望春玉兰WRKY基因家族生物信息学分析
2022-01-21戴晓港刘景胜李淑娴
戴晓港, 刘景胜, 李淑娴
(南京林业大学林学院/南方杨树工程技术研究中心/江苏省杨树种质创新与品种改良重点实验室, 江苏 南京 210037)
植物在整个生命活动过程中会不断受到各种生物胁迫(如食草动物和害虫的啃食、病原体侵染)和非生物胁迫(如干旱、低温、高温、土壤高盐碱等)的影响,然而植物不能自由移动来躲避这些不利环境,因此在漫长的进化过程中,植物逐渐从形态、生理、细胞以及分子水平发生进化以应对这些不利环境条件。当植物受到胁迫时会产生相应的应答反应,相关基因、蛋白质和代谢产物构成复杂调控网络以应答相应的胁迫。转录因子作为反式作用因子可以与靶标基因启动子区域相应的顺式作用元件特异结合,从而调控相关基因的转录,在植物应答生物胁迫和非生物胁迫中具有举足轻重的作用[1]。WRKY转录因子是植物中的超级转录因子家族之一,其典型特征是在氨基酸N-端有保守的WRKYGQK结构域,在C-端含有C2H2或C2HC型锌指结构[2]。根据上述保守结构域数量和锌指结构类型可将WRKY转录因子分为3组:第I组包含2个WRKY结构域和C2H2型锌指结构;第II组含有1个WRKY结构域和C2H2型锌指结构;而第Ⅲ类是由1个WRKY保守结构域和C2HC型锌指结构组成[2]。大量研究表明,WRKY在调控植物生长发育、生物胁迫和非生物胁迫[3-4]、次生代谢等多种生物过程中起着重要的作用[5-6]。如木本鸡脚棉中GaWRKY1转录因子通过调控卡丁烯合酶-A的活性,从而参与棉酚等倍半萜烯类化合物的合成[5];从黄花蒿分泌型腺毛中分离的AaWRKY1通过上调倍半萜环化酶基因ADS的表达,从而提高抗疟青蒿素的生物合成[6]。随着基因组测序的发展,药用植物越来越引起大家的关注,这也为鉴定更多的参与调控次生代谢的WRKY转录因子提供可能。
望春玉兰(Yulaniabiondii)是木兰科(Magnoliaceae)玉兰属多年生落叶乔木,是中国特有的珍稀濒危树种,原产湖北、河南、陕西、甘肃等地海拔600—1 200 m的阔叶林中[7]。望春玉兰先花后叶,种子鲜红,树形优美,被广泛应用于园林绿化[7],其干燥的花蕾是中药材“辛夷”的正品,具有散风寒、通肺窍、降压、杀菌等作用,是一种用途广泛的药用植物[8]。