濒危植物崖柏回归苗木存活和生长与环境因子的关联性*
2022-01-18马凡强郭泉水秦爱丽简尊吉黄吉勇王中兵张世强
马凡强 郭泉水 秦爱丽 简尊吉 黄吉勇 王中兵 杨 泉 张世强
(1.中国林业科学研究院森林生态环境与保护研究所 国家林业和草原局森林生态环境重点实验室 北京 100091;2.重庆市开州区国有马云林场 重庆 404500;3.重庆市开州区林业局 重庆雪宝山国家级自然保护区管理局 重庆 404500;4.重庆大巴山国家级自然保护区管理局 重庆 405900)
回归指在一个物种出现濒危的现有分布区域或已灭绝的历史分布区域重新建立该物种种群的活动(IUCN,1998),作为物种保护和种群恢复的重要策略之一,近年来,在越来越多的珍稀濒危植物保护实践中得到应用(周翔等,2011)。目前,全球已回归301种植物,其中,我国回归154种,并计划将10%左右受威胁的植物种回归到原生境(黄宏文,2018)。理论上,通过回归完全有可能恢复濒危植物种群,但实践中,不同植物的回归会遇到各种困难。美国、英国、澳大利亚和一些发展中国家开展珍稀濒危植物回归已有多年,但成功的甚少(周翔等,2011),我国大多数濒危植物的回归未见后续报道,其主要原因是成功的回归需开展一系列基础研究,回归后还要对回归种群进行长期监测,并及时解决回归中出现的问题(IUCN,1998;国家林业局,2016;国家林业和草原局,2020)。当前,国内外对珍稀濒危植物回归的研究很少,长期监测研究更是鲜见报道(Godefroidetal.,2011;黄宏文,2018),回归研究不足或监测缺失,不仅影响回归目标实现,而且也难以弄清回归成败的原因。
植物回归初期植株矮小,根系分布浅,对环境变化反应敏感,是回归成败的关键期(国家林业局,2016),回归中的问题大多发生在这一时期。已有研究表明,夜花树兰(Epidendrumnocturnum)植株长度若小于7 cm,则不能忍受干燥环境,存活率几乎为零(Stewart,2008);报春苣苔(Primulinatabacum)回归需有伴生苔藓护理(任海等,2003);在疏花水柏枝(Myricarialaxiflora)回归初期,如果汛期不漫灌降温就会生长不良,在春夏季不适时除草和施肥就会生长受阻(王勇等,2003)。
崖柏(Thujasutchuenensis)属柏科(Cupressaceae)崖柏属常绿乔木,为我国特有珍稀濒危裸子植物和白垩纪孑遗植物(汪松等,2004)、国家一级重点保护野生植物和国家优先抢救的极小种群野生植物。1892年,崖柏在重庆市城口县石灰岩山区被首次发现,此后“绝迹”了100多年,期间有关崖柏的研究几近空白;1999年被重新发现后(Xiangetal.,2002),众多学者围绕崖柏的拯救繁育开展了广泛深入研究(刘建锋等,2004;王祥福等,2008;金江群等,2013;Liuetal.,2013;朱莉等,2014;郭泉水等,2015;秦爱丽等,2015;马凡强等,2017;简尊吉等,2017;Qinetal.,2017;Jinetal.,2018),并于2016年野外回归了60万株。截至2019年,崖柏已回归4年,但未见对回归崖柏种群和生境及其相互关系进行系统的调查研究。
