青土湖不同年限退耕地植被物种多样性及土壤酶活性研究
2022-01-14王理德何洪盛宋达成闫沛迎
王 佳,田 青*,王理德,*,何洪盛,宋达成,闫沛迎
(1 甘肃农业大学 林学院,兰州 730070;2 甘肃省治沙研究所,兰州 730070;3 甘肃河西走廊森林生态系统国家定位观测研究站,甘肃武威 733000)
青土湖曾是民勤境内石羊河最大的湖泊[1],是武威市民勤县绿洲边缘的荒漠-绿洲过渡带[2]。然而,近年来,由于石羊河流域中上游地区人工绿洲的扩张,使该区生态用水日趋减少,自然植被枯萎,生态环境受到严重的破坏[3-4],使该区土地大面积弃耕而撂荒,若这些土地保护方式不当,便会加速该区风蚀沙化程度,成为新的沙尘源,形成恶性循环,严重破坏民勤绿洲生态系统[5],给当地生态和经济造成不可估量的损失,同时国内其他地区也出现同样问题[6-8]。20世纪90年代初,引起专家学者的高度重视,并开始对退耕地生态修复进行相关研究[6]。李进等[7]在河北坝上退耕地的植被研究表明“退耕的时间并不是影响植被演替与数目增加的唯一因素”。进入21世纪后,邱阳等[8]通过DCCA排序轴分类对黄土丘陵的退耕地植被演替的研究认为均匀度呈下降趋势,物种丰富度逐渐上升。可见,植被演替能促进退耕地的生态修复,实现可持续发展[9]。
植物多样性对生态系统的功能起着重要的作用[10],而植物多样性中对物种多样性的研究占有很重要的地位,是群落生态学研究的重要课题[11]。当前国内外对于植物多样性的研究有慕宗杰等[12]对沙地植被研究表明随年限的延长,沙地植物群落的物种数明显增加;张仕豪[13]对石漠化区域植被研究表明物种多样性总体随着石漠化的加剧呈下降的趋势;张巧凤等[14]对草原植被研究发现位于坡地的草原群落物种多样性普遍下降,而位于低山丘陵间平坦地带的草原群落物种多样性维持稳定或有所上升;杨晶晶等[15]对山地草甸植被研究发现放牧强度的增加能改变山地草甸植物群落物种组成;研究表明在高寒草甸区多样性指数、丰富度指数和均匀度指数总体上随退化程度的加剧呈减小的趋势[16-17],李军豪等[18]认为随高寒草甸退化程度的加剧,物种多样性总体呈减小的趋势,但在其退化前期多样性指数会有所增大。总体来说,植被演替过程比较缓慢这是所有学者的共识,只是植被自然演替特征的差异不同[19]。
土壤是构成生态系统的要素,既是反映土壤结构和水文状况及评价土壤质量的重要指标,也是植被生长发育的载体,为其生长提供所需的养分、水分和热量[20]。而植被生长又可改善土壤结构、保持土壤肥力。土壤和植被作为一个有机整体,两者相互作用、相互影响[21]。目前植被-土壤系统的相互作用在国内外的研究比较热门,赵富王等[22]对黄土丘陵区的相关研究表明,植物根系是植物与土壤进行物质交换的通道,其根系能够改善土壤结构,提高土壤稳定性。由政等[23]对于不同演替阶段群落根系分布与土壤团聚体特征的协同变化的研究发现,植物细根在群落演替以及土壤结构变化中发挥着重要的作用,土壤植被复合系统可提高土壤质量,改善土壤结构。因此,明确植被群落特征与土壤二者关系,对于退耕地植被恢复与利用具有重要的意义。