望春玉兰具有一定的耐寒性,喜微酸性土壤,但碱性土壤对生长的影响较大。望春玉兰全基因组测序的完成为WRKY转录因子家族的鉴定和分析提供了基础,本研究对望春玉兰WRKY基因家族成员进行鉴定,同时对蛋白理化性质、亚细胞定位、系统进化、保守结构域、顺式作用元件、以及在不同组织中的表达模式进行了初步研究,以期为深入研究该基因家族在望春玉兰中应答非生物胁迫以及次生代谢的调控奠定基础。
1 材料与方法
1.1 WRKY转录因子查找及理化性质分析
从国家基因库生命大数据平台(https://db.cngb.org/codeplot/)中下载望春玉兰基因组和转录组原始数据(CNP0000884)[9],利用hmmsearch v3.3.2将WRKY隐马尔可夫模型(Pfam: PF00847)和下载的望春玉兰蛋白序列进行比对,保留E-value≤1e-5的蛋白序列,再采用SMART(http://smart.embl-heidelberg.de/)进行保守结构域鉴定[10],去除不含WRKY保守结构域的蛋白序列。采用Expasy[11](https://web.expasy.org/protparam/)在线预测蛋白分子量、等电点等理化性质,并利用Cell-PLoc 2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)对鉴定的WRKY基因进行亚细胞定位预测。
1.2 WRKY转录因子进化树分析
从拟南芥基因组数据库(https://www.arabidopsis.org/browse/genefamily/WRKY-Som.jsp)下载拟南芥WRKY转录因子序列。根据SMART分析结果提取拟南芥和望春玉兰WRKY转录因子的保守结构域序列,并提交到MEGA-6.0[12]软件中利用ClustalW进行多重比对,然后用邻接法(Neighbor Joining method)构建进化树,bootstrap值设置为1 000,最后采用Figtree(http://tree.bio.ed.ac.uk/software/figtree/)对构建的进化树进行可视化,根据进化树中拟南芥WRKY家族的分类对望春玉兰WRKY家族进行类群的划分。
1.3 WRKY转录因子保守基序和基因结构