本研究采用典型抽样法,在重庆市开州区和城口县崖柏回归区布设样地,调查回归崖柏苗木的保存率和生长状况及其环境条件,通过灰色关联分析、回归分析和方差分析等,研究崖柏回归初期苗木存活和生长与环境因子的关系,揭示影响崖柏回归初期苗木存活和生长的主要环境因子,以期为回归崖柏生境管理提供科学依据。
1 研究区概况
崖柏回归区位于重庆雪宝山国家级自然保护区和大巴山国家级自然保护区及其周边。其中,雪宝山国家级自然保护区地处开州区境内(108°33′—108°53′E,31°31′—31°42′N),海拔460~2 626 m,属亚热带季风气候向暖温带季风气候过渡类型,年均气温6.0~10.0 ℃,年均降水量1 500 mm 以上(孙凡等,2006);大巴山国家级自然保护区地处城口县境内(108°27′—109°16′E,31°37′—32°12′N),在雪宝山国家级自然保护区北部,海拔754~2 686 m,气候类型与雪宝山国家级自然保护区相似,年均气温13.8 ℃,年均降水量1 261 mm(倪东萍等,2018)。2个保护区的地带性植被均为含有常绿阔叶层片的落叶阔叶林,土壤类型为山地黄棕壤和山地棕色森林土。野生崖柏分布在海拔700~2 200 m(马凡强等,2017),崖柏回归地海拔598~1 637 m。回归前期,土地利用类型多数为坡耕地,少数为有林地。回归崖柏苗木为人工繁育的3年生实生苗,回归时苗木基径8~12 mm,苗高35~45 cm。开州区和城口县分别回归50万和10万株,面积分别为253和47 hm2。
2 研究方法
2.1 样地设置与地形、植被、回归崖柏生长状况调查
采用典型抽样法,在崖柏回归区内布设142块20 m× 20 m样地(开州回归区110块,城口回归区32块)。用GPS测量样地的经度、纬度、面积、海拔、坡向和坡度;用钢卷尺和游标卡尺测量回归崖柏的苗高、地径和冠幅;通过现地观察和访问当地居民了解回归样地利用和管理方式;在每块样地内均匀布设3个1 m×1 m样方,调查植被盖度、植株平均高、优势植物种及其多度和盖度。开州和城口回归区的调查时间分别为2018年10—11月和2019年8—9月。
2.2 土壤调查与理化性质测定
在每块样地内均匀布设3个土壤取样点,首先从每个取样点0~30 cm土层取土样1 kg,然后将3个土样混合,从中取1 kg用于土壤化学性质分析;同时,从每个取样点土壤表层(0~15 cm)取1环刀土样,用于土壤物理性质测定。土壤化学性质测定指标包括全氮、全磷、全钾、有效氮、有效磷、有效钾和有机质含量和pH值,土壤物理性质测定指标包括密度、非毛管孔隙度、总孔隙度、含水率、最大持水量和毛管持水量和最小持水量。土样预处理和理化性质分析方法参照LY/T 1215—1999(国家林业局,1999),测定分析由中国农业科学院衡阳红壤实验站完成。
