杧果GeBP转录因子家族基因的鉴定及表达分析
2022-01-14孙瑞青刘志鑫夏煜琪高庆远蒲金基党志国2b
孙瑞青,杨 楠,刘志鑫,夏煜琪,孙 宇,高庆远,蒲金基,党志国,张 贺,2b
(1.海南大学 热带作物学院,海南 海口 570228;2.中国热带农业科学院 a.环境与植物保护研究所;b.农业农村部热带作物有害生物综合治理重点实验室;c.热带生物技术研究所;d.热带作物品种资源研究所,海南 海口 571101)
植物表皮毛(trichome)是大多数陆地植物表皮组织特有的一种结构,其种类多样[1],通常可分腺性毛状体和非腺性毛状体两种类型[2]。表皮毛能在一定程度上增加植物表皮的厚度,减轻昆虫、病原体对植物造成的损伤,防止热量、水分的散失,从而起到保护植物正常生长发育的作用[3-4],但是,不同植物表皮毛在不同环境中发挥的作用有所不同[5-6]。左香君等[7]发现,野生甘蓝Brassica incana的表皮毛在一定程度上对蚜虫具有抗性;宋海慧[8]认为,毛状体丰富的番茄Lycopersicon esculentum更加耐冷、抗旱,这说明植物表皮毛对生物及非生物均有抵御作用。
在拟南芥Arabidopsis thaliana中,无表皮毛增强子(Glabrous-enbancer binding protein, GeBP)可通过调控GLABROUS1(GL1)基因的表达来控制表皮毛的发生[9-10]。GeBP是一种核蛋白,其与黄色荧光蛋白(YFP)的共定位结果表明,该基因的表达仅限于3~5个亚核中,有一个中央结合的DNA结构域[11],C端区的主要特征是存在一种假定的亮氨酸拉链,表明GeBP中心区域可能形成同二聚体[12]。随后,GeBP转录因子家族在黄瓜Cucumis sativus[13]、水稻Oryza sativa[14]、毛竹Phyllostachys edulis[15]等种植物中均被发现,其均参与了植物表皮毛的形成。单雪萌等[15]对毛竹P.edulis的GeBP进行了qRT-PCR分析,结果表明,在16个GeBP家族成员中,有12个在带有表皮毛的叶、箨、箨片和纤毛中的表达量均高于其在没有表皮毛的笋中的表达量;拟南芥A.thaliana的AtGeBP/GPL基因在某些特定的组织中表达,对细胞分裂素信号作出响应[16],调控AtCPR5信号途径中的反应和细胞壁代谢相关基因的表达[11],但二者对细胞的增大存在着拮抗作用[17]。为给杧果种质资源的抗病性鉴定提供参考依据,鉴于植物表皮毛在其防御病害过程中的作用,对与杧果Mangifera indica表皮毛形成相关的GeBP转录因子家族的功能展开研究,对其MiGeBPs基因家族成员进行生物信息学分析,并利用qRT-PCR方法对MiGeBPs在杧果组织中及其在杧果受杧果胶孢炭疽菌和细菌性黑斑病菌侵染时的相对表达量进行分析,以期为杧果GeBP家族作用与功能的研究奠定基础。
1 材料与方法
1.1 杧果病害的胁迫处理及组织取样
从位于海南省儋州市宝岛新村的农业农村部儋州杧果种质资源圃(东经109.49°,北纬19.50°)中选取供试的杧果嫁接苗,选取‘贵妃杧’为供试的杧果品种,其砧木品种为‘海南土杧’。将树龄为1 a、叶龄为7 d、株高一致的杧果嫁接苗置于室温为25 ℃、相对湿度为70%~90%、光照12 h/黑暗12 h(即在室内黑暗与光照各12 h)的条件下以促进杧果嫁接苗的生长发育。