利用MEME(https://meme-suite.org/meme/tools/meme)在线预测望春玉兰WRKY蛋白的保守基序(Motif),Motif个数设置为10个,基序长度为6—70,其他参数均为默认值。根据MEME的分析结果,结合基因注释的gff文件,利用TBtools[13]对保守基序、基因结构和进化树进行整合,绘制望春玉兰WRKY转录因子保守基序和基因结构图。
1.4 WRKY转录因子顺式作用元件分析
利用perl脚本根据基因组注释的gff文件,从基因组中提取目的基因上游2 000 bp序列,用PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)对启动子区域进行顺式作用元件分析,最后利用TBtools[13]结合进化树并绘制顺式作用元件分布图。
1.5 WRKY转录因子染色体定位及串联重复基因分析
根据基因组注释的gff文件,使用TBtools[13]绘制望春玉兰WRKY转录因子在染色体上的分布图。串联重复基因的鉴定参照Lehti-Shiu对毛果杨基因组串联重复基因的鉴定标准:2个基因之间不超过350 kb且中间插入的基因不超过10个,进化树上它们还需要同属于1个亚家族[14]。
1.6 WRKY转录因子表达分析
将从国家基因库生命大数据平台中下载的望春玉兰叶片(CNX0149886)和花(CNX0149883)的转录组,以及实验室测序的根系转录组,使用Trimmomatic[15]过滤低质量序列,然后利用STAR[16]默认参数将上述过滤后的序列分别比对到望春玉兰基因组,统计根、叶和花分别比对到每个WRKY基因的序列数量,采用RPKM(reads per kilobase of transcripts per million mapped reads)对每个基因表达量进行均一化。
2 结果与分析
2.1 望春玉兰WRKY基因家族成员鉴定及理化性质分析
本研究共鉴定出望春玉兰WRKY基因家族有56个成员,各成员蛋白长度、理化性质、理论等电点和亚细胞定位等信息如表1。从表1中结果可以看出,蛋白长度在106—1 675个氨基酸之间,不同基因氨基酸数量存在较大差异。通过Expasy在线分析了望春玉兰WRKY转录因子蛋白理化性质,该家族基因蛋白分子量在12 215.15—191 326.88 Da之间,理论等电点在4.9—10.06之间。亚细胞定位结果显示,55个基因家族成员定位在细胞核,其中MBI36866_MAGBIO成员同时定位在细胞外间隙和细胞核,而MBI24213_MAGBIO还定位在细胞质。