2.3 气候数据与来源
各回归样地气候数据源于世界气候数据库(Hijmansetal.,2005),空间分辨率为30弧秒(相当于1 km空间分辨率)。选用的气候指标包括年均气温、月均昼夜温差、最冷月最低气温和年均降水量。
2.4 数据处理
运用Excel 2013软件整理样地环境资料、苗木保存率和生长指标(表1、2、3),采用SPSS 19.0软件进行数据处理和统计分析,将苗木保存率、生长指标和环境因子指标视为灰色系统。参考序列由保存率和生长指标构成,比较序列由环境因子指标构成,参照杨海宽等(2008)、刘思峰等(2014)、Du等(2020)分析方法,计算参考序列和比较序列的灰色关联度;对崖柏保存率和生长指标与定量的主要环境因子指标进行回归分析,同时对关联度排序前7位中定性环境因子指标下的崖柏保存率和生长指标进行方差分析(one-way ANOVA),并采用Boferroni法进行多重比较。利用Origin 9.1软件制图。

表1 崖柏回归样地定量的环境因子信息Tab.1 Quantitative environmental factor information of the plots of reintroduced Thuja sutchuenensis

表2 崖柏回归样地定性的环境因子信息Tab.2 Qualitative environmental factor information of the plots of reintroduced Thuja sutchuenensis

表3 回归崖柏苗木保存率和生长指标①Tab.3 Preservation ratio and growth indicators of reintroduced Thuja Sutchuenensis seedlings
3 结果与分析
3.1 回归崖柏苗木保存率和生长指标与各环境因子指标的灰色关联分析
开州和城口回归区崖柏苗木保存率和生长指标与各环境因子指标的灰色关联分析结果(表4、5)显示,开州回归区崖柏苗木保存率和生长指标与各环境因子指标的关联度为0.62~0.81,城口回归区为0.58~0.85,表明各环境因子指标对回归崖柏保苗木存率和生长都有一定影响。
从关联度排序结果(表4、5)可以看出,位居不同回归区首位的环境因子指标有所不同:在开州回归区为回归地利用和管理方式,在城口回归区为气候因子中的最冷月最低气温。2个保护区关联度排序前7位的环境因子指标具有明显共性:位居2个回归区前3位的环境因子指标均包括气候因子中的最冷月最低气温和年均气温,只是城口回归区分别排在第1位和第2位,开州回归区分别排在第2位和第3位;位居第4位的环境因子指标均为地形因子,开州回归区为坡向,城口回归区为坡度;位居第5~7位的大多数环境因子指标均为土壤因子,开州回归区为土壤有效钾含量、土壤有效氮含量和土壤总孔隙度,城口回归区为土壤全钾含量和土壤非毛管孔隙度。