模型能解释99.66%的响应值变化,仅有总变异的0.34%不能用此模型解释,模型的拟合效果很好且预测系数r2=0.9662接近校正系数,表明可用来预测响应值。
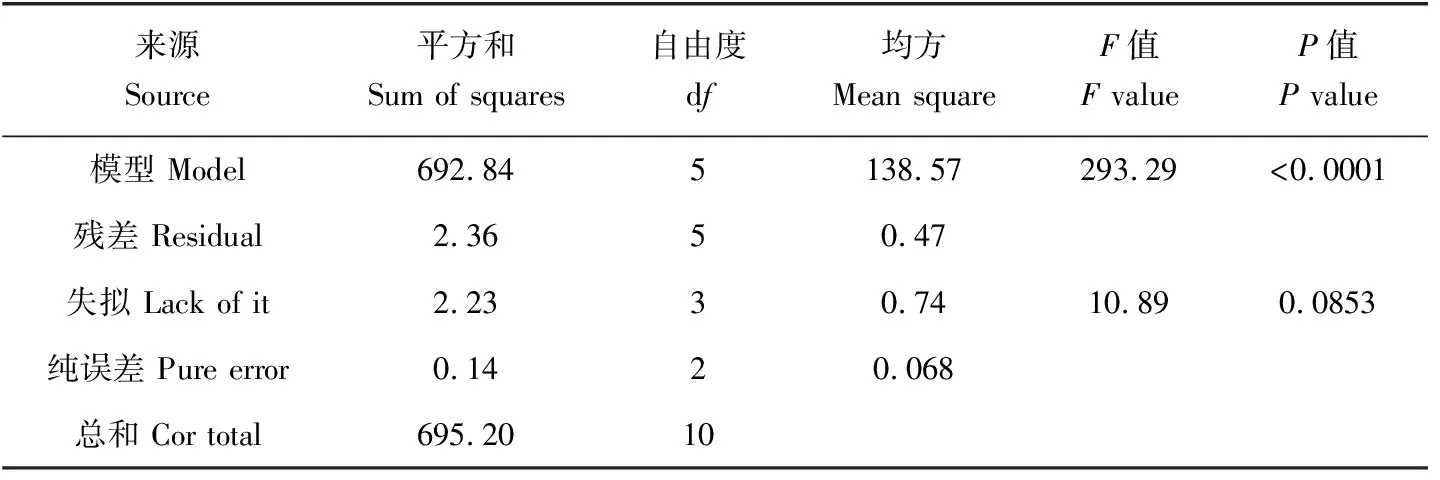
表7 体质量增长率回归模型的方差分析Tab.7 Variance analysis of regression model of the daily growth of body weight
回归模型系数的显著性检验结果(表8)显示,温度的一次效应、二次效应和盐度的二次效应对稚贝的体质量增长率达到极显著水平(P<0.01);而盐度的一次效应、温度和盐度的互作效应对稚贝的体质量增长率不显著(P>0.05)。
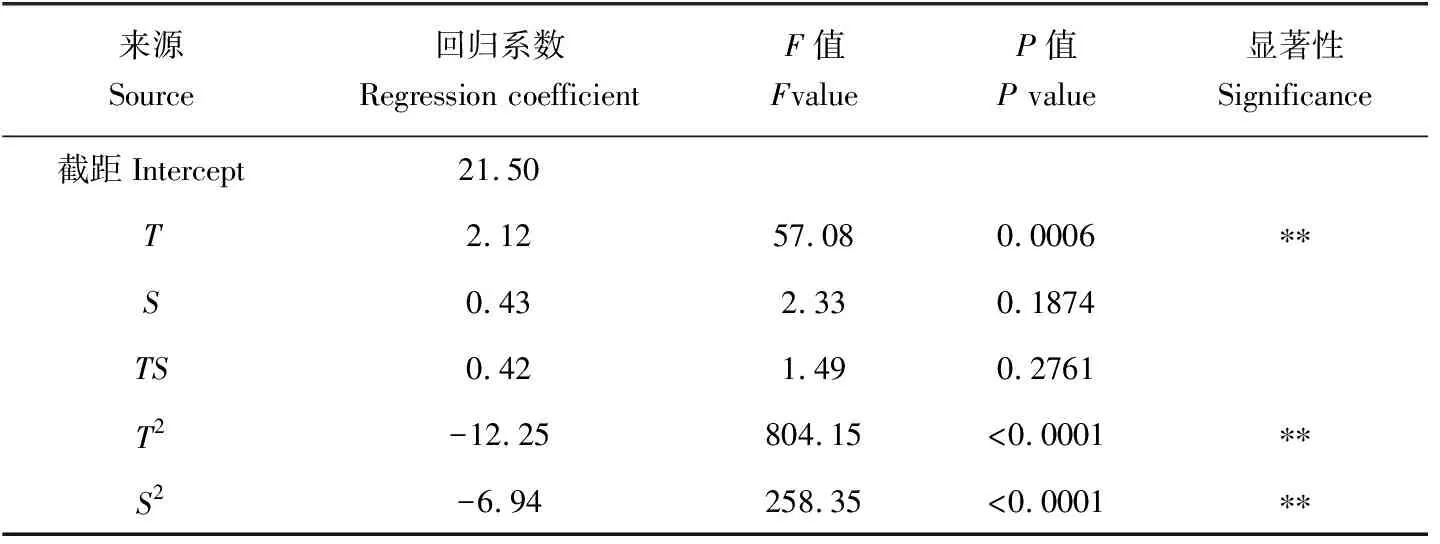
表8 体质量增长率回归模型系数的显著性检验Tab.8 The significance test for the regression coefficient of the daily growth of shell body weight
根据软件得到稚贝体质量增长率的响应曲面和等高线图(图3)。温度对体质量增长率的影响曲线较陡,影响显著(P<0.01),盐度对稚贝体质量增长率的影响曲线平缓,影响不显著(P>0.05)(图3a),温度效应大于盐度效应;等高线图为圆形,表明温度和盐度的交互作用不显著(P>0.05)(图3b)。随着温度升高和盐度的增大,体质量增长率呈先升后降的趋势。对体质量增长率的模型进行优化,结果显示,温度22.52 ℃、盐度28.20是体质量增长率的最优温度、盐度组合,此时体质量增长率最大达21.60 mg/d,可靠性为0.989。
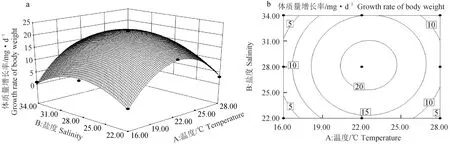
图3 温度、盐度及其互作效应对扇贝“渤海红”稚贝体质量增长率影响的响应曲面(a)和等高线图(b)Fig.3 Response surface plot (a) and its contour plot (b) of the effects of temperature,salinity and their mutual interactions on the daily growth of shell body weight of juvenile scallop “Bohai Red”
2.5 最优结果的筛选过程
使用相应曲面分析法筛选最优值,其过程为选择左侧优化功能下的数值选项,进一步确定温度与盐度的取值范围,确定响应值(存活率、壳长增长率和体质量增长率)的目标即最大值,响应值的低值取默认值,高值取一个较大且无法达到的值,最后在解决项选择最优方案即可。经软件优化得到温度、盐度的共同影响下存活率、壳长增长率和体质量增长率的最优结果,即当温度22.65 ℃,盐度28.51时,存活率为95.33%,壳长增长率为197.50 μm/d,体质量增长率为21.58 mg/d,其可靠性为0.975。
3 讨 论
3.1 温度对稚贝存活与生长的影响