土壤酶是土壤的重要组成部分[24],土壤酶活性的强弱是土壤肥力的重要指标,也是土壤健康程度的重要指标[25]。当前,关于土壤酶活性的相关研究主要涉及牧草对土壤酶活性的影响[26]、除草剂等农药施肥下土壤酶活性变化[27-28]、不同耕作方式对土壤酶活性的影响[29]、旅游踩踏对土壤酶活性的影响[30]、土壤结皮对土壤酶活性的影响[31]以及其他相关土壤酶活性的研究[32-33]。但对于不同年限退耕地土壤酶活性与植物演替相关性的研究甚少。本研究采用时空替代法,选取青土湖不同年限退耕地为研究对象,对植被物种多样性、土壤酶活性及相关性进行研究,旨在为该区生态系统的修复提供理论依据,为青土湖及其相似地区的生态环境建设提供技术支撑。
1 材料和方法
1.1 研究区概况
研究区位于甘肃省武威市民勤县东部的西渠镇,样地附近已无居民居住,地理坐标为39°01′44.11″~39°03′57.53″N、103°35′09.84″~103°37′55.49″E,平均海拔为1 305 m,东、西、北三个方向都是以以前的青土湖湖面为主,东部和北部与内蒙古自治区接壤,西靠甘肃省金昌市,南邻甘肃省武威市,多年的平均气温为7.4 ℃,极端最低气温是-28.8 ℃,极端的最高气温达38.1 ℃;年日照时数2 832.1 h;无霜期175 d,时期短,冻害天气多;全年无雨163天,年均降水量110 mm,主要集中在7-9月,占年均降雨量的73%;年平均风速2.3 m/s;沙尘暴日数26.8 d;该区气候属于温带大陆性干旱气候,干旱发生频率高,光照充足,四季分明,冬季寒冷,夏季炎热;土壤以灰棕漠土为主;地貌类型主要有3种,分别为平原地貌、风沙地貌和低山地貌;名优特产主要有:黑瓜籽、棉花、甜瓜、黄瓜;草本植物主要有碱蓬(Suaedaglauca)、骆驼蓬(Peganumharmala)、骆驼蒿(Peganumnigellastrum)、田旋花(Convolvulusarvensis)、藜(Chenopodiumalbum)、虎尾草(Chlorisvirgata)等;灌木植物主要有小果白刺(Nitrariasibirica)、红砂(Reaumuriasoongoria)、黑果枸杞(Lyciumruthenicum)、白刺盐爪爪(Kalidiumfoliatum)、柽柳(Tamarixchinensis)等。
1.2 野外调查与采样
2018年8-9月,选择未受自然因素和人为因素影响的试验区地段,退耕地时间为退耕1 a、2 a、4 a、8 a、13 a、20 a、30 a、40 a,并以耕地(CK)作为对照样地。测量并详细记录样方中物种数量、地理坐标、海拔高度(表1)。每个样地面积为100 m×100 m,在每个样地内四角和中心分别布设5个灌木样方(10 m×10 m)用于测定灌木的个体数、高度、盖度以及冠幅等指标;再在每个10 m×10 m的灌木样方内四角和中心各布设1个草本样方(1 m×1 m)用于测定草本的个体数、高度和盖度等指标。
在植被调查的同时,在样方周边分别采取剖面法和5点混合法采集土壤样品,土层分为上下2个土层(0~20 cm、20~40 cm),每个样点3次重复,筛掉土壤中的杂物,每个样品取重约1 kg,装入无菌袋放置于带冰块的泡沫箱内,带回实验室,存储在4 ℃的冰箱中,用于土壤酶活性的测定。