但关联度排序第7位之后的其他环境因子指标在2个保护区的位序对应性较差,共性也不明显。基于各环境因子指标关联度排序位置以及是否在2个回归区具有共征认为,关联度排序前7位的环境因子指标中,回归地利用和管理方式是对开州回归区崖柏苗木保存率和生长影响最大的环境因子指标,在城口回归区以气候因子中的最冷月最低气温影响最大;其次是气候因子中的年均气温,地形因子中的坡向、坡度以及土壤因子中的主要土壤理化指标;相对而言,关联度排序第7位之后的环境因子指标影响较小。
3.2 关联度排序前7位的定量环境因子指标与崖柏苗木保存率和生长指标的回归分析
开州和城口回归区关联度排序前7位(表4、5)的定量环境因子指标与崖柏苗木保存率和生长指标的线性回归分析结果(表6、7)显示,回归拟合方程的决定系数(R2)在开州回归区最大为0.28,在城口回归区最大为0.29。决定系数是因变量波动中能用自变量解释的比例,R2<0.01反映崖柏苗木保存率和生长指标波动能用环境因子解释的比例极低,决定系数普遍较低说明崖柏苗木保存率和生长受环境因子影响较多且影响较为平均。在开州回归区(表6),苗高与最冷月最低气温、年均气温显著正相关(P<0.05);地径与最冷月最低气温、年均气温极显著正相关(P<0.01);冠幅与土壤有效钾含量、土壤总孔隙度极显著正相关(P<0.01),与土壤有效氮含量显著正相关(P<0.05),与年均气温显著负相关(P<0.05);保存率与土壤有效钾含量极显著负相关(P<0.01)。在城口回归区(表7),苗高与最冷月最低气温极显著正相关(P<0.01),与月均昼夜温差显著正相关(P<0.05);地径与最冷月最低气温极显著正相关(P<0.01);冠幅与最冷月最低气温、土壤全钾含量极显著正相关(P<0.01),与年均气温显著正相关(P<0.05)。
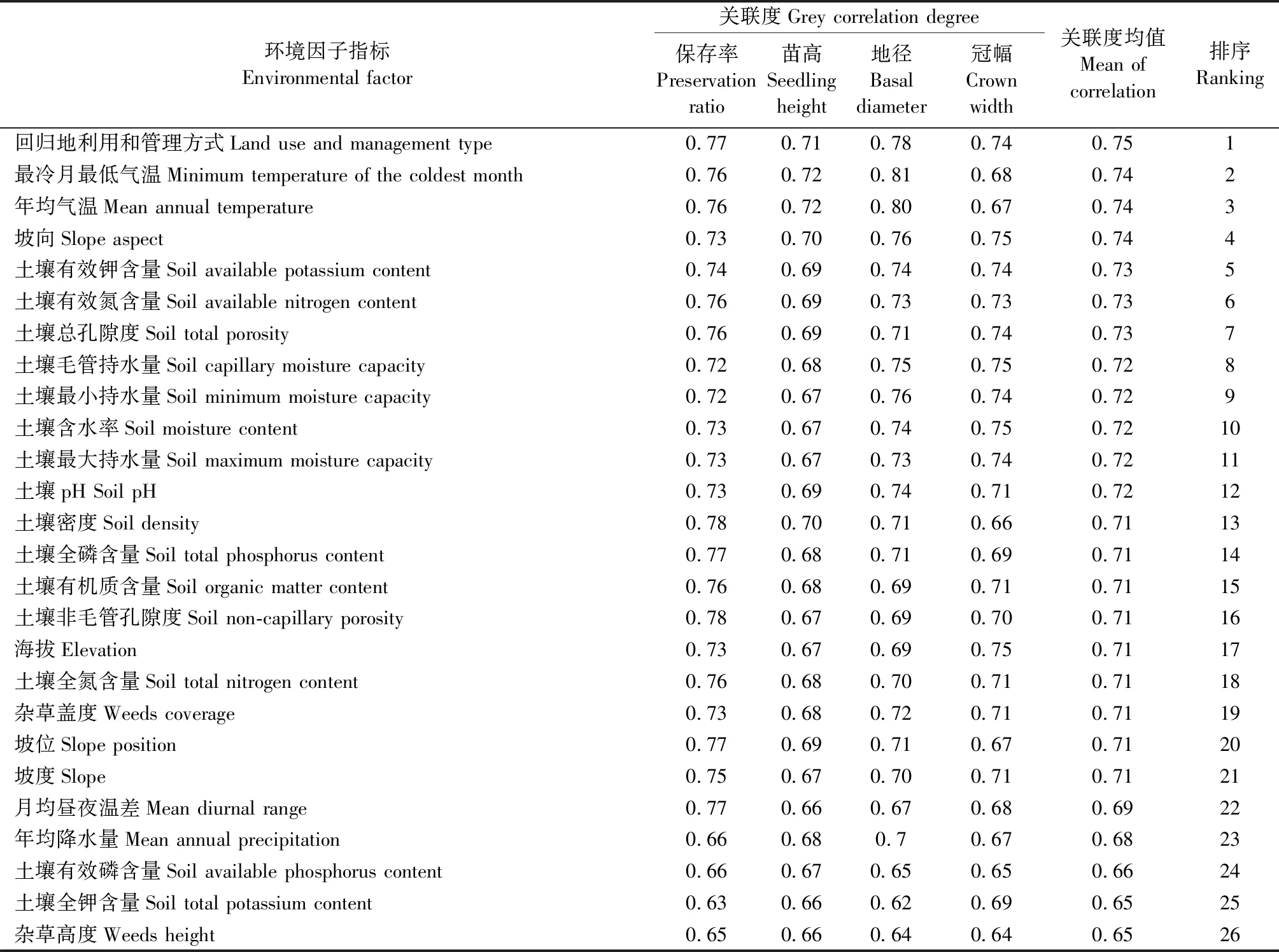
表4 开州回归区崖柏苗木保存率和生长指标与环境因子指标的灰色关联度及其排序Tab.4 Grey correlation and ranking of environmental factors with the preservation ratio and growth indicators of Thuja sutchuenensis seedlings in reintroduction area of Kaizhou

表5 城口回归区崖柏苗木保存率和生长指标与环境因子指标的灰色关联度及其排序Tab.5 Grey correlation and ranking of environmental factors with the preservation ratio and growth indicators of Thuja sutchuenensis seedlings in reintroduction area of Chengkou