对杧果嫁接苗分别进行胶孢炭疽菌Colletotrichum gloeosporioides(以下缩写为“Cg”)和细菌性黑斑病菌Xanthomonas campestrispv.mangiferaeindicae(以下缩写“Xcm”)两种病害的接种处理。分别用含量为1 mL的2×106个分生孢子的胶孢炭疽菌分生孢子悬浮液、含量为2×107CFU的细菌性黑斑病菌悬浮液对杧果嫁接苗叶片进行均匀的喷雾侵染处理,处理时间分别设为0、3、6、12、24、48、72 h,其中的0 h处理组为对照组。每个处理组各取3株杧果嫁接苗的叶片样品,剪碎后置于液氮中快速冷冻并保存于-80 ℃的温度条件下以备用。
从位于海南省东方市的岛西林场(东经108.67°,北纬19.11°)选择研究所需的‘贵妃杧’成年果树,砧木为‘海南土杧’,将嫁接后新梢上抽出新叶的时间记为0 d,取其幼叶(7 d,即叶片的形态初形成)、老叶(35 d,即叶子已完全老化、革质化,用作对照)、芽、盛花、幼果(7 d)、中果(21 d)、大果(35 d)和成熟果作为供试材料。将所采集的试材置于液氮中快速冷冻并保存于-80 ℃的温度条件下,以备于杧果GeBP基因家族的组织特异性相对表达量分析之用。
1.2 RNA的提取及cDNA的合成
按照天根生化科技有限公司RNAprep Pure多糖多酚植物总RNA提取试剂盒的说明方法对杧果叶片进行总RNA的提取,通过超微量紫外分光光度计(Nanodrop 2000C型)对总RNA的浓度进行测定,并保存于-80 ℃的温度条件下以备用[18]。第一链cDNA由RevertAid First Strand cDNA Synthesis Kit试剂盒反转录而成。
1.3 杧果全基因组数据的来源
杧果全基因组数据来源于NCBI(PRJNA487154)数据库,拟南芥A.thaliana、番茄L.esculentum、水稻O.sativa这3个物种的GeBP序列均下载自PlantTFDB(http://planttfdb.gao-lab.org/)数据库。
1.4 杧果GeBP基因家族成员的鉴定
AtGeBPs氨基酸序列从拟南芥数据库和PlantTFDB数据库中获得,以此作为查询对象,使用Blastp在线程序在杧果基因组数据库中进行比较,将E值调整为10-5。利用Pfam(http://pfam.xfam.org/)在线工具获取杧果蛋白序列结构域,最终获得所有杧果GeBP基因家族成员[19]。其他物种的GeBP基因家族成员直接下载于PlantTFDB数据库。
1.5 杧果GeBP基因家族的生物信息学分析
利用ExPASy - ProtParam tool在线工具对杧果GeBP转录因子家族成员的内含子、分子量、理论等电点、基因长度等理化性质进行预测[20],利 用SOPMA(https://www.expasy.org/)、NCBI Conserved Domain Search、SWISS-MODEL(https://swissmodel.expasy.org/)等在线工具对其蛋白质的二级结构、基因的保守结构域、蛋白质的三级结构分别进行预测;使用MEME在线工具(http://meme-suite.org/tools/meme)[21-22]分析杧果基因家族的蛋白质保守基序。