1.3 测定方法
1.3.1 物种多样性的测定植物物种多样性可加速恢复生态系统[34],物种多样性是植物群落的基本特征[35],因此选用Shannon多样性指数、Simpson优势度指数、Pielou均匀度指数、Margalef丰富度指数和重要指等对青土湖区不同年限退耕地的植被群落进行测量计算。计算公式分别为:
重要值:Ⅳ=(相对多度+相对盖度+相对高度)×100/3[36]

表1 不同年限退耕地的基本特征
Pi是频度,Ni是第i个物种的个体数,S是物种数量,N是所有物种的总个体数[37]。
1.3.2 土壤酶活性的测定(1)土壤过氧化氢酶活性采用容量法测定[38]。用A表示滴定25 mL原始的过氧化氢混合液所消耗的高锰酸钾量(毫升数),用B表示滴定土壤滤液所消耗的高锰酸钾量(毫升数),用T表示高锰酸钾滴定度的矫正值,代表高锰酸钾标定后的真正浓度与试验所需配制的高锰酸钾的浓度(即0.02 mol/L)的比值。(A-B)×T即为过氧化氢酶活性,表示30 min后1 g土壤的0.02 mol/L高锰酸钾的毫升数。
(2)土壤脲酶活性采用靛酚蓝比色法测定[39]。酶活性用24 h后1 g土壤中NH3-N的毫克数表示。
NH3-N(mg)=a×V×n/ma为从标准曲线中查得的NH3-N毫克数;V为显色液体积(50 mL);n为分取倍数;m为烘干土的质量。
(3)土壤蔗糖酶活性使用3,5-二硝基水杨酸比色法测定[39]。蔗糖酶活性以24 h后1 g土壤葡萄糖的毫克数表示。
葡萄糖(mg)=a×V×n/ma为标准曲线中葡萄糖毫克数;V为显色液体积(50 mL);n为分取倍数;m为烘干土的质量。
(4)土壤磷酸酶活性使用磷酸苯二钠比色法测定[39]。磷酸酶活性以24 h后1 g土壤中释出的酚的毫克数表示。
酚(mg)=a×V×n/ma为标准曲线中酚毫克数;V为显色液体积(50 mL);n为分取倍数jm为烘干土的质量。
1.4 数据处理与分析
Excel 2013软件用于基础数据处理,SPSS 20.0软件用于单因素方差分析、相关性分析,在差异显著条件下采用LSD进行多重比较。
2 结果与分析
2.1 青土湖区不同年限退耕地植物群落物种组成及重要值
在青土湖退耕40 a的植被自然演替过程中,调查的9个样地共出现16科32属42种植物,其中一年生草本(15种)占总物种数的35.71%、多年生草本(16种)占总物种数的38.10%、灌木(8种)占总物种数的19.05%,木质落叶大藤本、落叶乔木和缠绕草质藤本各1种占总物种数的7.14%,而且黎科(8属13种)、禾本科(7属7种)、蒺藜科(2属5种)3种植物种类占总物种数的59.52%,分别占总种属数的25%、21.88%和6.25%;剩余物种包括十字花科、菊科、柽柳科分别为2属2种,茄科为1属2种共占总物种数的19.05%。