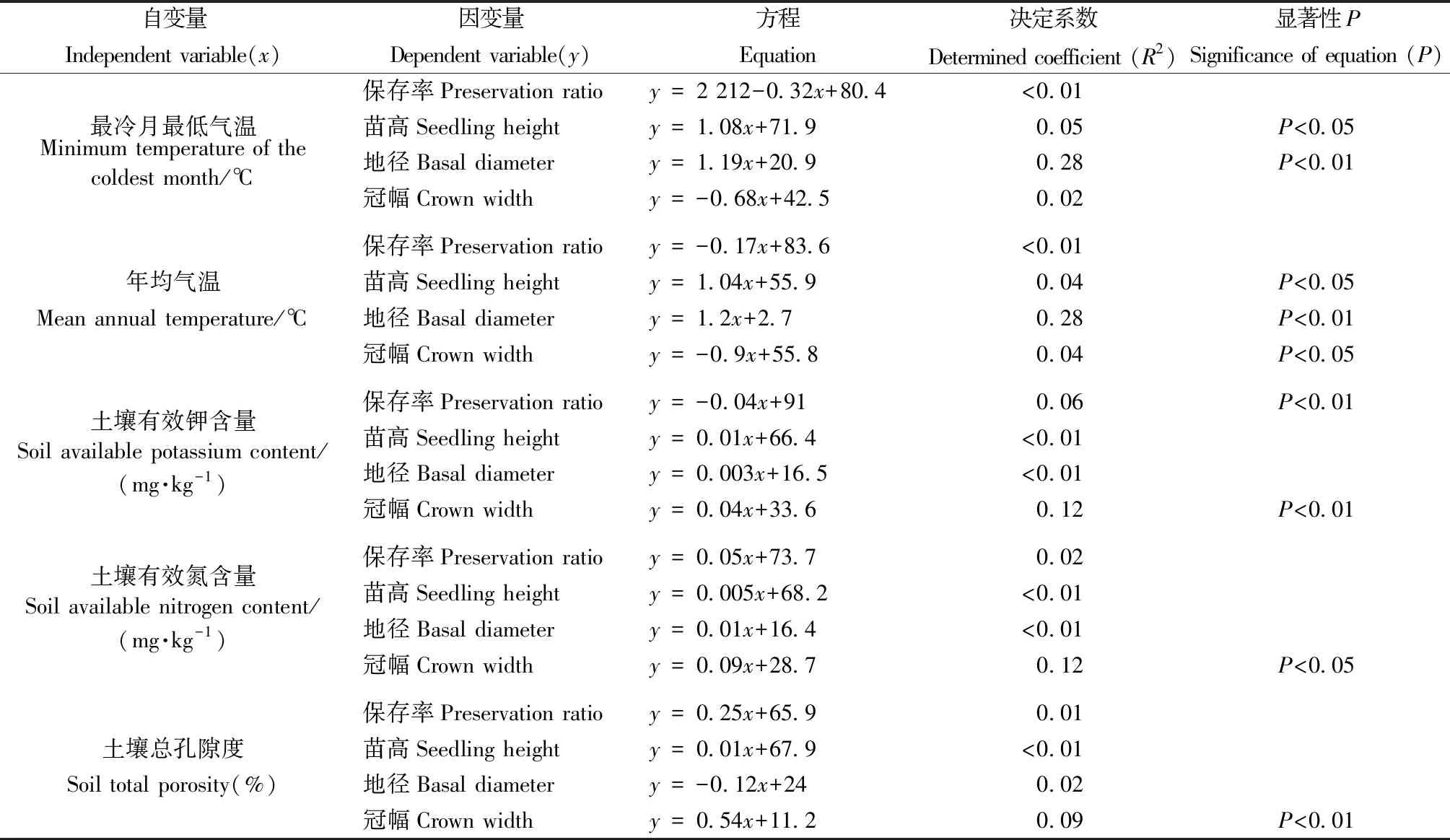
表6 开州回归区崖柏苗木保存率和生长指标与关联度排序前7位的定量环境因子指标的回归分析结果①Tab.6 Results of linear regression of preservation ratio and growth indicators of Thuja sutchuenensis seedlings with the main quantitative environmental factors in reintroduction area of Kaizhou

表7 城口回归区崖柏苗木保存率和生长指标与关联度排序前7位的定量环境因子指标的回归分析结果①Tab.7 Results of linear regression of preservation ratio and growth indicators of Thuja sutchuenensis seedlings with the main quantitative environmental factors in reintroduction area of Chengkou
3.3 关联度排序前7位中定性环境因子指标下崖柏苗木保存率和生长指标的方差分析
2个回归区关联度排序前7位的环境因子指标中仅有回归地利用和管理方式及坡向2个定性指标,且均出现在开州回归区。图1、2所示为回归地不同利用和管理方式及不同坡向崖柏苗木保存率和生长指标的方差分析和差异显著性检验结果。
由图1可以看出,回归地不同利用和管理方式下崖柏苗木保护率和苗高差异均不显著;地径表现为间作农作物>间作药材>撂荒>林冠下造林。其中,间作农作物、间作药材和撂荒地三者间差异不显著,但三者与林冠下造林差异均显著(P<0.05);冠幅表现为间作药材>间作农作物>林冠下造林>撂荒,其中,间作药材与间作农作物差异不显著,但与林冠下造林和撂荒差异显著(P<0.05)。总体而言,在间作农作物和间作药材的回归地上崖柏各项生长指标表现较好。

图1 回归地利用和管理方式对崖柏保存率和生长指标的影响Fig.1 Differences in preservation ratio and growth indicators of Thuja sutchuenensis in different type of land use and managementA:撂荒Abandoned agriculture land;B:林冠下造林Understory interplanting;C:间作药材Intercropping with medicinal materials;D:间作农作物Intercropping with crops.