1.6 系统进化树的构建
利用ClustalW程序,将存在于杧果和其他3个物种的GeBP基因家族成员的氨基酸序列进行对比。采用MEGA 7.0软件,使用邻近法,以获得的GeBP蛋白序列构建其系统进化树,将校验参数Bootstrap值调整为1 000次重复[23-24]。
1.7 荧光定量PCR分析
采用引物设计软件Primer 5设计PCR特异扩增引物[25-26],构建18 μL的反应体系,每个处理各设3次重复,使用UltraSYBR Mixture 试剂盒(北京康为世纪),利用Quant Studio 6 Flex软件进行实时荧光定量PCR检测。采用的qRT-PCR反应程序为:10 min、95 ℃预变性,15 s、95 ℃变性,1 min、60 ℃退火/延伸,45个循环;在60 ℃时进行荧光信号的收集[19]。根据MiGeBPs的CDS序列,设计实时荧光定量PCR检测所用的引物,以杧果MiActin1(Actin)作为内参基因[27],引物序列见表1。利用实时荧光定量PCR仪软件(Quant StudioTM 6 Flex)分析获得各个样品的Ct值,以0 h的表达量作为对照,采用2-ΔΔCt法进行数据统计,以确定MiGeBPs的相对表达量[28],并在此基础上使用HeatMap 2.0软件绘制MiGeBPs相对表达量的热图。
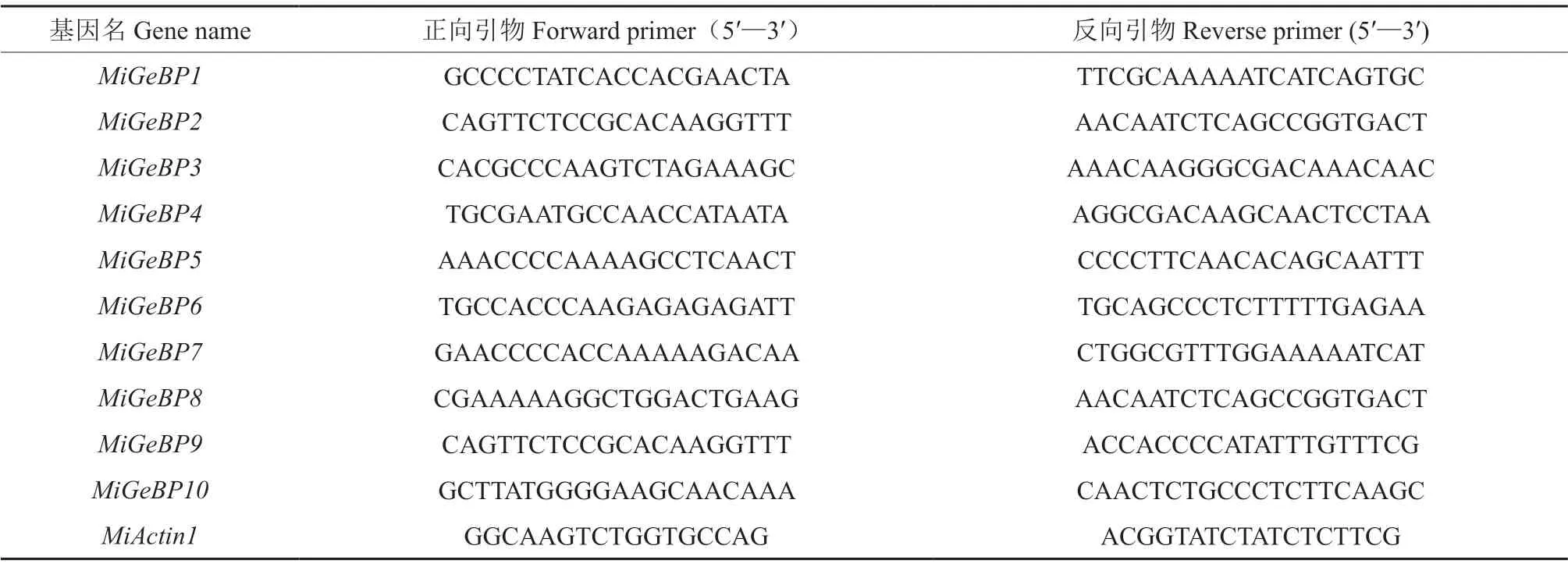
表1 荧光定量PCR检测所用引物序列Table 1 Primer sequences for RT-qPCR
2 结果与分析
2.1 杧果GeBP基因家族成员的鉴定结果
对杧果GeBP基因家族成员理化性质的分析结果见表2。表2显示,10个基因家族成员其理化性质的差异明显。10个基因家族成员的氨基酸个数差异较大;MiGeBP3的氨基酸数最多,达到527个;MiGeBP8的氨基酸数最少,仅有156个。其内含子数量,仅有MiGeBP3的为2个,其余9个基因的内含子数均为1个。10个基因家族成员的蛋白质分子量为18 020.02~58 488.64 Da,最大的约为最小的3倍。MiGeBP1、MiGeBP3、MiGeBP4、MiGeBP5、MiGeBP6、MiGeBP7的 等电点PI值均小于7,说明这6个蛋白均属于酸性;而MiGeBP2、MiGeBP8、MiGeBP9和MiGeBP10的等电点PI值均大于7,说明这4个蛋白均属碱性。其亲水性平均系数均为负值。其脂溶性系数范围为54.94~82.50。10个基因家族成员的不稳定指数为34.53~68.76;其中,MiGeBP6的不稳定指数最大,为68.76;MiGeBP2的不稳定指数最小,为34.53。由此可推测,MiGeBP基因家族成员均为不稳定的亲水蛋白。

表2 杧果GeBP基因家族成员基本理化性质的分析结果Table 2 Physical and chemical property of GeBP in M.indica
2.2 杧果GeBP基因家族蛋白质二级结构的预测结果