表1 望春玉兰WRKY家族基因基本信息及蛋白理化性质
2.2 WRKY转录因子进化分析
以望春玉兰和拟南芥WRKY基因蛋白保守域序列构建进化树,参照拟南芥WRKY基因家族的分类,根据进化树和保守域序列结构特征,将望春玉兰WRKY转录因子划分为Group Ⅰ,Ⅱ和Ⅲ共3个组。Group Ⅰ具有2个WRKY保守结构域,锌指结构为C2H2型,共有8个基因;Group Ⅱ只含有1个WRKY保守结构域,锌指结构也是C2H2型,但根据进化树分析结果又可将Group II进一步分为Ⅱa,Ⅱb,Ⅱc,Ⅱd,Ⅱe共5个亚组,其成员数量分别为9,7,10,7,4个;Group Ⅲ虽然也只含有1个WRKY保守结构域,但其锌指结构(C2HC)和其他2组不同,该组中共有7个基因。还有3个基因(MBI36007_MAGBIO、MBI46007_MAGBIO和MBI29696_MAGBIO)虽然含有WRKY保守结构域,但序列与其他基因分化较大而未被分类到现有的I,Ⅱ和Ⅲ组中(见图1)。
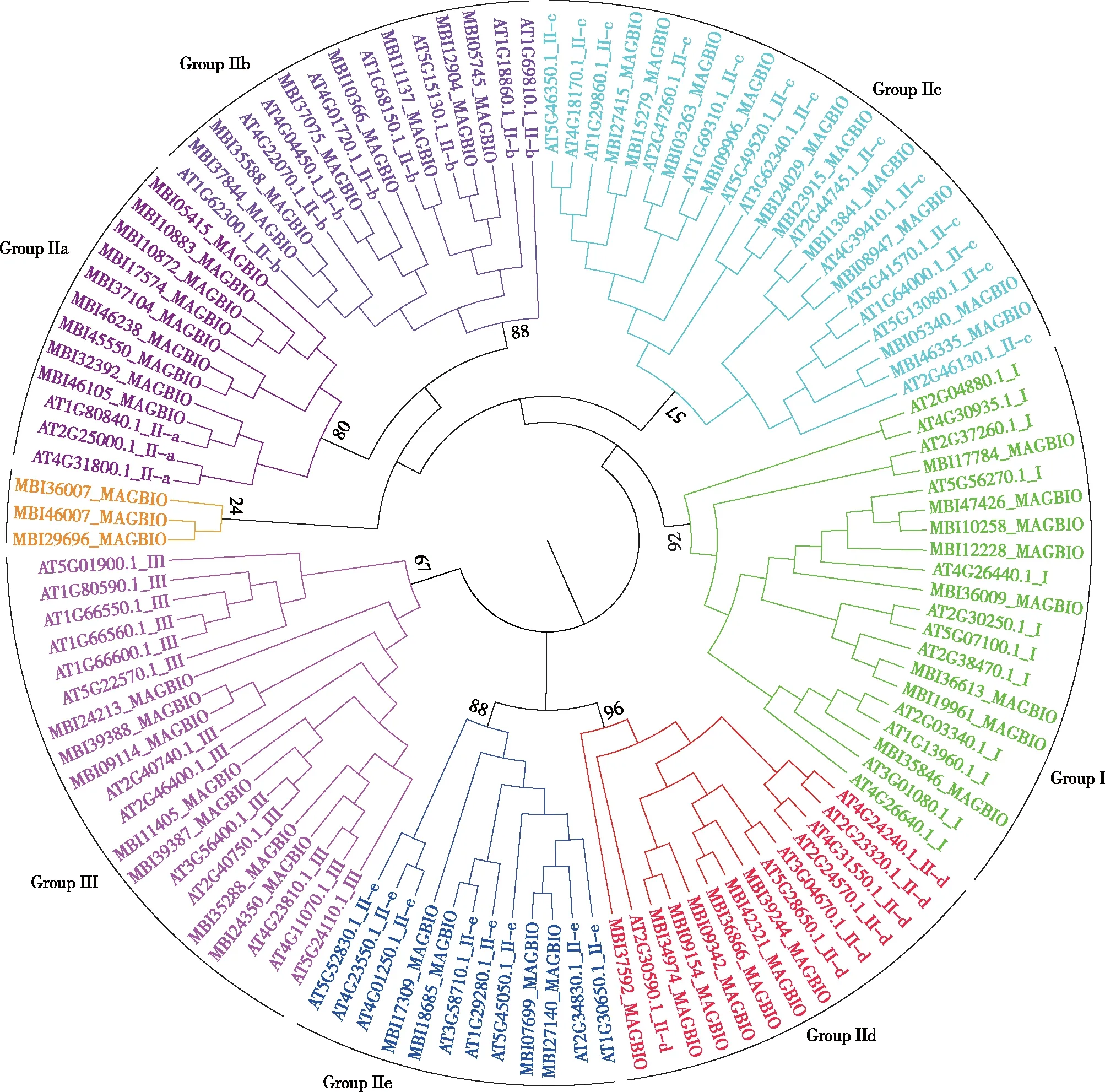
图1 望春玉兰和拟南芥WRKY基因家族进化树
2.3 WRKY转录因子蛋白基序分析
利用MEME在线分析望春玉兰WRKY基因家族的Motif,利用TBTools对Motif分析结果进行可视化(见图2)。望春玉兰WRKY蛋白共包含10个Motif,其中Motif 1,Motif 2和Motif 3是WRKY的保守结构域,Motif 1含有保守序列WRKYGQK,望春玉兰所有WRKY蛋白都含有这个保守结构域;Motif 1尾端2个蛋白和Motif 2构成了WRKY结构域的锌指结构(C2H2),其中48个基因含有Motif 2结构域;Motif 3是WRKY的完整结构域,其中7个基因同时含有Motif 3和Motif 1,这类基因因含有2个保守的WRKY结构域而被划分为Group I。Motif 4—Motif 10虽然在数据库中暂无功能记录,但这些Motif也为望春玉兰WRKY基因分类提供了重要参考,如Motif 8和Motif 10是Group I特有的,而Motif 5,Motif 6和Motif 9是Group IIa和Group IIb中特有。总体而言,望春玉兰WRKY基因家族不同分组中Motif是相似的,这个结果也有利地支持了进化树对基因家族分类的可靠性。