由图2可以看出,崖柏苗木保存率表现为半阳坡>阴坡>阳坡>半阴坡。阴坡,阳坡和半阳坡三者间差异不显著,但半阴坡与阴坡和半阳坡差异显著(P<0.05)。阴坡苗高高于其他坡向,但不同坡向间差异不显著;地径表现为阴坡>半阴坡>阳坡>半阳坡,阴坡、半阴坡和阳坡三者间差异不显著,但阴坡和半阴坡与半阳坡差异显著(P<0.05);冠幅表现为阴坡>半阳坡>阳坡>半阴坡,阴坡与其他坡向差异均显著(P<0.05)。总体而言,崖柏各项生长指标在阴坡表现较好。
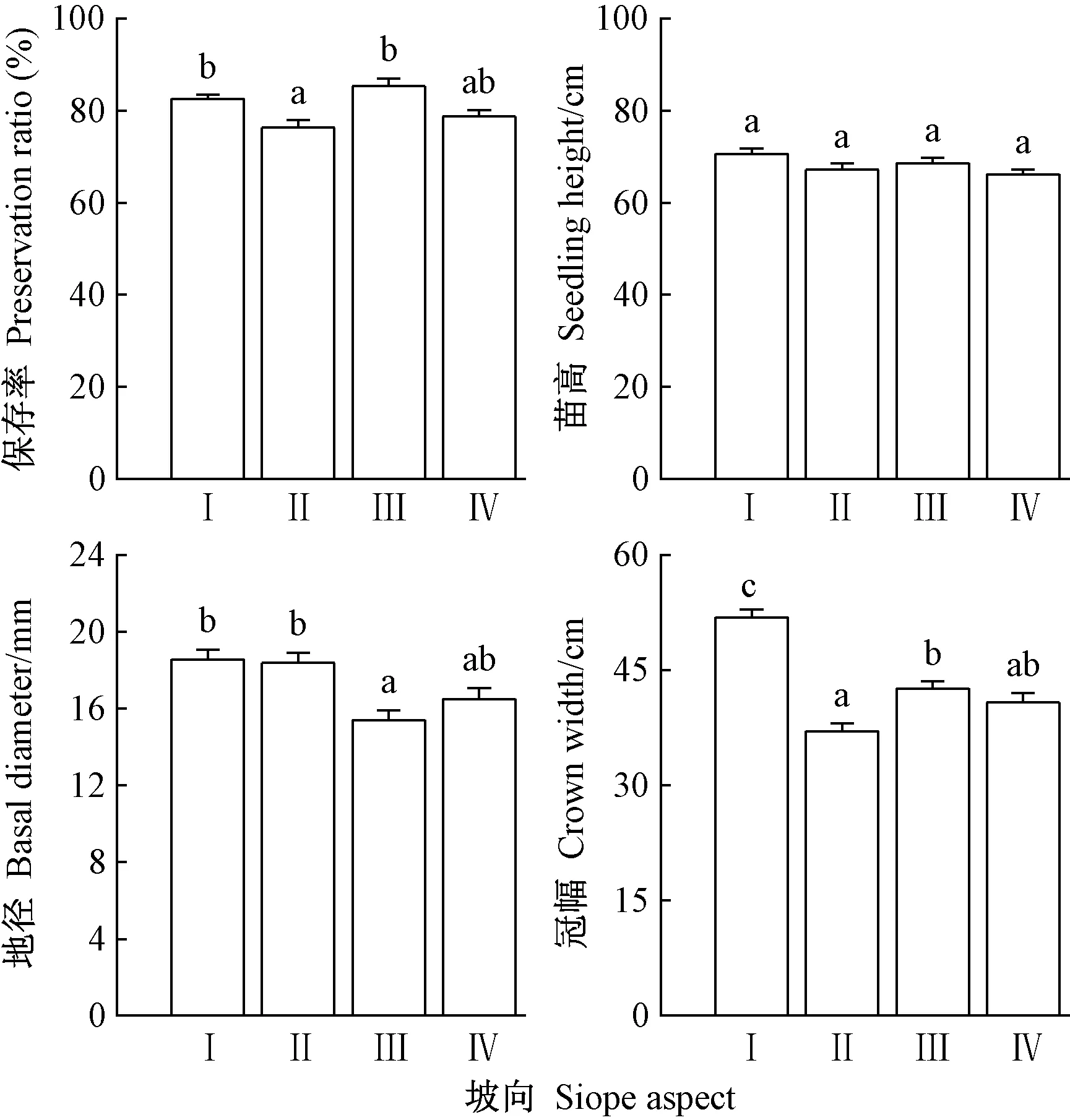
图2 坡向对回归崖柏保存率和生长指标的影响Fig.2 Differences in preservation ratio and growth indicators of Thuja sutchuenensis in different slope aspectⅠ:阴坡Shady slope;Ⅱ:半阴坡Semi-shady slope;Ⅲ:半阳坡Semi-sunny slope;Ⅳ:阳坡sunny slope.