利用SPOMA在线工具预测到的10个MiGeBPs蛋白质的二级结构如图1所示。其中,无规则卷曲和α-螺旋均为MiGeBPs蛋白二级结构的主要元件,此二者的占比之和均大于85%;但是,在MiGeBP2和MiGeBP6蛋白的二级结构中,无规则曲和α-螺旋的占比相差较大。延伸链和β-转角在MiGeBPs蛋白结构中的占比均小于15%;其中,延伸链的占比,MiGeBP5、MiGeBP7、MiGeBP10的均仅在5%以下;β-转角,该基因家族各个基因的占比均很小,其在MiGeBP4与MiGeBP5中的占比仅分别为1.88%和1.57%

图1 MiGeBPs蛋白二级结构的在线预测结果Fig.1 On line prediction results of MiGeBPs secondary structure
2.3 杧果GeBP基因家族保守结构域与三级结构的分析结果
利用NCBI在线软件分析得到的MiGeBPs保守结构域如图2a所示。从图2a中可以看出,MiGeBPs基因家族的各个成员都有保守结构域DFU573(domain of unknown function 573)基序,其保守结构域位置见表1,其氨基酸数为66~96个,MiGeBP1、MiGeBP2、MiGeBP3、MiGeBP4、MiGeBP5、MiGeBP6、MiGeBP7、MiGeBP8、MiGeBP9、MiGeBP10蛋白中所含有的氨基酸数分别为70、71、96、91、96、69、89、90、92、66个。这些数据清晰地显示出,MiGeBPs中10个基因家族成员其保守结构域的位置存在差异;另外,MiGeBP3蛋白质中还有含Fe(SCys)4中心的非血红素铁结合结构域。
基于同源建模原理,通过SWISS-MODEL在线软件预测MiGeBPs基因家族成员的三级结构,结果如图2b所示。预测发现,MiGeBP6、MiGeBP7、 MiGeBP8、MiGeBP9、MiGeBP10蛋白质三级结构模型的相似性极高,且无规则卷曲和α-螺旋均为10个基因家族成员结构的主要组成部分,这一预测结果与该基因家族蛋白质二级结构的预测结果(图1)一致。
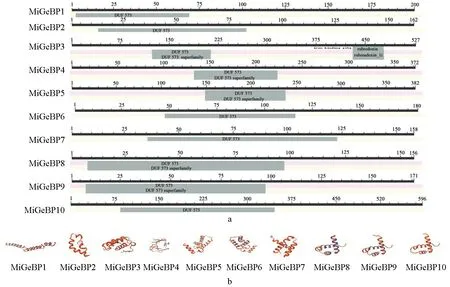
图2 MiGeBPs保守结构域和三级结构Fig.2 MiGeBPs conserved domain and tertiary structure
2.4 杧果GeBP基因家族10个成员的保守基序分析
利用MEME Suite(Suithttps://meme-suite.org/meme/)在线网站对杧果GeBP的10个基因家族成员的序列进行分析,结果如图3所示。图3表明,MiGeBPs基因家族成员包含1~5个基序,均含有基序4,其中,基序5的序列最长(48个)且最为保守,但仅有MiGeBP2、MiGeBP8和MiGeBP9成员中含有基序5;MiGeBP3、MiGeBP4、MiGeBP5、MiGeBP6、MiGeBP9和MiGeBP10蛋白中均包含了4个基序,其自N端向C端的排列顺序依次是基序1、基序2、基序4、基序3。
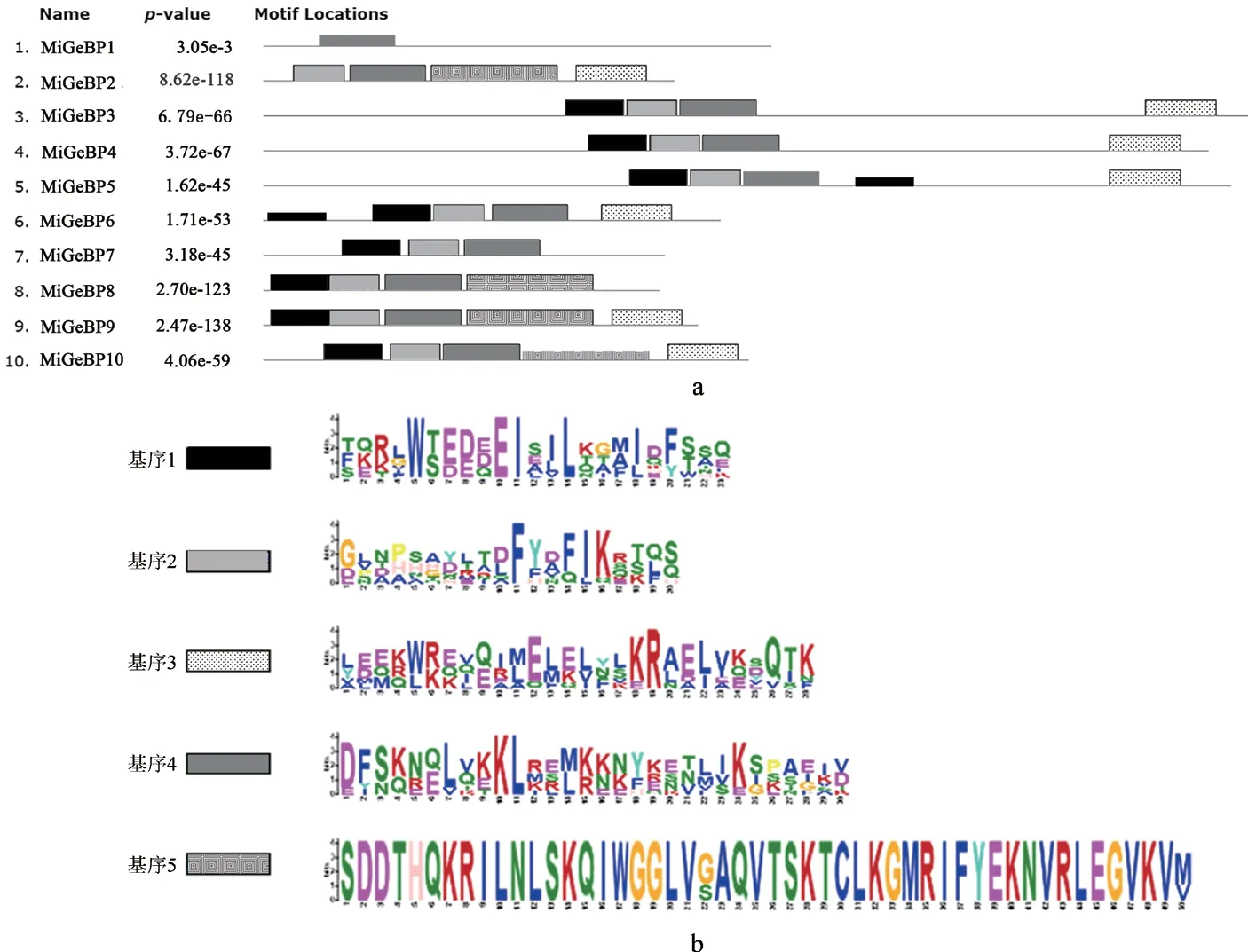
图3 MiGeBPs蛋白质保守基序Fig.3 Conserved motifs of MiGeBPs proteins
2.5 系统进化树的分析结果