其余物种如白花丹科(黄花补血草Limoniumaureum)、豆科(甘草Glycyrrhizauralensis)、胡颓子科(沙枣Elaeagnusangustifolia)、兰雪科(玛多黄花补血草Limoniumaureumvar.maduoensis)、萝藦科(鹅绒藤Cynanchumchinense)、葡萄科(地锦Parthenocissustricuspidata)、伞形科(茴香Foeniculumvulgare)、十字花科(独行菜Lepidiumapetalum)和紫草科(砂引草Tournefortiasibirica)分别为1科1属。
不同年限退耕地植被群落的物种组成和重要值见表2,在退耕1 a后,田间大部分为一年生草本[稗子草(Echinochloacrus-galli)、藜(Chenopodiumalbum)和地肤(Kochiascoparia)等]和少数多年生草本植物[赖草(Leymussecalinus)、田旋花(Convolvulusarvensis)、山苦荬(Ixerischinensis)和细叶骆驼蓬(Peganumharmala)等],并伴有少数灌木,如盐爪爪(Kalidiumfoliatum)、柽柳(Tamarixchinensis)等植物;退耕2 a后,一年生草本稗子草、盐生草、地肤、藜和狗尾草的重要值明显减少为10.726、7.716、6.334、7.599、0.412;退耕4 a后,植物群落中一年生和多年生草本植物种数量明显减少,其重要值也随之降低。碱蓬、田旋花等植物的重要值减小为1.212和2.467;退耕后8~20 a,群落内灌木植物的数量逐渐增多,但植物种类总数量较稳定,木质落叶大藤本地锦和小灌木红砂等的植物首次出现,其中一年生草本,如藜、地肤、碱蓬、独行菜等植物完全退出,群落优势种由草本植物逐渐转化成灌木植物,盐爪爪、黑果枸杞等灌木植物成为群落优势种,物种数由退耕1 a的19种降低到退耕20 a的13种;退耕30 a后,物种数下降到5种,主要以黑果枸杞和盐爪爪灌木植物为主,植物群落相对稳定;退耕40 a时耐盐碱植物盐爪爪成为该地的优势种,其重要值为36.008。
2.2 不同年限退耕地植被群落的物种多样性变化
由表3可知,在退耕40 a的自然恢复过程中,个体数(N)、物种数(S)和Margalef丰富度指数随退耕年限增加呈波动式下降的趋势,其中Margalef丰富度指数在退耕1~4 a中呈上升的趋势,在退耕4 a时达到最大值2.194,在退耕4~40 a中缓慢地下降。Shannon多样性指数和Pielou均匀度指数在退耕1~4 a中不断增加,在退耕4~13 a里逐渐减小,在退耕20 a后慢慢趋于稳定。Simpson优势度指数在退耕13 a时达到最大值0.412,但整体变化幅度较小,最终呈稳定的趋势。
2.3 不同年限退耕地土壤酶活性变化
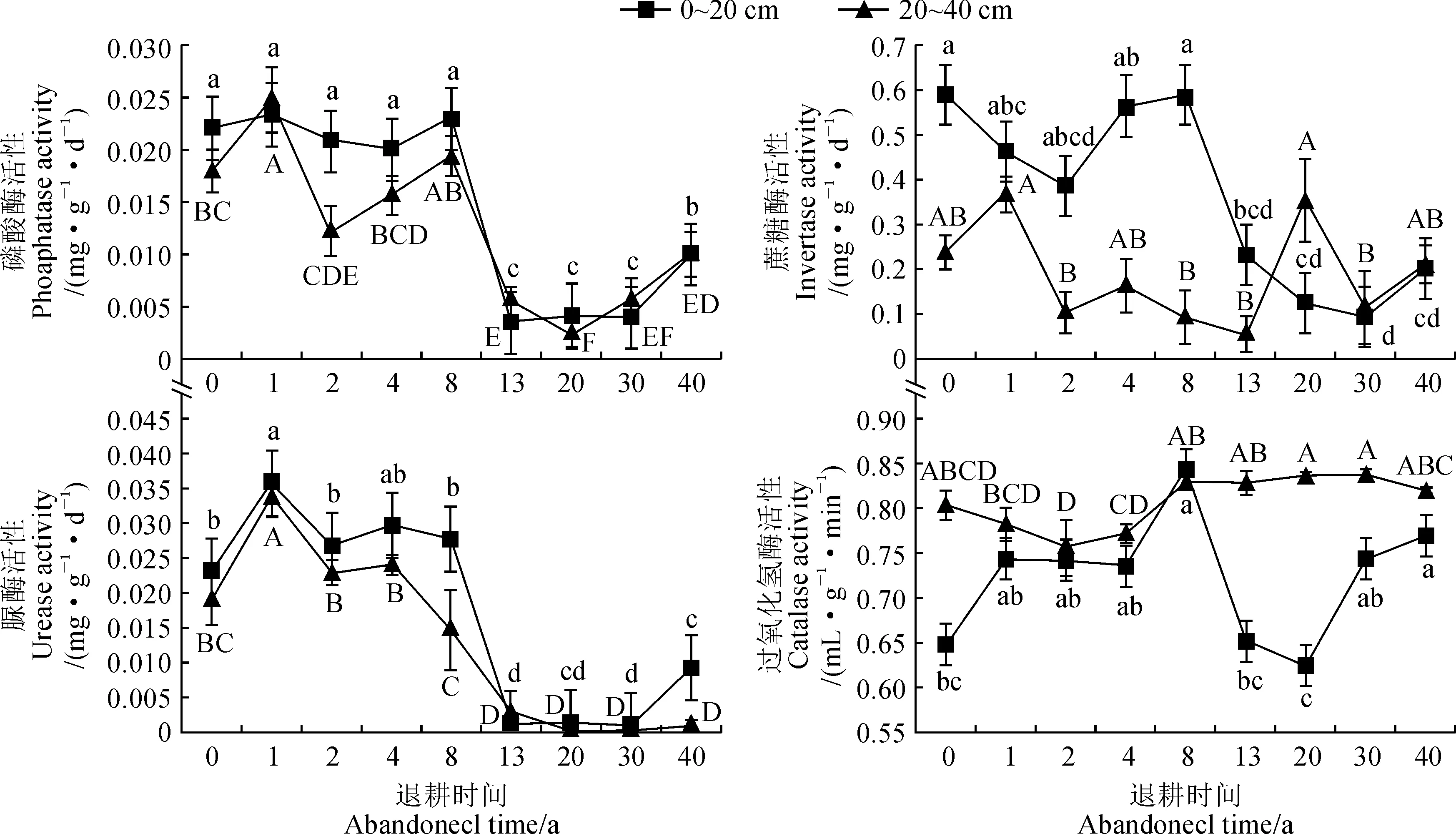