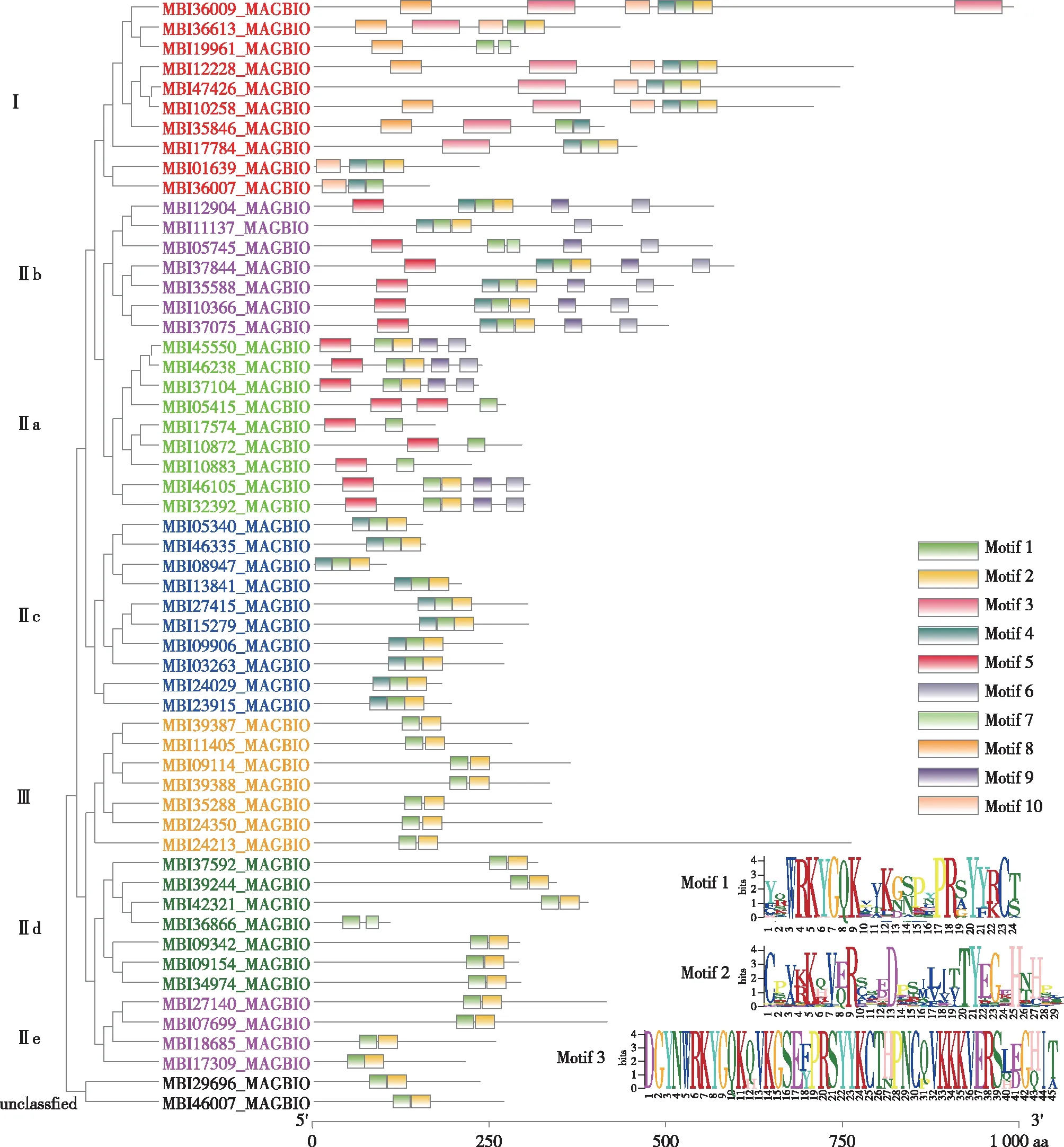
图2 望春玉兰WRKY基因家族蛋白保守结构域
2.4 WRKY转录因子启动子区域顺式作用元件
利用PlantCARE注释望春玉兰56个WRKY基因上游2 000 bp启动子区域序列,得到WRKY基因家族顺式作用元件,除最基本的TATA box,CAAT box和光响应元件之外,还含有参与各种激素、调控生长、参与次生代谢和非生物胁迫等响应元件。在激素类响应元件中,分别有53个和47个基因含有脱落酸反应元件(abscisic acid responsiveness)和茉莉酸甲酯反应元件(MeJA-responsiveness);应答水杨酸、赤霉素和生长素响应元件的基因相对较少,分别有32,26,18个。参与非生物胁迫相关的主要有无氧诱导(the anaerobic induction)、干旱诱导(drought-inducibility)、低温响应(low-temperature responsiveness)和防御胁迫(defense and stress responsiveness)等所必需的调节元件。MBI11137_MAGBIO含有多个干旱诱导响应顺式作用元件,推测该基因可能调控望春玉兰响应干旱胁迫。值得注意的是望春玉兰WRKY基因除了参与激素响应和非生物胁迫的顺式作用元件,其中还有9个基因(MBI08947_MAGBIO,MBI10258_MAGBIO,MBI10883_MAGBIO,MBI12228_MAGBIO,MBI19961_MAGBIO,MBI36613_MAGBIO,MBI36866_MAGBIO,MBI42321_MAGBIO,MBI46335_MAGBIO)含有类黄酮合成调节元件,参与植物的次生代谢。望春玉兰的花蕾和根系入药,表明了WRKY基因可能参与了次生代谢产物的合成调节。
2.5 WRKY基因家族染色体分布