4 讨论
运用灰色关联分析法,将苗木保存率、生长指标和环境因子指标视为灰色系统,作为整体进行比较,可以避免数据分析时因多因子孤立分散状态以及量纲不同、数值大小相差太多而难以比较的问题;另外,灰色关联度分析还具有工作量较少信息量较大的特点。根据关联度大小表示指标的重要程度,可以找出影响最大的因素,使分析简单易行(胡园春等,2020)。
温度是影响生物生理过程或参与新陈代谢的直接因子之一。在最低温度和最适温度之间,生物体内的生理生化反应和代谢活动随温度升高而增强,生长发育速度加快,低于最低温度或高于最高温度,生物则受害甚至死亡(李俊清等,2010)。朱莉等(2013)研究发现,引种栽培到北京的3年生崖柏苗木在冬季数日平均气温降至-5.0 ℃、极端最低气温达-10.0 ℃时会出现枝叶卷曲或干枯等冻害症状,到翌年春天,虽然有些枝条还会萌发,但生长速率明显下降。简尊吉等(2017)在重庆雪宝山国家级自然保护区及周边海拔178、1 360和2 250 m处布设样地,采用1.5年生崖柏容器苗作为供试材料,经过周年观测发现,海拔2 250 m处样地苗木保存率仅36%,且生长衰弱,而海拔178和1 360 m处样地苗木保存率达90%以上,且长势良好。本研究显示(表4、5),在26个环境因子指标灰色关联度排序中,气候因子中的最冷月最低气温、年均气温均位居2个保护区的第1~3位,回归分析结果(表6、7)显示,开州回归区崖柏苗木苗高、地径与最冷月最低气温和年均气温均极显著(P<0.01)或显著(P<0.05)正相关,城口回归区崖柏苗木苗高、地径和冠幅与最冷月最低气温和年均气温显著(P<0.05)或极显著(P<0.01)正相关,表明崖柏生长与气温关系密切,与前人研究崖柏苗木生长与气温关系的结论(朱莉等,2013;简尊吉等,2017)一致。然而,开州回归区崖柏苗木保存率和冠幅与气候因子中某些指标的相关性并不显著,说明开州回归区崖柏苗木生长还受其他环境因子指标的影响。
地形因子中的坡向可通过改变太阳辐射强度和降雨量再分配形成独特小气候,并间接影响土壤含水量(张昌顺等,2012;袁铁象等,2014)。阳坡太阳辐射强烈,照射时间长(McCuneetal.,2002;Heetal.,2017),气温和土温较高,土壤较干燥,阴坡则相反(刘旻霞,2017)。昝梅等(2013)研究发现,阴坡和半阴坡的植被叶面积指数明显高于阳坡和半阳坡,主要是因为阴坡和半阴坡植物蒸腾较少,土壤含水率较高,利于喜凉湿性针叶树生长。本研究表明,回归崖柏的苗高、地径和冠幅均以阴坡最大,阴坡苗木保存率略低于半阳坡,但二者差异不显著(图2),这可能与回归区坡向对小气候的影响程度以及崖柏喜温凉湿润气候的生态学特性有关。
土壤水、肥、气、热四大肥力要素是影响植物生长的关键因素(Hancocketal.,2011)。本研究结果显示(表6,7),在开州回归区,冠幅与土壤总孔隙度极显著(P<0.01)正相关,说明通气良好的土壤利于崖柏生长,冠幅与土壤有效钾含量、土壤全钾含量极显著(P<0.01)正相关,与土壤有效氮含量显著(P<0.05)正相关,表明土壤钾和氮含量增加可显著促进崖柏生长。这种现象与钾和氮对植物生长作用的理论(Tsayetal.,2011)相吻合。钾作为植物体内含量最多的阳离子和主要渗透物质,可调节细胞膨压和膜电位并激活某些酶活性(Tsayetal.,2011),还与许多和生物、非生物胁迫耐性相关的生理功能有关(Oosterhuisetal.,2014),供钾不足对植物光合作用等生理代谢活动不利,植物生长受到抑制(陈迪文等,2017)。土壤中可利用氮含量是影响植物生长的重要因子之一(LeBaueretal.,2008),氮和钾交互作用对植物生长的影响主要取决于氮的形态和浓度(Tsayetal.,2011)。