由图1可知,在0~20 cm土层中,土壤磷酸酶活性在退耕1 a时达到极大值0.023 mg·g-1·d-1,退耕13 a时达到极小值0.004 mg·g-1·d-1,在退耕8~13 a时波动幅度大,差异显著;在0 a时土壤蔗糖酶活性达到极大值0.592 mg·g-1·d-1,在退耕1~2 a时下降,4~8 a时上升后再迅速下降,在退耕30~40 a时回升,在退耕30 a达到极小值0.092 mg·g-1·d-1,且在不同年限退耕样地间差异性均不显著;土壤脲酶活性变化比较明显,在退耕0~1 a呈上升趋势,退耕1 a时达到极大值0.036 mg·g-1·d-1,在退耕1~2 a呈降低趋势,退耕2~8 a较为稳定,退耕8~13 a脲酶活性差异显著,波动幅度大,在退耕13~30 a时脲酶活性变化平稳,在退耕30~40 a时回升;过氧化氢酶活性总体呈先上升后下降再逐渐趋于稳定,退耕8 a时达到极大值0.842 mL·g-1·min-1,退耕20 a达到极小值0.624 mL·g-1·min-1,8~13 a和20~30 a过氧化氢酶活性波动幅度大,差异显著。在20~40 cm土层中,土壤磷酸酶活性呈先升高(0~1 a)后波动式下降(1~20 a)再升高(20~40 a)的趋势,退耕1 a时达到极大值0.0250 mg·g-1·d-1,退耕20 a达到极小值0.002 mg·g-1·d-1,退耕1~2 a和8~13 a土壤磷酸酶活性波动幅度大,差异显著。除退耕1 a、13 a、30 a和40 a以外,其他退耕年限均表现出土壤磷酸酶活性上土层高于下土层;土壤蔗糖酶活性在退耕1 a时达到极大值0.368 mg·g-1·d-1,退耕13 a时达到极小值0.053 mg·g-1·d-1,在退耕0~1 a时迅速上升,退耕1~2 a时急速下降,退耕2~13 a时呈波动式下降趋势,退耕13~20 a呈上升趋势,退耕20~30 a呈下降趋势,退耕30~40 a呈上升趋势,在退耕0~13 a中土壤蔗糖酶活性随土层加深而降低,在退耕20~40 a中随土层加深而升高;脲酶活性呈先上升后下降趋势,退耕1 a时达到极大值0.034 mg·g-1·d-1,30 a时达到极小值0.0003 mg·g-1·d-1,除13 a外,其他年限的土壤脲酶活性均随土层加深而降低;但土壤过氧化氢酶活性的变化趋势不显著,退耕2 a时达到极小值0.756 mL·g-1·min-1,退耕40 a时达到极大值0.819 mL·g-1·min-1,除退耕8 a外,其他年限的土壤过氧化氢酶活性均随土层加深而升高。
2.4 植物群落特征与土壤酶活性间的相关性
对不同深度的土壤酶活性和植物群落特征之间的关系进行相关性分析,由表4可知,0~20 cm土层中,过氧化氢酶活性与个体数呈显著负相关关系(P<0.05);物种数与土壤蔗糖酶活性、磷酸酶活性以及脲酶活性呈显著正相关关系(P<0.05);20~40cm土层中,物种数和土壤脲酶活性呈极显著正相关关系(P<0.01),而与过氧化氢酶活性呈显著负相关关系(P<0.05);Margalef丰富度指数与0~20 cm土层的磷酸酶活性和蔗糖酶活性均呈显著正相关关系(P<0.05),与脲酶活性呈极显著正相关关系(P<0.01);20~40 cm土层中,Margalef丰富度指数与脲酶活性呈极显著正相关关系(P<0.01),与过氧化氢酶活性呈显著负相关关系(P<0.05);Shannon多样性指数、Pielou均匀度指数和Simpson优势度与4种土壤酶活性间相关性较低且均不显著(P>0.05)。
