ABC转运蛋白在植物花器官生长发育中的研究进展
2022-01-15申慧敏史俐莎田清尹刘家伟王良桂岳远征
申慧敏,史俐莎,田清尹,刘家伟,王良桂,岳远征*
(1 南京林业大学 风景园林学院,南京 210037;2 江苏省风景园林重点实验室,南京 210037)
ABC转运蛋白(ATP-binding cassette transporter)广泛存在于生物体中,是目前已知最大、功能最多的转运蛋白家族之一,并在真核生物与原核生物的跨膜物质运输中发挥重要的作用[1-2]。最初对于该基因家族的研究始于医学领域,近年来在动物、植物以及微生物中都对该基因家族开展了广泛的研究[3]。现有研究表明该家族成员几乎参与了植物生长发育各阶段的物质运输,例如植物激素运输、重金属离子外排、植物应对环境胁迫、次生代谢产物的运输等。此外,该基因家族成员的转运底物较为多样,不仅包括植物体内激素、金属离子、糖类、脂质、次生代谢产物和外源物质,而且还包括生物碱、糖、肽、无机离子以及类固醇等物质[4-7]。近期,在许多单子叶和双子叶植物中已经进行了该基因家族的鉴定,并对部分基因进行了深入的功能研究[8-11]。目前ABC家族成员在增强植物抗性以及调节植物生长物质运输等方面的研究已经比较系统,在植物花器官中对于ABC家族成员的研究也已经有较多的报道,但是还未有ABC转运蛋白影响花器官生长发育方面的综述。基于目前ABC转运蛋白的研究现状,本研究总结了该蛋白家族的结构特点和亚家族分类情况,并主要从ABC转运蛋白家族对于植物花器官观赏性状的影响方面进行概括,以期能为植物观赏性状的改良提供参考。
1 ABC转运蛋白的结构特点
ABC转运蛋白又称ATP结合盒转运蛋白,因其含有核苷酸结合域而得名。ABC转运蛋白具有两种特征结构域:核苷酸结合域(nucleotide-binding domains,NBD)和跨膜结构域(transmembrane domains,TMD),依据这两种特征结构域的不同数目,ABC转运蛋白被分为全分子转运蛋白、半分子转运蛋白以及可溶性蛋白[12]。其中,全分子转运蛋白具有两个核苷酸结合域和两个跨膜结构域,半分子转运蛋白仅含有一个核苷酸结合域和一个跨膜结构域,而可溶性蛋白则只含有核苷酸结合域[13]。全分子转运蛋白是进行物质运输的最小功能单位,半分子转运蛋白必须要形成同源或者异源二聚体才能进行物质运输,而可溶性蛋白则不具备物质运输功能。
NBD和TMD这两种特征结构域的结构、功能都有所差别。NBD的长度约有200个氨基酸,具有ATP酶结合位点,该结构域序列较为保守,包括结合盒A(P-loop)基序以及结合盒B基序、D环、H环、ABC特征序列(C基序)和Q环(图1)[13-15]。保守序列基序Walker A和Walker B由C基序分隔开,其中Walker A结构域和Walker B结构域在所有ATP结合蛋白中都存在,而C基序则是ABC转运蛋白所特有的,可用于区别ABC转运蛋白和其他ATP酶[16]。其中,NBD作为ABC蛋白最典型的结构域,结构相对保守,在不同物种中具有30%~40%的同源性;而TMD的序列则并不保守,其结构高度多样化,在初级序列、长度、结构和跨膜螺旋的数量上存在很大差异[15-16]。此外,NBD和TMD在物质运输中也具有明确的分工,NBD可以通过水解ATP为物质的转运提供能量;而TMD则负责识别底物,由两个TMD通过结构变化组成物质跨膜转运的通道,使底物通过磷脂双分子层进而完成物质运输的过程[17-20]。
