石斛花形态的系统发育保守性及性状关联
2022-01-19王晓静冯春慧刘振亚
孙 梅,田 昆,,王晓静,冯春慧,刘振亚
(1.西南林业大学 国家高原湿地研究中心,云南 昆明 650224;2.中国科学院 西双版纳热带植物园 热带森林生态学重点实验室,云南 勐腊 666303;3.云南省滇池生态定位观测站,云南 晋宁 650600)
石斛属(Dendrobium)植物是兰科(Orchidaceae)最大的属之一,拥有1 200 多种.由于本属植物花色艳丽多样,花具香味,花期长,具有重要的观赏价值,被誉为“四大观赏洋兰”之一,因而在园艺上被广泛栽植[1-2].由于本属植物为附生植物,野外生境要求较为苛刻,且对环境和人类干扰十分敏感[2-4].因此,过去数十年的大量开发利用,导致本属植物在野外天然生境下的数量急剧减少[5].利用分子技术从基因水平上进行品种改良,将会因为野外物种基因库的减少受到限制.而通过在植物的耐受范围内控制生长小气候、养分等环境因子,并利用植物性状对这些环境因子的适应性来改变某些花性状,进而达到品种改良的目的,是对本属植物进行品种改良的发展方向.通过控制环境因子对花性状进行品种改良,首先需要确定花各部分结构的性状可变性,进而选择对环境因子的变化较为敏感的性状,即可变性较高的性状作为品种改良的方向.
花是植物重要的繁殖结构,对植物在其进化过程中适应多变环境以及成功繁殖具有关键作用[6].研究花性状的适应进化是了解花性状变化趋势及适应策略的重要基础[7-8].石斛属植物化石最早记录于中新世早期,其花形态在植物的进化发展过程中发生了多次演化,对植物适应多样复杂的环境十分重要[9].了解本属花不同的性状的进化保守性以及性状间的进化关联性,不仅是了解花结构适应环境变化的基础,也是园艺学上对多个花性状进行同时改良的依据,但是目前还没有查阅到本属植物花性状的相关保守性信息.性状主要受系统发育影响是引起植物性状较为保守、可塑性较小的重要原因[10].前期通过对石斛属叶片的研究发现,石斛属植物叶片的形态结构和元素含量受系统发育和环境的共同影响,一些性状如叶片密度、上表皮厚度、气孔指数、比叶重、叶片干物质含量以及C、N、P 含量等表现出强烈的系统发育保守性,并且不同形态结构性状以及经济性状间均存在显著的关联进化[11-12].对兰科兜兰属花形态的保守性研究也发现,唇瓣形状、花瓣形状、花瓣宽度以及花的数目表现出强烈的系统发育保守性[7],花寿命与单位面积的花干重之间存在显著正相关[8].据此推测,石斛属植物的花形态性状中可能有一些具有系统发育保守性,并且不同花形态性状间可能存在关联进化,协同适应传粉者和外界环境的变化.
系统发育独立对比分析(phylogenetically independent contrast analysis,PIC)是近年来研究性状的进化保守性和关联适应性的重要方法[10-13].该方法认为:生物的系统发育存在时间序列上的自相关,如果物种的性状受系统发育影响较强,那么物种在进化时间上较相近,生态适应性相似,性状也较保守,可变性较低[10].该方法也可以用来检测两个性状间的进化关联.性状进化关联的系统发育影响较强时,两个性状的种平均值相关关系(species means)和去除系统发育影响后的PIC 对比值(contrasts)相关关系一致;而当这两种关系不一致时,意味着重要的生态学意义,表明是对环境辐射适应的结果,其中PIC对比值间的关系是对演化历史的反映,因此将更有意义[14].
本研究选择8 种典型的、观赏价值较高的石斛属植物为研究对象,通过构建其系统进化树,测定其8 个关键的花性状,检测花性状的系统发育信号,并应用PIC 分析的方法,检测花性状间的物种平均值相关性和PIC对比值相关性,探讨石斛属植物花性状的系统发育保守性石斛属植物花各性状的关联适应性.本研究将为了解花性状的环境适应策略、园艺学筛选易变的花结构进行花品种改良以增加石斛花的观赏性等方面提供理论依据.