利用ClustalW程序和MEGA 7.0在线软件构建了包含杧果(10个)、拟南芥(23个)、番茄(11个)、水稻(13个)的GeBP基因家族成员的系统进化树,结果如图4所示。图4表明,4个物种的57个氨基酸序列的系统进化树可以分为5个分支(ClassⅠ~Ⅴ),其中的MiGeBPs分 布 在4个分支中。MiGeBP2、MiGeBP7、MiGeBP8和MiGeBP9均分布在Class Ⅰ分支中,但MiGeBP2、MiGeBP8和MiGeBP9与 番茄的Solyc05g051330.1.1聚为1个小的分支,而MiGeBP7与拟南芥的2个成员(AT5G41765.1、AT4G00232.1)聚为1个小的分支;MiGeBP5、MiGeBP6均分布在Class Ⅱ分支中;MiGeBP1、MiGeBP10均分布在Class Ⅲ分支中;MiGeBP3和MiGeBP4均分布在Class Ⅳ分支中,且聚为1个小的分支;Class Ⅴ分支中聚集了拟南芥中的AT4G00390.1和水稻中除LOC Os01g14720.1之外的12个水稻GeBP基因家族成员。综合分析可知,杧果GeBP基因家族成员全部分布在以双子叶为主的Class Ⅰ、Ⅱ、Ⅲ、Ⅳ分支中,没有家族成员分布在以单子叶水稻为主的Class Ⅴ分支中。

图4 杧果GeBP蛋白的系统进化树Fig.4 Phylogenetic tree constructed from GeBP proteins
2.6 病原菌侵染时MiGeBP基因家族的表达分析
利用Quant Studio 6 Flex软件进行实时荧光定量PCR引物的溶解曲线检测,结果如图5所示。从图5a—b中可以看出,供试的10个MiGeBP基因家族成员的qRT-PCR引物的溶解曲线均具有单一的峰值,说明引物具有特异性,可用于后续的实验分析中。采用琼脂糖凝胶电泳(2%)检测[29]实时荧光定量PCR的产物,结果(图5c)显示,目的基因扩增出的条带与预期的结果一致,这进一步说明所设计的引物具有特异性,并可用于后续的实验分析中。
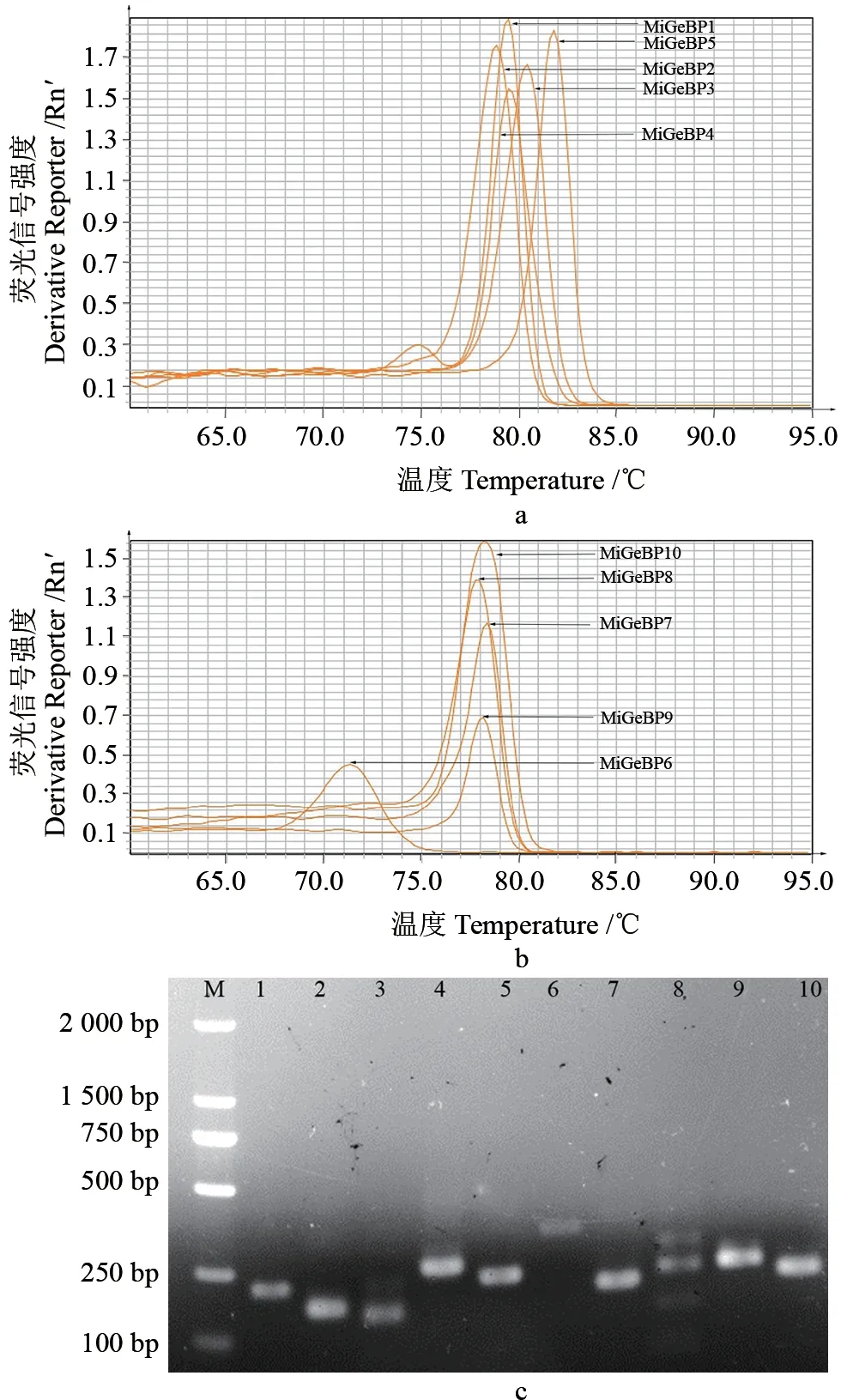
图5 杧果GeBP基因的溶解曲线及琼脂凝胶电泳图Fig.5 GeBP gene dissolution curve and AGAR gel electrophoresis of M.indica
2.7 MiGeBP基因家族在杧果各组织中的特异性表达分析
利用qRT-PCR方法分析获得相应的Ct值,采用 2-ΔΔCt法进行数据统计,以老叶(35 d)的相对表达量作为对照(记为1),将相对表达量高于1.5的定义为显著上调表达,将相对表达量小于0.5的定义为显著下调表达,统计结果如图6所示。图6表明,在供试的8个组织中,MiGeBP9、MiGeBP10基因在4个组织中均显著上调表达,MiGeBP9基因在芽、盛花、大果(35 d)和成熟果中均显著上调表达,MiGeBP10基因在幼叶(7 d)、盛花、大果(35 d)和成熟果中均显著上调表达;MiGeBP2、MiGeBP3、MiGeBP7和MiGeBP10基因在4个组织器官中均有显著表达。
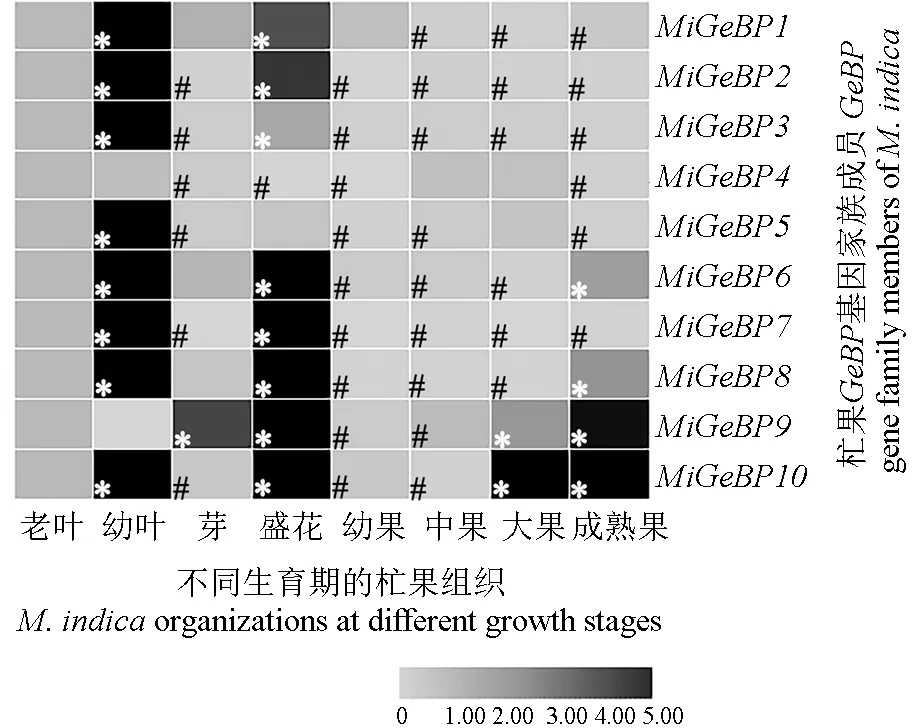
图6 杧果GeBP基因家族在不同组织相对表达量热图Fig.6 Heatmap of relative expression of M.indica GeBP family in different tissues