望春玉兰基因组共有19条染色体,而WRKY基因只分布在其中14条染色体上(见图4),Chr08,Chr09,Chr12,Chr13和Chr17均不含有WRKY基因。在含有WRKY基因的染色体中,Chr15,Chr14和Chr04这3条染色体含有WRKY基因的数量约占总数的48.2%,分别有12,9,6个;其余11条染色体WRKY基因的数量在1—3个不等。串联重复是导致基因家族扩张的主要方式之一,对望春玉兰WRKY基因复制方式分析发现,这个基因家族中有9个串联复制事件,包含20个WRKY基因,其中7次串联复制发生在Chr14和Chr15上,这也是引起WRKY基因家族成员在染色体上分布不均匀的主要原因。
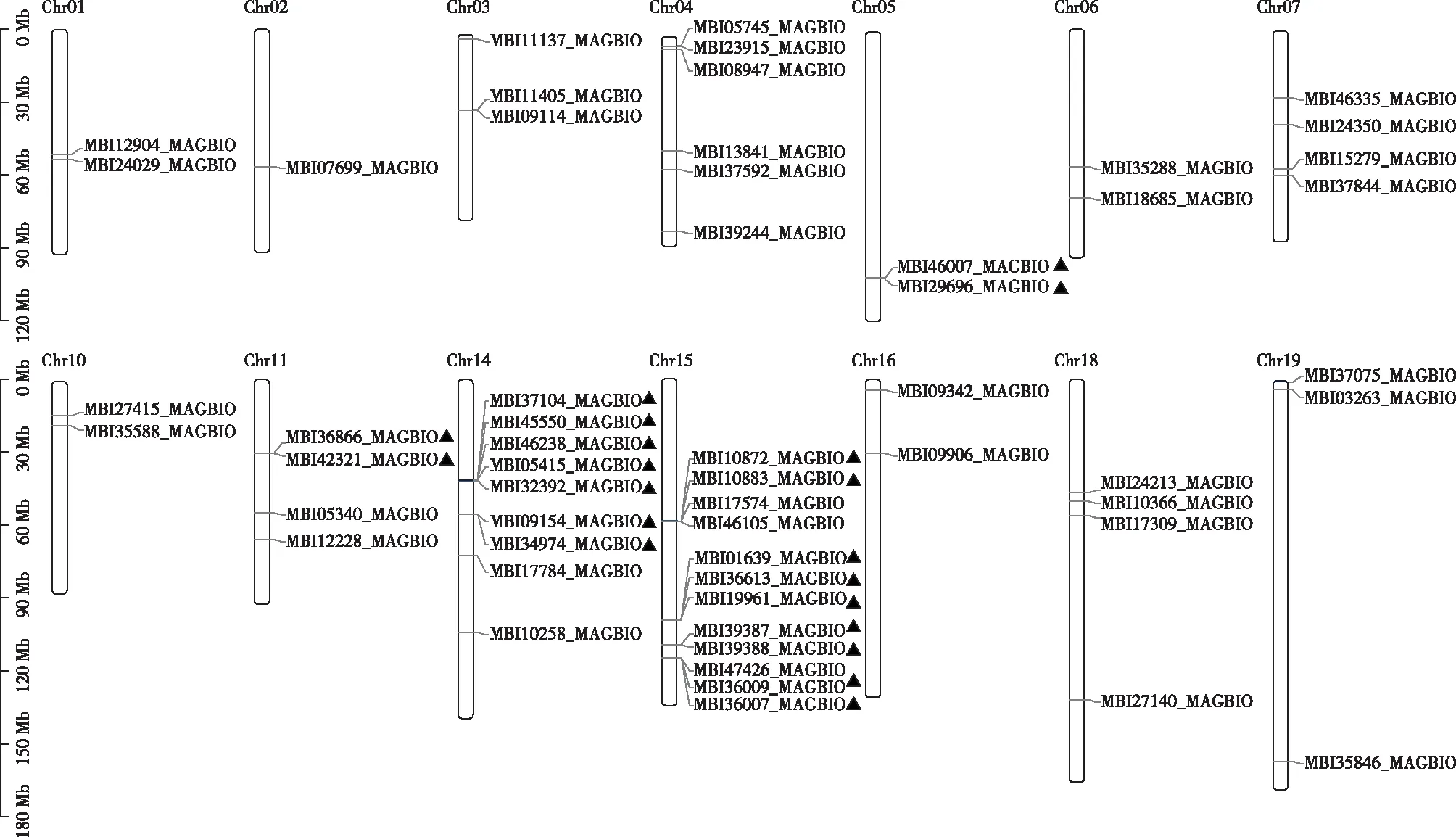
图4 望春玉兰WRKY基因家族在染色体上的分布(串联复制)
2.6 望春玉兰WRKY基因家族组织表达分析
基因表达模式可以揭示植物不同组织的生物学功能。将望春玉兰叶片、花和根系转录组测序序列分别比对到基因组,分析WRKY家族基因在不同组织中的表达模式,结果如图5所示。从图5中可以看出,Group I中除了MBI36007_MAGBIO在根系中不表达,Group IId中除了MBI36007_MAGBIO在根系和叶片中不表达外,其余基因在所有组织中均检测到表达,且Group IId中的基因整体表达量均较高。Group IIa中只有MBI32392_MAGBIO和MBI46105_MAGBIO在3个组织中均能检测到表达,有5个基因在3个组织中均未检测到表达,而MBI37104_MAGBIO和MBI05415_MAGBIO是叶片组织特异性基因,只在叶片中有表达量。Group IIb中有1个基因MBI37844_MAGBIO在3个组织中表达量均较高,FPKM值均超过10,还有3个基因只在根系特异表达。Group IIc中除了MBI24029_MAGBIO在叶片中特异表达外,其余基因在不同的组织中也都能检测到表达。

图5 望春玉兰不同组织WRKY基因家族表达模式
3 讨论
WRKY是植物中较大的基因家族,已被证实在调节植物生长、发育、非生物胁迫中起到重要作用。随着全基因组测序发展,已有几十种双子叶植物全基因组WRKY基因家族被鉴定和分析,而其基因家族中基因数量在45—188个不等[17]。本研究从望春玉兰基因组中鉴定了56个WRKY基因,比较分析发现,望春玉兰基因组(2.20 Gb)大小约是拟南芥(125 Mb)的18倍,但望春玉兰WRKY基因的数量(56个)却只有拟南芥(74个)的3/4;同样,望春玉兰基因组约是木本植物毛果杨(0.55 Gb)的4倍,但WRKY基因的数量却不足毛果杨(122个)的1/2[17]。上述结果说明了基因组中,基因家族基因的数量并不是由基因组大小决定的。