1 材料和方法
1.1 研究地点和实验材料本研究在中国科学院西双版纳热带植物园(XTBG;北纬21.41°,东经101.25°;海拔570 m)内的野生兰花园开展.该植物园位于我国云南省西南地区,受亚洲西南季风气候的影响,一年分为典型的雨季(5月至10 月)和旱季(11月至次年4月).根据植物园内版纳生态站气候数据记录,XTBG所在位置年平均气温21.7 ℃,最热月(7 月)平均气温25.5 ℃,最冷月(1 月)平均气温14.8 ℃;年平均降水量约1 560 mm,其中85 %的降水发生在雨季.在此野生兰花园内,原生的毛紫薇(Lagerstroemia villosa Wall.ex Kurz.)、川楝(Melia toosendan Sieb.et Zucc.)、潺槁木姜子(Litsea glutinosa(Lour.)C.B.Rob.)、圆锥木姜子(Litsea liyuyingi Lion.)、大黄栀子(Gardenia sootepensis Hutchins.)以及铁力木(Mesua ferrea L.)等木本双子叶植物作为石斛属以及其他附生兰科植物的附主.
为使环境效应的影响最小,选择已经在野生兰花园内生长了多年的8 种石斛属植物为研究对象,分别为报春石斛(Dendrobium polyanthum Lindl.)、兜唇石斛(Dendrobium cucullatum(Roxb.)C.E.Fischer)、美化石斛(Dendrobium loddigesii Rolfe)、玫瑰石斛(Dendrobium crepidatum Lindl.ex Paxt.)、鼓锤石斛(Dendrobium chrysotoxum Lindl.)、流苏石斛(Dendrobium fimbriatum Hook.)、球花石斛(Dendrobium thyrsiflorum Rchb.f.)和小黄花石斛(Dendrobium jenkinsii Lindl.).经过多年的生长,这些植物已经充分适应了野生兰花园内相似的生长环境,且每个所选种在野生兰花园内至少有6 株长势良好的植株个体,以保证实验的样本量.
1.2 花性状测定于植物花期(3—4 月),每个种选择6 株长势良好的植物为研究材料,每株植物随机选取5 朵新鲜的花为研究对象.沿花梗基部取下花,并迅速放入内含湿纸球的自封袋内,挤出袋内空气并密封后放入保鲜盒内,带回实验室迅速进行花性状测定.
在实验室中,用游标卡尺测量并记录花开口大小(flower opening size,FOS).然后,分解花的各部分结构,用游标卡尺测量花梗长(flower pedicel length,FPL)、花梗宽(flower pedicel width,FPW)以及萼囊深(flower calyx sac depth,FCSD).将每一片花瓣、唇瓣和萼片分别过Li-Cor 3000A 叶面积仪,直接记录唇瓣面积(flower lip area,FLA)和中萼片面积(flower middle sepals area,FMSA),花瓣面积(flower petals area,FPA)和侧萼片面积(flower lateral sepals area,FLSA)分别计算为其两片花瓣和两片侧萼片面积的平均值.
1.3 系统进化树系统进化树基于拼接的核ITS序列以及叶绿体rbcL、matK、ycf5 序列重建而成,这些基因序列下载自GenBank(http:/ /www.ncbi.nlm.nih.gov)网站,基因序列登记号见表1.