在杧果各组织中进行特异表达的10个GeBP基因家族成员中,MiGeBP1基因在幼叶和盛花中均上调表达,且在幼叶中的相对表达量最高,其在中果(21 d)、大果(35 d)、成熟果中的表达量均显著下调;MiGeBP2、MiGeBP3和MiGeBP7基因在各组织中均显著表达,但此3个基因都仅在幼叶(7 d)和盛花中显著上调表达,而在芽、幼果(7 d)、中果(21 d)、大果(35 d)及成熟果中的表达均显著下调;MiGeBP4基因在芽、盛花、幼果(7 d)和成熟果中的表达均显著下调;MiGeBP5基因仅在幼叶中上调表达,且其相对表达量最高,而在幼果中的相对表达量最低;MiGeBP6基因在幼果(7 d)、中果(21 d)、大果(35 d)中的表达均显著下调,大果(35 d)中的相对表达量最低,在幼叶(7 d)、芽及成熟果中上调表达,其在幼叶中的相对表达量达到最高;MiGeBP8基因在除芽以外的其他组织中均显著表达,其在幼叶(7 d)、盛花和成熟果中均显著上调表达,而在幼果(7 d)、中果(21 d)和大果(35 d)中均显著下调表达;MiGeBP9基因在芽、盛花及大果(35 d)、成熟果中均显著上调表达,而在幼果(7 d)、中果(21 d)中均显著下调表达;MiGeBP10基因在各组织中均显著表达,而在芽、幼果(7 d)和中果(21 d)中均显著下调表达,但在其他组织中均上调表达,其在成熟果中的相对表达量最高,而在中果(21 d)中的相对表达量最低。综上所述,MiGeBP基因家族成员在杧果不同组织中的表达存在特异性。
2.8 病原菌侵染时MiGeBP基因家族的表达分析
利用qRT-PCR方法获得相应的Ct值,采用2-ΔΔCt法进行数据统计,以MiGeBP基因家族成员经细菌性黑斑病菌或胶包炭疽菌侵染0 h时的表达量作为对照(记为1),将表达量高于1.5的定义为显著上调表达,将表达量小于0.5的定义为显著下调表达,统计结果如图7所示。图7a表明,经细菌性黑斑病菌侵染后,在2个及2个以上检测时间点均上调表达的基因分别有MiGeBP2、MiGeBP4、MiGeBP9和MiGeBP10,在2个及2个以上检测时间点均下调表达的基因分别有MiGeBP1、MiGeBP 7、MiGeBP8、MiGeBP10;在侵染处理后6 h时各个基因都有显著性的表达,其中,MiGeBP1、MiGeBP3、MiGeBP4、MiGeBP5基 因均下调表达,其他基因均上调表达。MiGeBP8基因在侵染处理后的各个检测时间点均显著表达,但仅在侵染处理后6 h时是上调表达的,而在其他检测时间点均下调表达;MiGeBP9基因同样在侵染处理后的各个检测时间点都是显著表达的,其在侵染处理后24 h时内一直下调表达,而在其他检测时间点均上调表达。
图7b表明,经胶孢炭疽菌侵染后,MiGeBP5、MiGeBP6基因在0~72 h内的各个检测时间点均显著上调表达,分别在侵染处理后24、12 h时的相对表达量达到最高;另外,在3个及3个以上检测时间点均上调表达的基因分别有MiGeBP1、MiGeBP2、MiGeBP3、MiGeBP10,MiGeBP1、MiGeBP2、MiGeBP3、MiGeBP10基因分别在侵染处理后24、48、24、12 h时的相对表达量最高,在2个及2个以上的检测时间点均下调表达的基因是MiGeBP9,其在侵染处理后3 h时的相对表达量最低。侵染处理后的48 h时,MiGeBP基因家族的多个成员均显著表达,而MiGeBP7和MiGeBP9基因均下调表达。