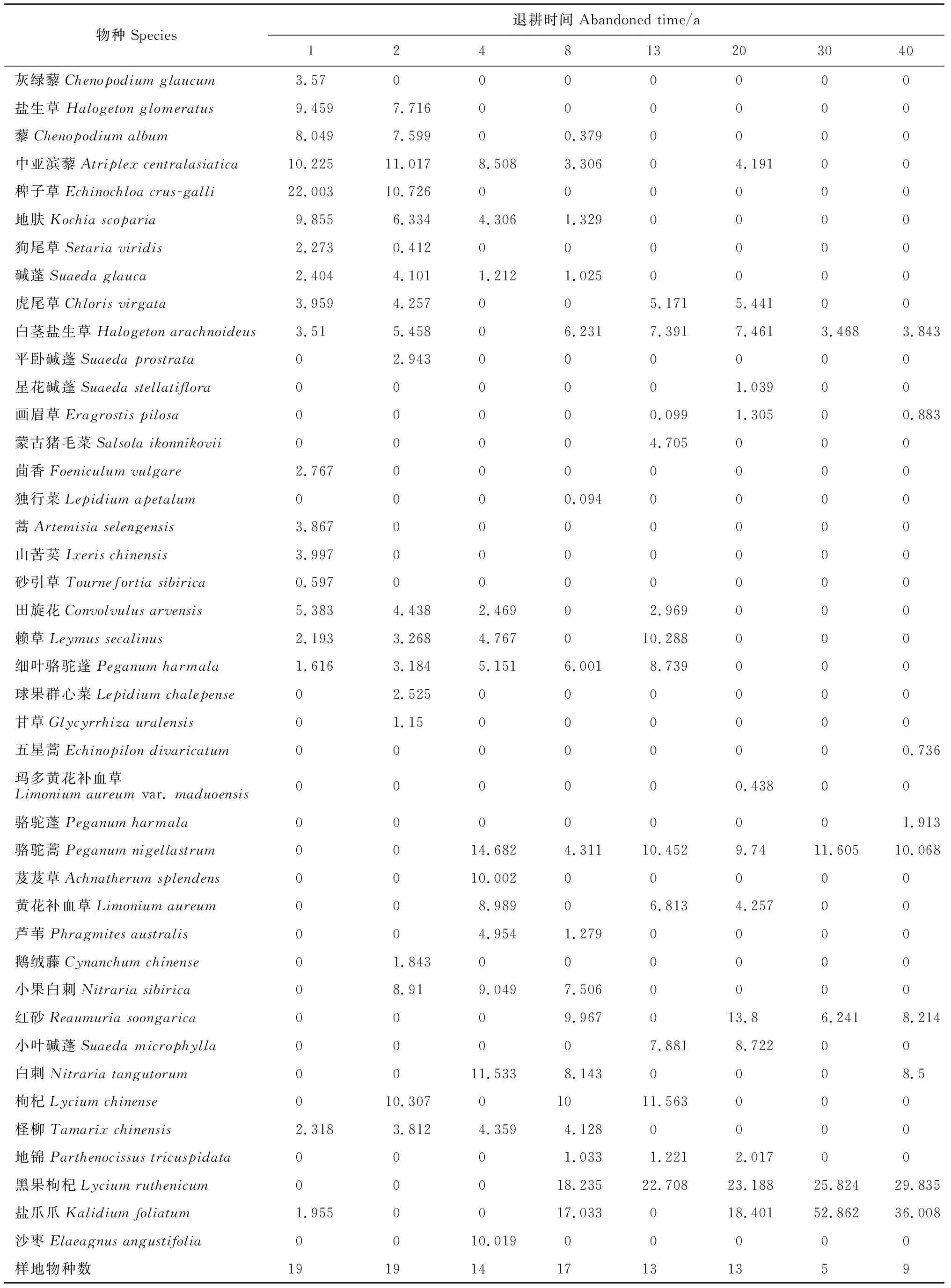
表2 不同退耕年限植被群落的物种组成和重要值
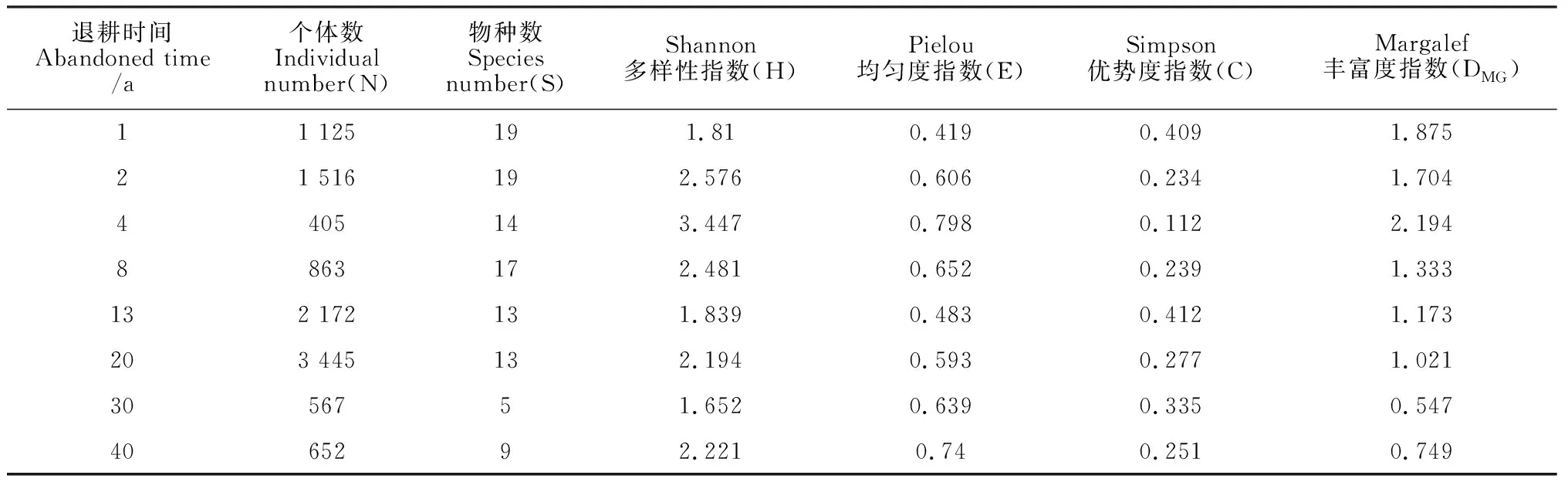
表3 物种多样性随退耕年限变化表
不同字母表示处理间差异达P<0.05图1 不同年限退耕地土壤酶活性变化Different letters above the bars mean significant difference at the 5% levelFig.1 Changes in soil enzyme activities of fallow land in different years
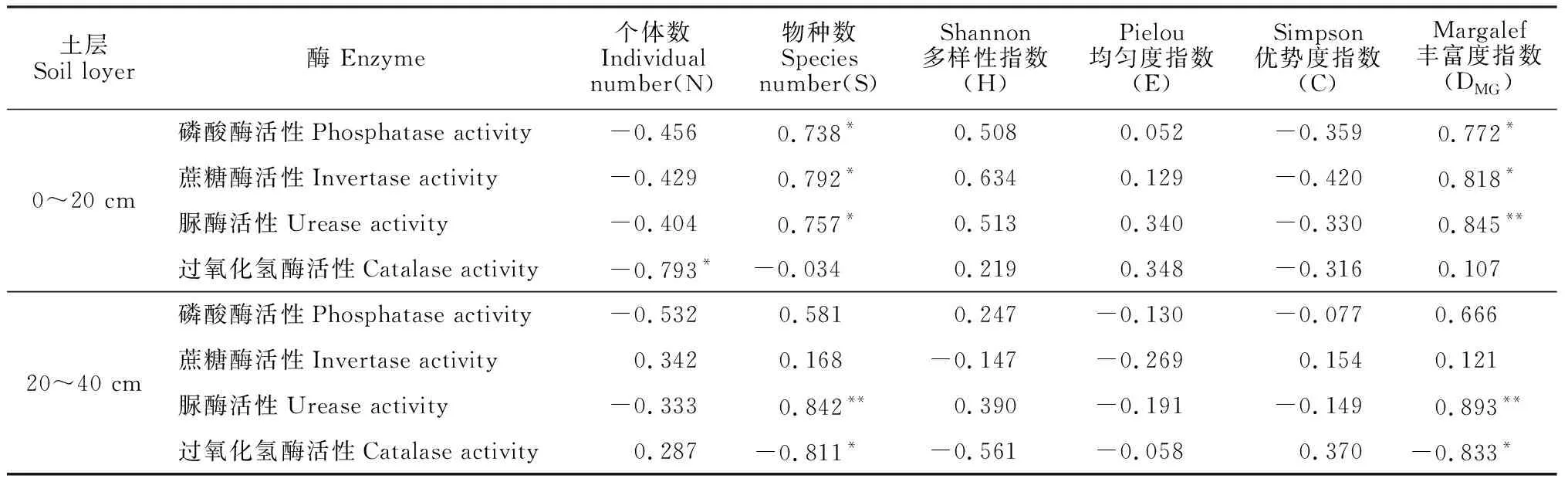
表4 植被群落多样性和土壤酶活性相关性表
3 讨 论
群落结构是植物群落的基本属性,也是认识群落的组成、变化和发展趋势的基础[40],群落物种多样性是一个群落结构和功能复杂性的量度[41],研究物种多样性是群落生态学的重要研究课题[42],利于阐明植物群落结构特征的发展阶段及稳定性的问题[43]。
从不同样地植被群落物种组成及重要值综合分析来看,在青土湖退耕40 a的植被演替过程中,9个样方共出现16科32属42种植物,其中,藜科、禾本科、蒺藜科、十字花科、菊科、柽柳科、茄科以上7科占总物种数的78.57%,比例比较大,说明这些植物在退耕地的植被演替中起着重要的作用,而且物种构成表现为多数种属于少数科、少数种属于多数科,大部分植物种为单属单科;植被演替过程中表现出连续性特征,如白茎盐生草基本在不同年限退耕地中连续出现。此外,不同年限退耕地植被群落演替还具有递进性。总体来说,随退耕年限增加,植物群落演替大致经历了地肤→盐生草→狗尾草→骆驼蒿→小叶碱蓬→黑果枸杞→红砂→盐爪爪的演替过程。符合荒漠地区植物区系的总体特征,这种结构和王理德[44]对民勤退耕区次生草地研究相一致。
青土湖40 a退耕地植被恢复过程中,随退耕年限增加,物种在逐渐减少并趋于简单。在退耕1~4 a中,Shannon多样性和Pielou均匀度指数不断增加,说明在退耕初期时,群落内物种的种类在逐渐增加,此时由于退耕前农作物的施肥,使得植被的生长空间和生长资源相对充足,一年生草本植被生长没有受到限制;在退耕4~13 a中逐渐减小,在退耕20 a后慢慢趋于稳定。当一年生草本植被数量达到土地承载上限后,植被的生长空间和生长资源不足,不能供所有植被生长,因此开始出现竞争,部分植被没有足够的生长空间和生长资源被淘汰,因此植被种类开始减少,最后趋于稳定。Simpson优势度指数的变化表明,植物群落物种在自然演替过程中向着简单、稳定的方向逐渐发展。退耕过程由草本植物向灌木演替,这一结论与李治元等[45]研究的石羊河下游盐渍化弃耕地植被演替特征分析的结果相似,但与白文娟等[46]研究黄土丘陵沟壑区退耕地、李永强等[47]研究典型草原区撂荒地的结果不同,这是可能是由于不同环境条件所导致的,本实验地为青土湖干旱区退耕地,与典型草原区撂荒地、黄土丘陵沟壑区退耕地的水文、土壤、地形、气候等不同,因此导致结果也不同。