回归地利用和管理方式在开州回归区位居关联度排序首位(表4),在城口回归区则位居关联度排序第14位(表5),说明即使同一因子在不同回归区的影响程度也有所不同。回归地不同利用和管理方式下崖柏地径和冠幅差异显著,以间作农作物的各项生长指标表现较好(图1),这与崖柏苗木能够适应农作物创造的弱光环境有关。刘建锋等(2011)通过对全光、遮荫50%和遮荫25%这3种光环境下的崖柏幼苗光合能力和叶绿素荧光参数的研究发现:遮荫可导致叶片表观量子效率和最大净光合速率增加;随光强减弱,叶片光化学猝灭系数和电子传递速率逐渐降低,但叶片叶绿素含量显著增加,表明崖柏幼苗对光具有较强的耐受范围(尤其是低光)。另外,间作农作物可有效解决崖柏回归地的锄草和松土问题,对农作物的肥水管理也能促进崖柏生长。林冠下造林对崖柏生长的负面影响主要包括林木与崖柏苗木争夺水肥以及长期遮荫不利于光合产物积累2方面。回归后长期撂荒,会导致杂草丛生,回归初期崖柏苗木低矮,雨季高草倒伏会像草毯一样覆盖崖柏苗木,使其生活在高温、高湿、长期不透光的恶劣环境中,另外高大草本旺盛生长对崖柏苗木枝叶生长也起到限制作用,这可能是开州回归区崖柏苗木保存率和冠幅与年均气温未呈正相关的原因之一。
自然环境因子(如气候、地形、土壤理化性质等)对回归初期崖柏苗木保存率和生长的影响很难人为调控,但对分布在海拔高、气温较低地段的崖柏增施钾肥可促使其生长健壮,从而增强对不利环境的抵御能力。回归地利用和管理方式是可以改变的,及时清除妨碍回归崖柏苗木存活和生长的杂草灌木或间作低矮农作物,应是开州回归区确保崖柏健康生存的一项重要举措。
目前,我国对回归初期的年限界定还没有统一标准,参照我国对珍稀濒危植物回归后管理期限的要求(国家林业局,2016),崖柏回归初期监测应坚持10年左右。本研究区崖柏回归3~4年(苗龄6~7年),随着回归年限增加,影响其存活和生长的环境因子及其影响程度还会发生变化,有待长期监测。
5 结论
1)崖柏回归初期,气候、地形、土壤、生物及回归地利用和管理方式对崖柏苗木存活和生长均有一定影响,反映出环境因子的综合作用,但各环境因子指标的影响程度在不同回归区有所不同,在开州回归区以回归地利用和管理方式的影响最大,在城口回归区以气候因子中的最冷月最低气温影响最大。对整个崖柏回归区崖柏苗木保存率和生长指标影响较大的是气候因子中的年均气温,地形因子中的坡向、坡度以及土壤因子中的部分土壤理化指标,相对而言,其他环境因子影响较小。
2)开州回归区崖柏苗木苗高、地径与最冷月最低气温、年均气温,冠幅与年均气温、土壤有效钾含量、土壤有效氮含量和土壤总孔隙度以及城口回归区崖柏苗木苗高、地径和冠幅与最冷月最低气温,冠幅与年均气温,苗高与月均昼夜温差,冠幅与土壤全钾含量显著(P<0.05)或极显著(P<0.01)正相关。阴坡回归崖柏的地径和冠幅最大,半阳坡苗木保存率最高,间作农作物的崖柏苗木保存率和苗高最高、地径最大,栽植在林冠下崖柏苗木地径最小,间作药材苗木冠幅最大,撂荒苗木冠幅最小。