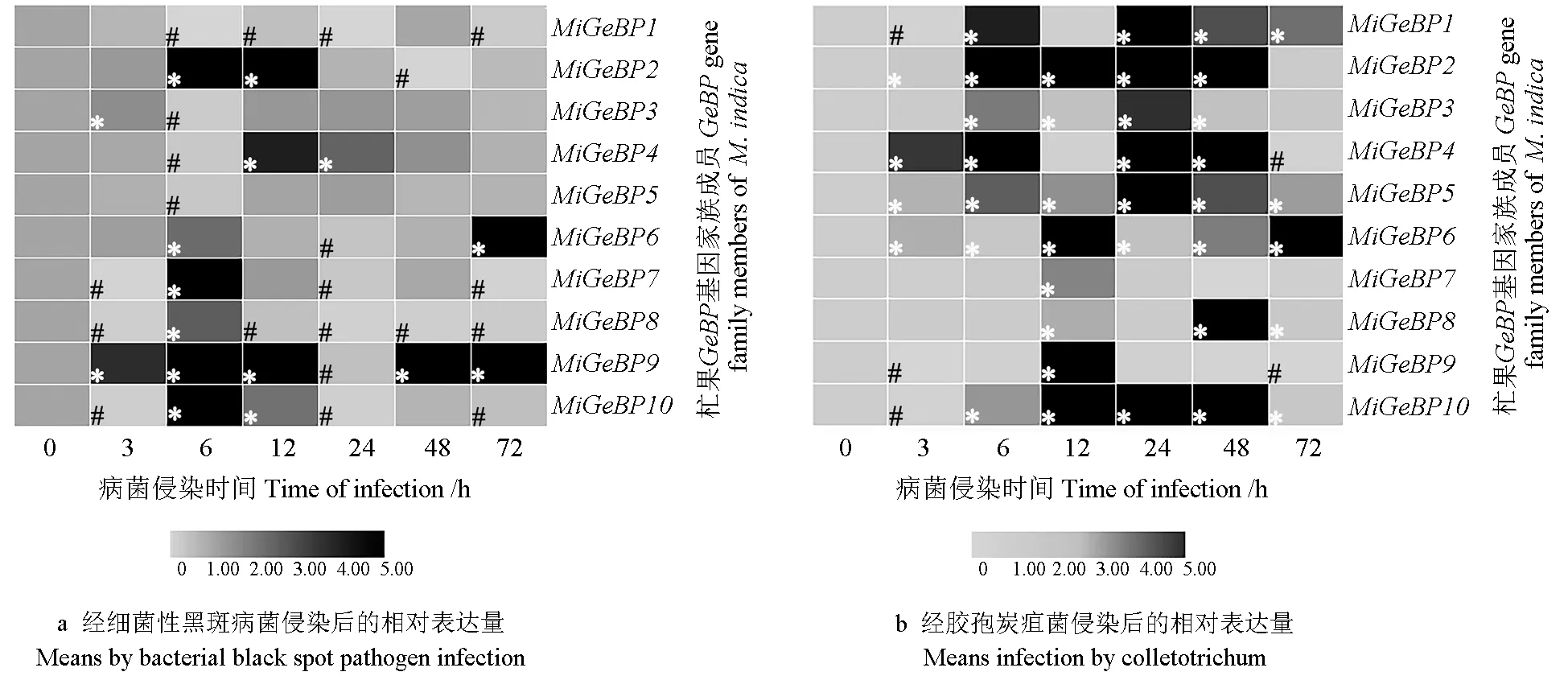
图7 杧果GeBP基因家族在病原菌侵染后的相对表达量热图Fig.7 Heatmap of relative expression of the mango GeBP family after pathogen infection
3 讨 论
植物转录因子存在特殊结构,可以调控基因表达,是重要的调节基因之一,如与植物表皮毛形成密切相关的GeBP转录因子。GeBP中的亮氨酸拉链(75%相似)和基本域(75%相似)两个基序的排列特点及邻近性表明了GeBP可能属于bZip转录因子家族,但两个基序间的距离大于9个残基,所以,其又不完全属于标准意义上的bZip蛋白[12,30],通过敲除拟南芥的GeBP(AtGeBP)中心域、bZip基序的假定基本区域和C端区域后发现,基本结构域的删除对酵母的生长没有影响,这表明GeBP没有bZip转录因子的作用,因为基本结构域是bZip蛋白结合DNA所必需的[12]。以上结果表明,这是一个新的蛋白家族。
杧果基因组测序的完成,有利于杧果转录因子功能的深入研究。本研究从全基因组水平上对杧果GeBP基因家族的10个成员进行了鉴定。结果表明,MiGeBPs转录因子在氨基酸个数、蛋白质分子量、等电点上均有较大的差异,且除MiGeBP3中含有2个内含子之外,其余家族成员都只有1个内含子;而毛竹的16个基因中含有内含子的基因仅有3个,番茄的10个基因中含有内含子的基因仅有2个。这一鉴定结果表明,GeBP基因家族结构较为简单、稳定。对GeBP基因家族成员的蛋白质二级结构的预测结果表明,无规则卷曲和α-螺旋是其结构中的主要元件;在对MiGeBPs保守结构域的分析中发现,该家族成员均有DUF573结构域,这一结果与陈凯等[10]对番茄GeBP转录因子家族的研究结果一致,说明DUF573结构域在该基因家族中具有普遍性。在构建杧果与番茄、拟南芥和水稻这4个物种的系统进化树时发现,MiGeBP2、MiGeBP3、MiGeBP4、MiGeBP5、MiGeBP6、MiGeBP7、MiGeBP8、MiGeBP9与番茄和拟南芥的GeBP家族成员分别聚集在ClassⅠ、Ⅱ、Ⅳ分支中,MiGeBP1、MiGeBP10所在Class Ⅲ分支的8个蛋白中仅存在水稻的1个蛋白(LOC Os09g014750.1.1),而水稻GeBP基因家族中的剩余成员与AT4G00390.1聚集在ClassⅤ分支中。由此可以推测,杧果与番茄、拟南芥均有较近的亲缘关系,而与水稻的亲缘关系较远。这一研究结果表明,杧果与双子叶植物的亲缘关系较近,而与单子叶植物间的亲缘关系较远。陈凯等[10]在构建番茄与拟南芥的系统进化树时发现,番茄的Solyc07g052760.1、Solyc07g052830.1、Solyc07g052900.1和拟南芥的GeBP/GPL间的关系均较近,他们由此推测,GeBP这个家族参与了番茄表皮毛细胞伸长的调控。研究中发现,上述番茄的Solyc07g052760.1、Solyc07g052830.1、Solyc07g052900.1这3个成员与MiGeBP 5均聚集在Class Ⅱ分支中,暗示着该基因可能与杧果表皮毛的形成或发育相关。