温度是对海水贝类生长及存活有着重要影响的环境因子之一[20]。温度适宜时,升温可能促进海洋生物幼虫的生长[21-22],稚贝的生理活动变强,贝类生长加快;温度不适宜时,高温会影响贝类的血细胞和酶活性[23],虽短时间内可提高机体免疫能力,但长时间会对细胞造成伤害[13],低温会抑制贝类体内的新陈代谢相关酶活性[24-25],贝类体内外的平衡被打破,能量转移以维持正常存活,生长部分的能量补给不足,因此出现生长缓慢、不生长甚至出现死亡的现象。由于扇贝是滤食性动物,其摄食活动与其体内的纤毛有极大的联系,当温度适宜时,其纤毛活动加强,摄食活动亦随之增强[26],在水温13~33 ℃时, 香港巨牡蛎(Crassostreahongkongensis)[27]的滤水率、摄食率和吸收率随着水温的升高先增后降,菲律宾蛤仔(Ruditapesphilippinarum)[28]的研究也具有相似变化规律。贝类摄取食物后,其有机碳源主要通过呼吸代谢消耗,除去排泄代谢以及粪便形式排出的部分能量,余下的用于其本身的生长[29-30]。泥蚶(Tegillarcagranosa)浮游幼虫及稚贝的适宜生长温度分别为25~33 ℃和15~35 ℃,而泥蚶幼虫和稚贝较强的高温耐受性与其生态环境相适应[12],墨西哥湾扇贝幼虫和稚贝的适宜生长温度均为25~30 ℃[31]。本研究结果表明,温度的一次效应与二次效应对稚贝的存活均产生影响,适宜存活的温度为22~28 ℃,最适生长温度为22 ℃。这可能与其是北方养殖品种有关。
在试验温度范围内,随着温度的升高其稚贝的存活率、壳长增长率和体质量增长率均表现为先升后降的趋势。琴文蛤(Meretrixlyrate)幼贝的生存温度为12.2~35.6 ℃,最适生长温度为27~30 ℃[32],栉江珧(Atrinapectinate)的早期幼虫生长发育适宜温度为27~30 ℃[33],岩扇贝(Crassadomagigantea)的最适生长温度为15 ℃,文蛤(M.meretrix)稚贝的适宜生长温度为24~27 ℃[34],随试验温度的升高,以上贝类生长指标均呈现先升后降的趋势。试验中生长速度和存活率存在明显正相关关系,即生长速度快的组别存活率也较高。海湾扇贝南方亚种(A.irradiansconcentricus)选育中,优良选育家系的生长速度与存活率均高于对照组,而其他家系低于对照组[35];但也有研究表明,低温时幼虫生长缓慢,存活率较高[36]。当温度在一定范围内变化时,贝类的呼吸状况也会发生改变,其耗氧率与排氨率会随着温度的升高而增加[24,37],亦会随温度的下降而降低[38],有可能是温度下降促使贝类的呼吸代谢底物发生变化。
3.2 盐度对稚贝存活与生长的影响
盐度是影响海洋生物生长与存活的另一重要自然环境因子,雨季和旱季会影响水体盐度,从而让贝类置于不稳定的盐度环境下,贝类属于变渗透压的软体动物,会根据水体的盐度变化,调节细胞膜上的Na+/K+-ATP酶来维持体内外平衡,同时为鳃和肾脏上的离子调控提供驱动力[39]。在盐度改变时,就会调节自身的渗透压保持体内外平衡,而且一定程度上会影响贝类的摄食率。林丽华等[40]研究了盐度对香港巨牡蛎摄食和代谢的影响,其结果也表明,滤水率和摄食率会随着盐度的增加呈先升后降的趋势,与陈丽梅等[29]的研究结果一致。低盐度环境下贝类为适应环境会关闭贝壳,降低贝类与外界水流的交换频率,摄食率下降;高盐度环境超过贝类本身可调控范围,正常生理平衡被打破,也会影响贝类的摄食率[41]。若贝类持续在不适盐度环境下生存,长期关闭贝壳以应对环境的改变,会影响贝类的存活与生长[42],甚至会因为摄食不足而死亡,降低存活率。