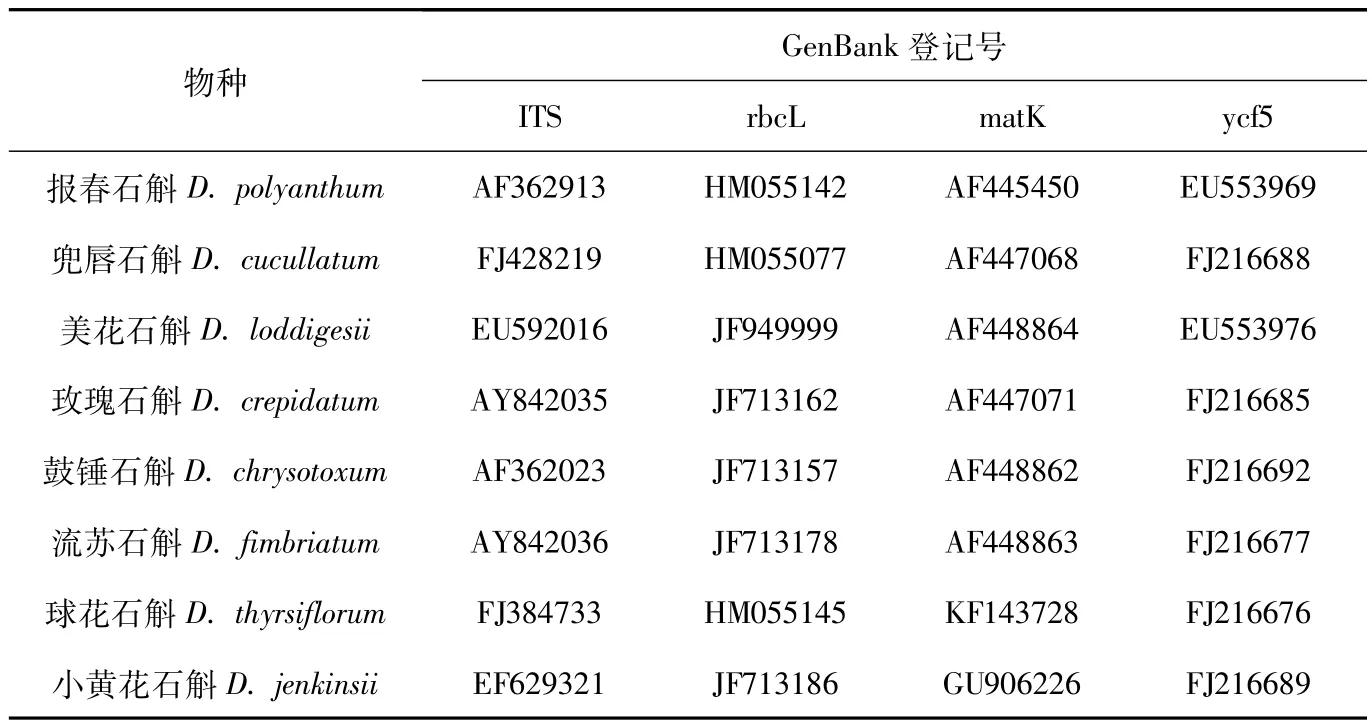
表1 本研究石斛属植物的GenBank登记号Tab.1 GenBank accession numbers of Dendrobium species in this study
由于石豆兰属(Bulbophyllum)与石斛属植物的亲缘关系较近[15],本研究选择密花石豆兰(Bulbophyllum odoratissimum)作为构建系统进化树的外类群.用MEGA v.5.0 软件的“CLUSTALW”模块进行序列比对,并对某些位点做手动轻微的调节.使用PAUP v.4.0b10 软件进行进化一致性分析.通过Modeltest v.3.7 软件由“Akaike”信息准则进行最优模型选择.本研究数据集的最优模型是GTR(generalized time reversible)+G(gamma shape)模型.用MrBayes v.3.2 软件,并用“贝叶斯法”对基因序列矩阵进行系统发育分析.在贝叶斯分析中,系统进化树通过运行马尔科夫链(Metropolis-coupled Monte Carlo Markov,MCMC)生成,每运行100 次选择一棵树,共运行100 万次.用后置检验估计节点的稳定性.
1.4 数据分析数据均由R v.3.01(http:/ /ftp.ctex.org/mirrors/CRAN/)进行分析.分析前,所有的花性状数据均进行log10 转化以增加正态性和方差齐性.为了评估花功能性状的系统发育历史,首先基于“布朗运动”模型,并以“K-统计”检测了这些花性状的系统发育信号(K 值).K >1 表明性状比布朗运动期望的随机值更加保守,受系统发育的影响较强;K <1 表明性状的可变性较高,受系统发育的影响较弱;而K =1 则表明性状与期望值一致,服从随机变化[7].K 统计及其相对应的P 值由R统计中“Picante”程序包估计[16].
系统发育独立对比分析(PIC)首先用Phylocom软件的“analysis of traits(AOT)”模块计算叶片性状的节点对比值[17].在系统进化树中,每个节点的性状对比值都是相互独立的,作为更内部节点的加权值,并计算为该节点后两个姐妹种的性状值差异,然后将这些加权值除以期望的变化量即分开两个物种的枝长的平方根.最后,这些对比提供了N-1(N指物种数目,本研究中N =8)个独立的数据点,每个点都代表了一个进化分歧[18].由R 程序包中的“Pearson-相关分析”分别计算PIC 分析前、后的花性状间的相关性.