2 ABC转运蛋白家族成员的鉴定
近年来伴随着基因组测序技术的不断发展,越来越多的基因家族已被鉴定出来,利用生物信息学筛选的方法可以更加方便快捷地挖掘到需要的基因,并为进一步的基因功能研究奠定基础。自从最早在拟南芥中发现了第一个ABC转运蛋白基因之后,在其他植物中也陆续展开了该基因家族的研究,目前在单子叶植物玉米(Zeamays)、水稻(Oryzasativa)以及双子叶植物拟南芥(Arabidopsisthaliana)、辣椒(Capsicumannuum)、西红柿(Solanumlycopersicum)、菠萝(Ananascomosus)、葡萄(Vitisvinifera)、月季(Rosachinensis)等植物中已经进行了该基因家族的鉴定[8-11]。ABC家族成员在植物中的鉴定结果表明,该家族成员在不同的植物中数目有较大的差别(表1),大部分植物中ABC家族成员数目在100~200的范围之内,其中在甘蓝型油菜(Brassicanapus)中该基因家族成员最多,达到314个,而在喜阴悬钩子(Rubusoccidentalis)中则数目最少,仅有98个成员[26-27,29];同时,ABC转运蛋白家族中不同亚家族成员的数目也存在一定的差异,如该家族在植物中共有8个亚家族,ABCB,ABCC,ABCG为成员数目较多的3个亚家族,其中ABCG亚家族成员数目则往往最多[24]。

图1 ABC转运蛋白中核苷酸结合域(NBD)结构示意图Fig.1 Schematic diagram of NBD structural structure

表1 不同植物中ABC家族成员的数目
3 ABC基因家族的分类及功能概述
ABC转运蛋白家族庞大,家族成员较多。目前应用较广泛的分类方法是人类基因组织命名法,该命名法规定ABC家族共有9个亚家族(ABCA-ABCI),其中有8个亚家族(ABCA-ABCG和ABCI)存在于植物中[29]。ABC转运蛋白在植物中的8个亚家族成员之间的结构组成和主要生物学功能也存在一定的差异(表2)。
ABCA亚家族成员既包括全分子转运蛋白又包括了半分子转运蛋白,其中全分子转运蛋白又被命名为AOH,半分子转运蛋白则被命名为ATH,全分子类转运蛋白的功能可能与甾醇的运输有关,而半分子类的转运蛋白的功能还不清楚;ABCB亚家族同样也包含全分子转运蛋白和半分子转运蛋白,全分子转运蛋白又称MDR型转运蛋白,半分子转运蛋白则包括TAP、ATM和LLP三类,全分子类ABCB蛋白的主要功能是解毒和运输生长素,半分子类ABCB转运蛋白则大多参与响应重金属胁迫;ABCC亚家族成员基本都是全分子转运蛋白,又称为MRP相关蛋白,它们与运输离子进入液泡有关;ABCD亚家族蛋白通常是半分子PMP(perox异构膜蛋白)转运体,但是在拟南芥和葡萄中也发现了全分子的ABCD蛋白,它们与脂肪酸进入过氧化物酶体的运输有关;ABCE和ABCF这两个家族的成员在生物进化过程中都较为保守,在植物中的成员数目及相关功能研究也较少,ABCE与核糖体相关生物转化有关,被国际生物医学研究组织作为一种核糖核酸酶抑制剂(RNase L,RLI)[30-31],而ABCF亚家族统称为GCN,同时该家族成员与植物的抗性相关[32]。ABCG亚家族有全分子和半分子转运蛋白,全分子成员也称为PDR转运体,与重金属胁迫以及生长素的转运相关[33-34],WBC转运蛋白是ABCG亚家族中的半分子成员,可用于角质层脂类和烷烃的运输[35-37],拟南芥中已经报道了3个ABCG转运体可以有效地运输角质层的脂质,进而促进角质层的发育[38-40]。ABCI转运蛋白家族具有单一的NBD结构域,一些研究表明该家族成员与细胞色素C成熟复合物密切相关[30-31]。