李卫东等[43]研究表明,盐度的二次效应对华贵类栉孔扇贝(Mimachlamysnobilis)幼虫的存活影响显著。本研究结果也表明,稚贝的存活与盐度有线性关系,适宜的存活盐度为22~34,但稚贝的生长与盐度无显著相关,极有可能是试验所设盐度梯度在稚贝所承受范围之内。在试验盐度范围内,随着盐度的升高其稚贝的存活率、壳长增长率和体质量增长率均表现为先升后降的趋势,这与壳黑长牡蛎(Crassostreagigas)[44]、宽壳全海笋(Barneadilatata)稚贝[45]、大珠母贝(Pinctadamaxima)[46]和合浦珠母贝(P.fucata)[46]的研究结果一致。在适宜盐度范围内,稚贝存活率较高而且生长较快,能量大部分用于生长,如若超出适宜盐度,稚贝就会分配其余能量调控渗透压[47-48],因此过高或过低的盐度均会对稚贝的存活与生长造成一定影响。
3.3 温度与盐度互作效应对稚贝存活与生长的影响
研究表明,当生物试验受到多因素影响时,互作效应的研究更能揭示客观规律[49]。Kong等[11]研究表明,皱纹盘鲍(Haliotisdiscushannai)稚贝的存活和生长不仅受温度和盐度的影响,还受二者之间的互作效应影响;Xue等[50]研究表明,方斑东风螺(Babyloniaareolata)稚贝的生长受温度、盐度及二者互作效应影响,但稚贝的存活率只受温度影响;孔宁等[49]的温度与盐度双因素方差分析表明,其各自以及互作效应均对皱纹盘鲍生长和存活造成影响,并在存活率方面,盐度效应强于温度效应,而特定生长率与之相反;刘志刚等[12]研究结果指出,温度与盐度对华贵类栉孔扇贝幼贝存活及生长有互作效应。而梁玉波等[51]研究表明,随着幼虫的发育,温度与盐度对栉孔扇贝(Chlamysfarreri)的存活有互作效应,对生长无互作效应;桑士田等[52]对菲律宾蛤仔的研究表明,温度、盐度及pH对蛤仔稚贝生长无交互作用;李卫东等[43]研究表明,温度和盐度对华贵类栉孔扇贝幼虫存活在一定范围内亦无互作效应。有一些学者认为,温度和盐度如果存在互作效应,那其中一个因子要接近极限[53-54],而当其中一个因子在安全范围内,单因子试验能够反映温度或者盐度对海洋生物的影响[55],如若两因子均在适宜范围,其互作效应就不明显。笔者对扇贝“渤海红”稚贝的存活与生长进行了温度与盐度的研究,发现稚贝存活受到温度和盐度的互作影响,而且温度效应高于盐度效应,但稚贝的生长不受温度与盐度互作的影响。在养殖扇贝“渤海红”时要注意温度与盐度对稚贝的影响,也要注意其温度与盐度的互作效应。
4 结 论
经软件优化得到温度、盐度的共同影响下存活率、壳长增长率和体质量增长率的最优结果,即当温度为22.65 ℃、盐度为28.51时,存活率为95.33%,壳长增长率为197.50 μm/d,体质量增长率为21.58 mg/d,其可靠性为0.975。本试验确定扇贝“渤海红”稚贝的存活与生长的最适温盐组合,可为亲本培育新品种提供一个适宜暂养温度,为人工养殖提供试验依据,从而为缩短养殖周期达到利益最大化提供可行性,还利于对其与墨西哥湾扇贝杂交后代的温盐适应性进行评价。