2 结果
2.1 性状的系统发育保守性本研究8 个花性状沿系统进化树明显变化(图1),并且所有花性状的系统发育信号(K 值)均>0.8,暗示系统发育影响石斛花性状的变化,其中,花开口大小FOS(K =1,P =0.049)、花瓣面积FPA(K =1.04,P =0.041)、中萼片面积FMSA(K =1.08,P =0.046)以及花梗宽FPW(K =1.02,P =0.048)的K值均>1,表现出强烈的系统发育保守性;而唇瓣面积FLA、侧萼片面积FLSA、花梗长FPL和萼囊深FCSD的K <1,表明这些性状的可变性相对较高,受环境因素的影响较大(表2).

表2 石斛属植物的8 个花性状的变化及系统发育信号(K-统计)Tab.2 Variations and phylogenetic signals(K-statistics)for eight flower traits of Dendrobium species
图1 中,系统进化树上节点的数值代表植物类群在该节点处分开的支持率大于50%,系统进化树枝上的数值代表两个相邻节点间的距离,密花石豆兰是外类群.

图1 本研究采用的系统进化树及其对应的8 种石斛属植物的花性状值Fig.1 The phylogram and its eight corresponding Dendrobium species with their flower traits in this study
2.2 性状间的进化关联基于物种的花相关性分析显示,花表观大小性状,包括花开口大小、花瓣面积、唇瓣面积、中萼片面积及侧萼片面积间均存在显著正相关(表3).但去除系统发育的影响后,仅中萼片面积与花瓣面积、侧萼片面积之间仍然显著正相关,而其他性状间的关系均不再显著(图2).

表3 8 个花性状的物种间相关性及系统发育独立对比(PIC)关系Tab.3 Pairwise cross-species and phylogenetically independent contrast(PIC)correlations among the eight flower traits

图2 FMSA、FPA、FLSA间的物种相关性及系统发育独立对比关系Fig.2 Pairwise cross-species correlations and phylogenetically independent contrast correlations between FMSA,FPA and FLSA
无论是否考虑系统发育的影响,花梗宽与花瓣面积、中萼片面积、侧萼片面积之间均存在显著正相关(图3).

图3 FPW与FPA、FMSA、FLSA之间的物种相关性以及系统发育独立对比关系Fig.3 Pairwise cross-species correlations and phylogenetically independent contrast correlations between FPW and FPA,FMSA as well as FLSA
去除系统发育的影响之后,中萼片面积、花梗宽均与萼囊深之间存在显著负相关,而花梗长与萼囊深之间的相关性不再显著(图4).
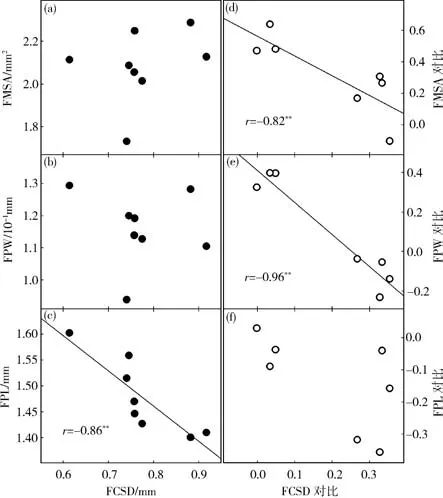
图4 FCSD与FMSA、FPW、FPL之间的物种相关性以及系统发育独立对比关系Fig.4 Pairwise cross-species correlations and phylogenetically independent contrast correlations between FCSD and FMSA,FPW as well as FPL
本文图表中每一组关系的显著性水平:*表示P <0.05,**表示P <0.01,***表示P <0.001.
3 讨论
石斛属植物花形态性状表现出较强的系统发育信号(K >0.8).前期对本属植物叶片功能性状的研究也得出相似的结果,即石斛属植物的形态结构、生物量和元素含量的系统发育信号也较强[11-12].这些研究相互补充、相互验证,表明石斛属植物的功能性状受环境和系统发育的共同影响,在研究本属植物的功能适应性时应该考虑系统发育的因素.石斛属植物的最早化石记录为中新世早期[9],随后在漫长的地质长河里经历了多次演化,形成了如今种类繁多的附生植物类群[19].