根据WRKY基因保守结构域序列和锌指结构类型,同时结合拟南芥WRKY基因家族的分类,将望春玉兰WRKY基因家族分为3个组:Group Ⅰ,Group Ⅱ和Group Ⅲ,分别含有8,38和7个基因。然而在拟南芥和毛果杨中,Group I基因的数量占主导地位[18],而望春玉兰中Group II基因数量占67.9%,说明Group II在进化过程中可能发生了更多的基因复制。这与对望春玉兰WRKY基因家族串联复制分析结果一致,在望春玉兰WRKY基因家族中共发现9个串联重复,其中Group II中有5个串联重复共产生了11个基因。望春玉兰WRKY基因家族Group II可进一步细分为IIa,IIb,IIc,IId和IIe共5个亚组,分别含有9,7,10,7,4个基因。对拟南芥、水稻、葡萄等7种植物基因组WRKY基因家族Group II基因数量分析发现,Group IIa基因的比例为8.8%—12.2%之间[18],而望春玉兰Group IIa基因的比例为23.7%,约是其他物种的2—3倍,说明Group IIa在望春玉兰中发生了大量扩张。已有研究表明,WRKY基因家族的Group IIa基因可以提高拟南芥苗期和苹果愈伤的渗透胁迫能力和抗盐性[19],也是调节水稻先天免疫能力的重要转录因子[20],望春玉兰Group IIa基因的大量扩张,可能是在长期进化过程中提高非生物胁迫的表现。
WRKY不仅调节植物生长、发育和参与非生物胁迫的调节,还参与调控植物的次生代谢。Xu等在棉花中克隆的GaWRKY1转录因子可以调控卡丁烯合酶-A的活性,从而参与调节棉花倍半萜烯类化合物的合成[5];Liu等研究发现药用植物杜仲WRKY转录因子参与苯基丙胺酸和黄酮类物质合成代谢的调控[17]。望春玉兰转录因子顺式作用元件分析发现,WRKY启动子区域不仅含有大量参与非生物胁迫的顺式作用元件,还有9个基因含有参与黄酮类物质合成代谢的顺式作用元件,其中MBI19961_MAGBIO和MBI08947_MAGBIO分别在根系和花中高表达,而根系和花是望春玉兰药物合成的主要组织,因此这2个基因可能影响望春玉兰根系和花的次生代谢。