表2 ABC转运蛋白各亚家族的结构域组成以及功能概述
4 ABC转运蛋白对植物花器官的影响
花是植物的重要观赏部位,优美的花型、鲜艳的花色以及花自身所散发的香味都是植物重要的观赏特性,同时植物的生殖繁育也离不开花器官的正常发育。近年来越来越多的ABC转运蛋白在植物花器官中有了功能报道,其中在花色、花香、花瓣形态这些观赏性状以及花药、花序等花器官生长发育方面均有涉及,本研究就ABC转运蛋白在不同植物花器官中的功能进行了总结(表3)。
4.1 ABC转运蛋白对植物花瓣形态的影响
花瓣形态是观赏植物的重要观赏特性之一,现有研究表明ABC家族的成员因运输花瓣表皮细胞角质层形成所需的脂类物质可以影响花瓣的正常伸长,导致产生折叠的花瓣进而改变花瓣的形态。目前发现拟南芥中AtABCG11、AtABCG12以及AtABCG13都对花瓣形态有影响,拟南芥花瓣角质层主要成分是链内羟基脂肪酸,这3个ABCG类转运蛋白都在花瓣角质层中发挥作用,协同作用于角质层的形成[39-40]。在这3个基因中AtABCG11运输的底物最为广泛,该基因的缺失不仅会导致花瓣折叠,同时还会导致器官的融合以及发育不良;AtABCG11不仅可以形成同源二聚体,也可以和AtABCG12共同形成异源二聚体来发挥功能。同时,AtABCG13的缺失会导致出现与AtABCG11突变体类似的折叠型花瓣,研究表明这两个基因可能在同一途径的不同阶段调节花瓣伸长,即AtABCG11参与脂肪酸的产生途径,而AtABCG13则会转运AtABCG11生成的产物;另外,AtABCG13还与AtABCG11、AtABCG12和AtABCG15 这3个基因共同表达,均具有蜡质转运的功能[41]。
在蒺藜苜蓿(Medicagotruncatula)中,MtABCG13突变体可通过调节花瓣和雄蕊表面角质层的分泌,影响花瓣和雄蕊的伸长,导致柱头外露。野生型蒺藜苜蓿的花瓣在伸长时可以平滑地穿过萼片和花药,而MtABCG13突变体的花瓣伸长时则由于花药和萼片的接触而受阻,使花瓣不能正常伸展进而发育成折叠的花瓣,由于折叠型花瓣占据了大部分空间就会限制雄蕊管的正常伸长,因此产生柱头外露的现象[42]。但柱头外露的现象目前只在豆科植物中发现,这可能也与豆科植物独特的花瓣结构有关,在其他植物中还有待探究,同时柱头外露使得只能自花授粉的植物可以进行异花授粉,对于自花授粉植物杂交育种具有重大意义。
4.2 ABC转运蛋白对植物花色的影响
花色也是植物较为重要的观赏特性之一,植物体内的色素主要包含类黄酮、类胡萝卜素以及生物碱类色素3类。其中,类黄酮是植物体内分布最广泛的一类色素,可以使植物的花色呈现由浅粉到紫色的观赏特性,类黄酮类主要包括花青苷和辅色素,花青苷的积累很大程度上影响植物的颜色表型[43]。