石斛属花开口大小、花瓣面积、中萼片面积、花梗宽表现出强烈的系统发育保守性(表2),表明这些花性状的变化遵循随机演化规律,受环境的影响较小,可能对维持植物基本的生存和繁殖具有重要意义[20-21].相反,受系统发育影响相对较小的其他性状,如唇瓣面积、侧萼片面积、花梗长和萼囊深,则受环境的影响较大,这些性状的变化能够促使植物快速、有效地适应环境变化,引起适应辐射[22].在对石斛属花性状的选择性改良上,若仅采取改变生长条件的方式进行,宜选取这类受系统发育影响较小的性状作为品种改良的方向.
植物性状间的关联暗示了这些性状对植物功能的共同作用效果,进而反映了植物对环境的功能适应策略[11,18,23].花表观大小性状间均存在显著正相关(表3),但去除系统发育的影响后,仅中萼片面积与花瓣面积、侧萼片面积间仍然显著正相关,而其他性状间的关系均不再显著(图2).石斛属花表观大小是关系其传粉机制和繁殖是否成功的重要性状[24-25],这些性状间的相关可能是在细胞的生长和伸长过程中形成的自相关[26].尤其是根据植物花的发育过程,花瓣和萼片是同源,有相同的基因编码,后经分化分别形成萼片与花瓣[26-27].它们二者之间的正相关在基因上即有所表现.性状间的系统发育独立对比关系(contrasts)与物种值相关性不同,表明这些关系是在植物环境适应变化的过程中逐渐建立起来的,对植物的适应辐射具有促进意义[7,22].而中萼片面积与花瓣面积、中萼片面积与侧萼片面积间的相关则可能对花功能的发挥具有更加重要的意义.可以想象,石斛属在形成的初期(物种起源时期),对于花传粉机制起到关键作用的是中萼片面积与花瓣面积以及侧萼片面积间的协同相关,并且这两个关系在石斛属后来的进化历程中一直发挥作用[7,11,28].
石斛属植物的花通过花梗与花序或者茎干相连,花梗承担了整个花的质量[29].花瓣、萼片占据着花质量的主要部分.类似叶柄对叶片质量的支撑作用,花梗长和花梗宽都将对承担花质量发挥作用,一般来说,较短、较粗的花梗产生的伸张压力较小,能够承担更重的质量.花梗宽与花瓣面积、中萼片面积、侧萼片面积之间均存在显著正相关(图3),表明相比于花梗长,石斛属植物花梗宽在承担花质量上发挥主要作用.萼囊深对于石斛属植物调节花的角度,促进不同昆虫对花进行传粉具有重要作用[30].萼片在花的发育过程中最先发生,中萼片对于保护花被,维持花的完整性,、维持花的张开状态以及吸引传粉昆虫等方面意义重大[26].中萼片面积、花梗宽均与萼囊深的系统发育独立对比值之间存在显著负相关(图4),反映了石斛属植物在维持花的张开和承担花的质量与花角度调节方面存在权衡.不同性状在对同一个功能组合发挥作用时,常常表现出权衡,例如叶片在水分传输和散失之间的权衡[31],花在授粉投入与果实生产之间的权衡[32]等.萼囊的形成和花角度调节需要的能量投入可能较高,萼囊与中萼片和花梗宽间的关系,也可能暗示了花在具体是往增加花大小方面投入能量还是往增加角度调节力方面投入能量间的权衡.二者体现了花的不同传粉策略,反映了石斛属植物在进化历程中一直通过权衡二者间的能量投入,实现了不同小生态位花的复杂传粉和繁殖机制[6].
综上所述,石斛属植物花形态性状受系统发育的影响,花开口大小等性状表现出强烈的系统发育保守性对维持植物基本的生存和繁殖具有重要意义;而系统发育信号较弱的性状可能是促使植物快速、有效适应环境变化.中萼片面积与花瓣面积、侧萼片面积间的相关性将对传粉具有更加重要的意义.萼囊与中萼片、花梗宽间的显著负相关,暗示了花的能量分配策略,体现了花的不同传粉策略,反映了石斛属植物在进化历程中不同小生态位花的复杂传粉和繁殖机制.本研究对了解花性状的传粉和环境变化适应策略以及园艺学筛选易变的花结构进行花品种改良具有重要指导价值.