土壤酶是土壤中不可缺少的成分,可反映土壤的环境健康状况[48],可作用于转化土壤的有机质[49]。在不同土层(0~20 cm和20~40 cm)的4种土壤酶活性随退耕年限增加表现出先升高后下降再逐渐趋于稳定,在退耕初期,随着植被的恢复,其种类和数量逐渐增加,植被的根系和枯落物为土壤微生物提供了良好的生存环境,土壤微生物数量种类也随之增加[50],而土壤酶多来源于土壤微生物和植被根系,与此同时植被枯落物和植物根系分解也会需要大量土壤酶进行催化,因此土壤酶活性逐渐升高。随后植被经过竞争演替阶段逐渐稳定下来,植被数量和种类也随之降低,其产生的枯落物以及土壤中的有机质也随之减少,因此土壤酶活性逐渐降低趋于稳定。本研究发现随退耕年限增加,土壤磷酸酶活性与土壤脲酶活性的变化显著,但土壤蔗糖酶活性与土壤过氧化氢酶活性变化均不显著,两者均于退耕1 a时达到最大值,这与韩福贵等[51]在石羊河下游退耕地的研究结果相似,但是与孟立君[52]研究的不同种植年限土壤酶活性特征、刘雨等[53]研究燕沟流域退耕地土壤酶活性特征的结果有差异。这是由于退耕初期,土壤中持有原来耕作时的大量水分和养分,致使退耕后,出现大量的杂草,以1 a生的草本植物为主,种类繁多,植物根系释放出较多的酶,土壤中有机质和其他因子含量较多,加之在土壤表层伴有大量的腐殖质及枯枝落叶等,使土壤中微生物数量增加,土壤物质代谢能力加强,致使土壤酶活性大幅提高,之后,土壤水分和养分随退耕年限增加而逐渐被消耗,部分植物死亡,极少数耐干旱的植被存活下来,植物种类的降低,其产生的枯枝落叶和腐殖质相应减少,微生物的数量也随之下降,土壤脲酶活性和磷酸酶活性均出现不同程度的下降,但随退耕年限的增加,土壤过氧化氢酶活性升降变化不显著,说明该区退耕年限的变化对土壤过氧化氢酶活性的影响不明显。
从土层垂直分布来看,土壤磷酸酶、蔗糖酶和脲酶活性均随土层深度的增加而下降,这与有关的研究报道一致[54],但与王学林等[55]在沙地樟子松人工林土壤酶活性的研究结果不同。这是因为研究区20~40 cm土壤层的腐殖质和枯枝落叶比0~20 cm土壤层的少,0~20 cm土壤层有充分的营养源,促进微生物大量的繁殖和生长,而且,土壤表层的水分条件,温度条件及通气状况较好,使微生物的生长速度加快,代谢更加活跃,导致土壤表层酶活性大。另外,由于该区光照充足,蒸发强烈,降雨稀少,天气干燥,属于典型的温带大陆性气候[56],若遇降雨也只能存留在土壤的表层,随土层不断的加深,水分和养分也随之降低,细根和凋落物的数量减少,抑制微生物生长,导致土壤酶活性随土层加深而降低,但是深层土壤的酶活性(蔗糖酶)出现了较高的现象,主要是因为在退耕13 a后,植被数量和种类因缺水等因素降低,出现了以黑果枸杞为主的单一群落,降低对土壤水分的消耗[57],在这一阶段,大多数的植被主要是以深根系的灌木植物为主,根系作为土壤与植物进行信息传递和物质交换的重要载体,是植物响应外界环境改变的重要途径,也是构成不同根际微生态特征的关键因素[58]。根系分泌物,腐殖质及枯枝落叶的增加,会直接作用于根际土壤,加速了土壤的养分积累,这会使根际土壤酶活性在一定程度上有所提高,从而改善土壤性质,提高土壤肥力[59]。而土壤肥力提高后,可以为该研究区的植被生长提供更丰富的营养物质。