花青苷的运输也影响植物的花色,植物体内存在高效的花青苷转运机制,可以将细胞质内合成的花青苷运输到液泡内。花青苷分子运输到液泡中包括以膜为介质、以囊泡为介质和以转运子为介质的3种方式。已有研究表明ABC转运蛋白家族中的MRP/ABCC亚家族参与到花青苷以膜为介质的转运途径中,研究认为位于液泡膜上的MRP亚家族成员可以形成一种谷胱甘肽交联复合物,识别花青苷并将其跨膜转运至液泡内,这一转运机制在玉米、拟南芥、水稻、葡萄中均有报道。例如,玉米中的ZmMRP3、拟南芥中的AtMRP1和AtMRP2、水稻中的OsMRP15以及葡萄中的VvABCC1都被认为是定位于液泡膜上负责向液泡转运花青素的转运体;此外,ZmMrp3、ZmMrp5、ZmMrp6和ZmMrp7在所有合成花青素的组织中都有表达[44-46],这也暗示着它们也有可能会对花色有一定的影响。
4.3 ABC转运蛋白对植物花香的影响
植物花瓣是花香物质释放的主要场所,释放的香气不仅可以增加花朵的观赏价值,而且还可以吸引传粉者,此外,有的花香物质还具有杀灭有害细菌、帮助植物进行防御的功能。 植物释放的花香物质主要分为3类,分别是萜类化合物、苯类/苯丙素类化合物以及脂肪酸类化合物,其中萜类化合物和苯类/苯丙素类化合物的研究相对较多[47]。
目前关于植物花香方面的研究主要集中在花香物质合成这一过程中,已经报道了较多的可以调控花香物质合成的功能基因和转录因子,但是关于花香物质从花瓣细胞中释放到外界的分子机制却鲜有报道。早期的观点认为是挥发性有机化合物通过扩散作用释放到空气中进而产生了香气,直到矮牵牛(Petuniahybrida)中PhABCG1基因的发现,才证明了挥发性有机化合物也可以通过主动运输的形式运到植物体外。PhABCG1基因在花瓣中特异表达,可以将矮牵牛花瓣内合成的挥发性苯类/苯丙素类化合物通过主动运输转运到花瓣表皮细胞外[48],通过降低矮牵牛花瓣中PhABCG1的表达量,发现在花瓣中各挥发性有机化合物的挥发量降低,而相应的细胞内部的挥发性有机化合物的含量则明显增加,以上研究表明了ABC家族成员可以促进矮牵牛花香物质中挥发性有机化合物的释放。
4.4 ABC转运蛋白对花粉发育的影响
植物的正常繁殖离不开雌雄配子的结合,只有雌雄配子能够正常发育,才可以结合形成正常的合子,使得植物可以正常的繁殖。当花药或者胚珠发育不正常时就会造成植物的不育。目前在拟南芥、水稻中的研究表明,ABC家族的成员在花粉的生长发育过程具有重要的作用。
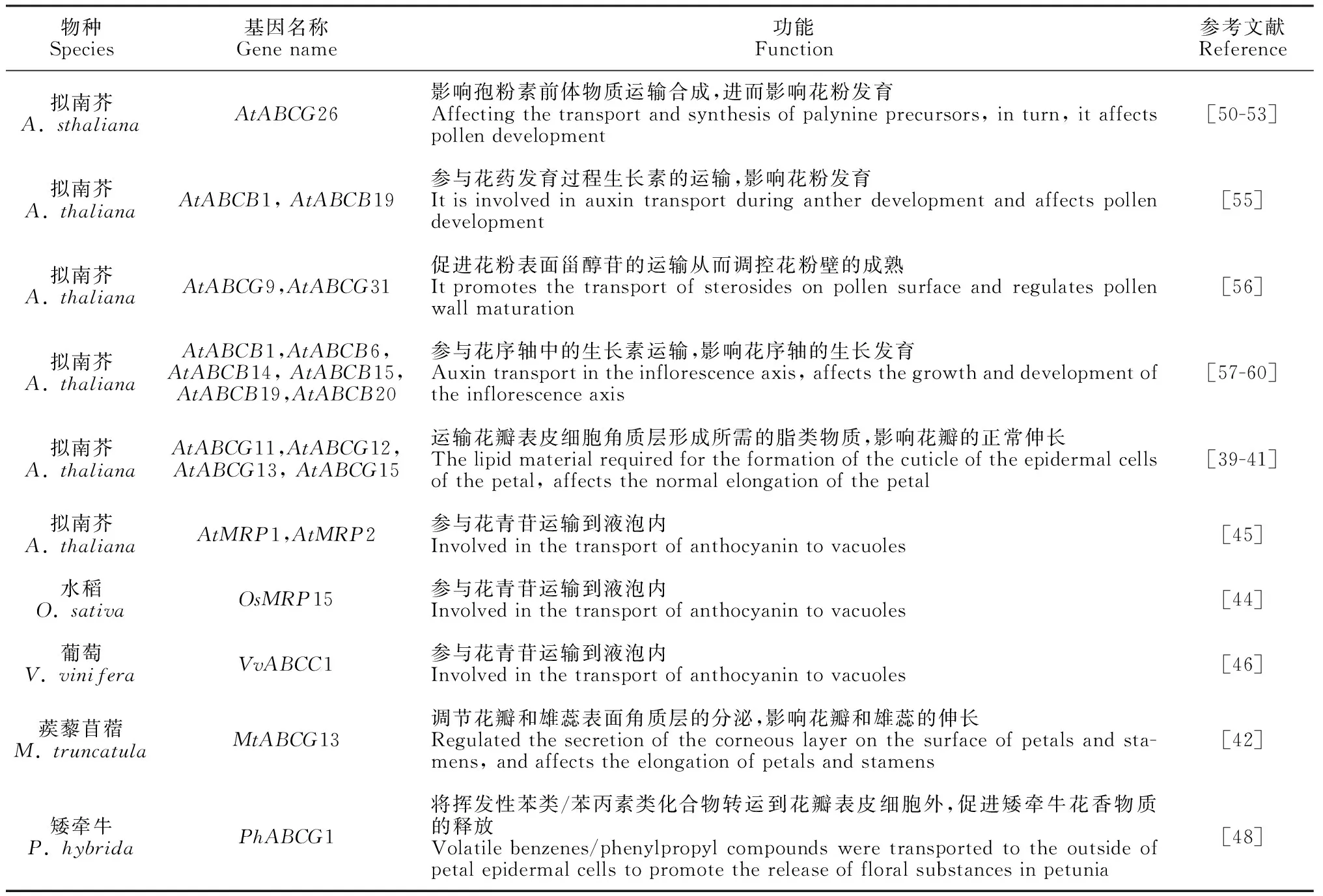
表3 在花器官中有功能报道的ABC家族成员
植物的花粉粒细胞具有完整的细胞壁和质膜,其中典型的花粉壁又包括花粉外壁、花粉内壁和花粉外被3层,每1层的组成成分也有所不同,花粉外壁的主要成分是孢粉素,而花粉内壁的主要成分则是纤维素,外被层则是一些脂质、蛋白质、色素和芳香族化合物[49]。研究发现多个ABC基因家族成员在双子叶植物拟南芥中参与到花粉发育过程中,如AtABCG26、AtABCG1、AtABCG16、AtABCB1、AtABCB19、AtABCG9以及AtABCG31。其中,AtABCG26与孢粉素合成所需的基因共同表达,并在花芽早期高度表达,同时应用反向遗传学的方法也证明了AtABCG26基因的缺失会影响花粉壁的正常发育,导致正常花粉形成受阻,使得不能形成正常的花粉最终造成了拟南芥的雄性不育;光学显微镜和透射电镜观察花粉结构发现,AtABCG26基因会影响孢粉素前体物质从绒毡层到花粉表面的运输,导致孢粉素不能正常合成,因此AtABCG26是花粉外壁形成和花粉成熟所必需的[50-53];同时,拟南芥中AtABCG1和AtABCG16这2个基因在花粉减数分裂后期具有重要的作用,它们的表达异常也会导致花粉不能正常发育[54]。