有关研究者在对番茄的研究中发现,番茄GeBP家族成员在其花、叶、根、果等组织中均有表达,但是不同的基因在不同组织中的表达量存在差异[10]。qRT-PCR检测结果表明,MiGeBP1~MiGeBP10在杧果不同组织中均表现出明显的组织表达特异性,其中,MiGeBP8基因在杧果的芽、盛花、大果(35 d)和成熟果中均显著上调表达,且在盛花中的相对表达量最高;MiGeBP4基因在杧果各组织中的表达量均较低。随着果实的发育,MiGeBP8与MiGeBP9基因的表达量均逐渐增高,且其在成熟果中的表达量最大。植物不同物种的GeBP转录因子在其不同组织中和不同发育时期均有差异表达[10-11]。黄瓜中的GeBP转录因子与GL1基因顺式调控原件结合调控该基因的转录,从而调控叶片、花、果实上毛状体的形成,但对根毛的形成却无明显影响,这也反映出GeBP组织表达的特异性[31];Lu等[32]采用扫描电镜观察了杧果叶片上的表皮毛,结果发现,老叶片上的表皮毛较少,这暗示着杧果叶片上表皮毛的形成受到了抑制。而笔者采用qRT-PCR的方法分析了杧果不同组织中GeBP转录因子的转录活性,结果发现,GeBP家族的多数成员在幼叶、花等器官中均上调表达,而在果实等器官中均下调表达,这说明GeBP家族成员调控杧果不同组织表皮毛形态建成的机制可能不尽相同。有关研究结果表明,植物在受到盐、干旱等胁迫时其表皮毛的长度与密度均会增加[33];Weinhold等[34]的研究结果表明,烟草凭借其表皮毛的分泌物抵御天蛾幼虫的侵害,这表明植物表皮毛在抗病抗逆中发挥着重要作用。而GeBP基因调控着植物表皮毛细胞的伸长,由此推测,GeBP基因直接或间接地参与了植物抗病抗逆反应。检测中发现,以分属于不同类型的杧果细菌性黑斑病菌和胶孢菌侵染杧果叶片时,MiGeBPs基因的响应程度有所不同:以细菌性黑斑病菌侵染时,多数基因表现为显著下调,由此可以推测,MiGeBPs基因以负调控的方式参与了杧果的免疫反应;而以胶孢炭疽菌侵染时,多数基因均持续性显著上调表达,这说明MiGeBPs基因以正调控的方式参与了杧果的免疫反应。因此,有关“病原菌—GeBP基因—表皮毛”三者之间的调控关系将是下一步研究的重点,通过研究其调控关系也许可以解析GeBP基因在植物免疫反应中的作用机制。
4 结 论
本研究采用生物信息学的方法分析了杧果GeBP转录因子家族基因成员的分子特征和表达模式,结果发现,该家族基因成员均属于DUF573超家族,其与双子叶植物的亲缘关系均较近。在病原菌的侵染过程中,各个家族成员均有不同程度的响应:以杧果细菌性黑斑病菌侵染后,MiGeBP8在侵染处理后0~72 h内的各个检测时间点均显著表达,除侵染处理后6 h时呈显著上调表达外,其余检测时间点均呈显著性下调表达,MiGeBP9在侵染处理后0~72 h的检测时段内也都呈显著性表达,除侵染处理后24 h时呈显著下调表达外,其余时间段均呈显著上调表达;以杧果胶孢炭疽菌侵染后,MiGeBP5、MiGeBP6在侵染处理后0~72 h的检测时段内均显著上调表达。由此可以推测,GeBP基因家族成员不同程度地参与了杧果的免疫反应。