土壤酶系主要来源于微生物的分泌、残体的腐解及动植物的分泌[60],土壤酶的含量可直接或间接地受植物的影响[61]。植物群落特征与土壤酶活性相关性分析表明物种数越多,土壤脲酶活性越强,可见土壤脲酶活性对植物群落特征的变化影响较大;通过0~40 cm土层Margalef丰富度指数与土壤脲酶活性的相关性分析表明:Margalef丰富度指数越大,土壤脲酶活性越高,Margalef丰富度指数对土壤脲酶活性影响较大。总体来看,Shannon多样性指数、Simpson优势度和Pielou均匀度指数与4种土壤酶活性影响较小;个体数、物种数、Margalef丰富度指数对4种土壤酶活性影响较大。这主要是植被在自然恢复的过程中,其覆盖度的增加,生长产生的枯枝落叶和腐殖质以及根系的分泌物也随之增加,大量枯枝落叶使土壤有机质和其他相关因子含量较高,提供了大量的酶促反应底物,最终影响土壤酶活性,使其发生变化。同样的,土壤酶活性的变化又会使植物的生长受到影响。植被覆盖率下降,土壤中留存的速效钾、土壤有机质等与之相关的理化因子含量下降,酶的转化和分解过程减弱,循环变慢,酶活性功能也随之减小。
4 结 论
1)青土湖退耕40 a自然恢复过程中,9个样方中共出现16科32属42种植物,物种构成表现为:多数种属于少数科,大部分植物种为单属单科。
2)随着退耕年限的增加,群落优势种由草本植物逐渐转化成灌木植物。
3)植物Margalef丰富度指数呈波动式下降的趋势;Shannon多样性指数和Pielou均匀度指数总体趋势为先增加后减小,再逐渐趋于稳定;Simpson优势度指数整体变化幅度较小,最终呈稳定的趋势。植物群落物种在自然演替过程中向着简单、稳定的方向逐渐发展。
4)在不同土层(0~20 cm和20~40 cm)中,4种土壤酶活性随退耕年限增加总体表现出先升高后下降再趋于稳定的趋势。均在退耕8 a后显著下降,且磷酸酶和脲酶活性均显著低于蔗糖酶活性和过氧化氢酶活性。与对照样地(CK)相比,磷酸酶活性与脲酶活性均随退耕年限增加而下降;土壤磷酸酶、蔗糖酶和脲酶活性均随土层加深而降低。
5)通过相关分析显示,在0~20 cm土层中,过氧化氢酶活性与个体数呈显著负相关关系;物种数与蔗糖酶活性、磷酸酶活性以及脲酶活性的关系显著相关,呈正相关关系;20~40 cm土层中,物种数与土壤脲酶活性呈极显著正相关关系,与土壤过氧化氢酶活性呈显著负相关关系;Margalef丰富度指数与土壤0~20 cm土层的磷酸酶活性和蔗糖酶活性均呈显著正相关关系,与脲酶活性呈极显著正相关关系;在20~40 cm土层中,Margalef丰富度指数与土壤脲酶活性呈极显著正相关关系;Shannon多样性指数、Pielou均匀度指数和Simpson优势度与4种土壤酶活性间相关性较低且均不显著。
6)青土湖退耕地植被自然演替过程中,物种多样性逐年降低,多样性指数总体趋于减少,土壤酶活性对特种多样性影响较大,特别是脲酶活性对物种多样性起着关键性作用。