除此之外,AtABCB1与AtABCB19也都参与到了花药的发育过程中,作为生长素转运体,它们在雄蕊发育早期、绒毡层发育和减数分裂细胞中都发挥着重要作用[55],其中的AtABCB1起主要作用,而AtABCB19则起协同作用。另外,ABC家族成员也会影响花粉的活力,例如在花粉发育后期表达的AtABCG9和AtABCG31,可以促进花粉表面甾醇苷的运输从而可以调控花粉壁的成熟,AtABCG9和AtABCG31的缺失则会使得花粉失去活力[56]。
4.5 ABC转运蛋白对花序生长发育的影响
花序是由许多花在花序轴上有规律的排列形成的,花序的存在影响着植物花朵的着生方式,而花序的生长发育过程离不开生长素的极性运输。生长素极性运输和积累是决定维管分化的重要因素,对细胞次生细胞壁的发育起着至关重要的作用,已有研究表明ABC转运蛋白中的多个ABCB亚家族成员参与了生长素的转运过程[57]。
在拟南芥研究中发现,AtABCB1、AtABCB6、AtABCB14、AtABCB15以及AtABCB20均参与了花序轴中的生长素运输,进而影响了花序轴的生长发育[59]。其中,AtABCB1是在拟南芥中发现的第一个ABCB成员,它在花序轴中的表达量非常高,通过超量表达以及RNAi抑制法均证明了该基因参与花序轴中生长素的运输[57]。同时,研究还发现将AtABCB1的同源基因AtABCB19突变后,会导致拟南芥花序轴以及下胚轴中生长素极性运输下降,进而导致花序轴向地性增强,影响花序轴的形态[59]。还有研究表明AtABCB6、AtABCB20、AtABCB15、AtABCB14在花序轴的生长素向基部运输过程中发挥着重要作用[61],其中AtABCB14与AtABCB15的突变可导致花序轴中生长素含量下降。而AtABCB14通过运输花序轴中的生长素进而影响花序轴维管发育和气孔开闭,其中AtABCB14缺失的植株花序轴中的维管束不能正常发育,暗示着AtABCB14可能通过参与拟南芥花序轴的木质化进而影响正常花序的形成[60]。
5 总结与展望
对于园林植物而言观赏性状的改良一直是其研究的重点之一,只有不断获得一些观赏性状优良的新品种,园林植物才能在园林绿化中更好地发挥作用。目前,研究表明ABC转运蛋白对植物花色、花香以及花瓣形态等观赏性状均具有显著的影响。因此研究ABC转运蛋白在植物花器官生长发育中的功能对于改良植物观赏性状具有重要的意义,例如可以尝试通过超量表达与花香转运相关的ABC家族基因进而增强观赏植物花瓣的香气释放速度,通过对转运花色苷相关的ABC家族成员进行沉默或者超量表达可以减轻或者增强植物的花色调控,同时还可以利用ABC家族成员尝试植物折叠型花瓣的创制,进而在某些观赏植物中创造独特的花型。
目前,ABC转运蛋白在植物花器官中的研究大部分还都集中在拟南芥、矮牵牛等模式植物中,在其他植物中研究较少,虽然在许多植物中已对该基因家族进行了全基因组家族成员的鉴定,但是关于其在植物观赏性状方面功能的研究仍相对较少。因此,应加强植物观赏性状方面的研究,如在花色、花香以及花瓣形态方面的研究。近年来随着生命科学技术的发展,越来越多的ABC蛋白家族成员的功能将会被报道出来,期望将来ABC蛋白家族成员会在植物观赏性状改良中发挥越来越重要的作用。
