水稻抗稻瘟病分子机制研究进展
2022-01-14程新杰岳洪亮张梦龙朱国永代金英
程新杰,岳洪亮,张梦龙,施 伟,朱国永,代金英
(江苏沿海地区农业科学研究所,江苏 盐城 224000)
稻瘟病被称为水稻“癌症”[1],是由稻瘟病菌(Magnaporthe oryzae)引起的真菌性病害,是最为严重的水稻病害之一,且稻瘟病的危害面积和危害程度较大,发病时会影响粮食产量甚至造成绝收[2]。全球每年由稻瘟病导致的经济损失约为660亿美元[3]。因发病时期和部位不同,稻瘟病被分为苗瘟、叶瘟、节瘟和穗颈瘟等,其中尤以穗颈瘟对水稻生产的危害最大[4]。根据稻瘟病的发病条件,主要有以下防治方法:掌握防治适期,喷施相应的药剂;处理带病稻草和种子消毒,减少菌源;稀疏播种,培育无病壮秧苗;种植不过于密集;合理施肥、灌水,适时烤田;选用抗病良种等。在稻瘟病发病时,化学防治是最为快速有效的方法,但该方法对于环境危害巨大,不符合当今生态农业的发展目标。
目前在水稻新品种审定中,稻瘟病抗性是不可或缺的评定标准。由于稻瘟病的生理小种具有高度变异的潜力,其新基因型的产生不可避免地降低对水稻中抗性基因的敏感度[5],同时,稻瘟病菌会产生抗药性,进而使得化学农药的防治效果不断降低,这也是优良抗性品种在种植3~5年后抗性会逐渐丧失的原因[6]。因此,目前防治稻瘟病最安全有效且对环境友好的措施是挖掘新的抗稻瘟病基因,用来选育新的抗稻瘟病品种[7]。基于水稻稻瘟病的研究现状与成果,本文主要总结了近年来水稻稻瘟病抗病机制的研究进展,对该领域的研究热点和难点进行讨论,并对未来的防控方向提供一定的思路,为今后的抗病育种工作提供一定的理论基础。
1 水稻稻瘟病菌的侵染与防御机制
1.1 水稻稻瘟病菌侵染机制
随着科技的进步,科研人员对稻瘟病菌侵染过程的了解更为深入,侵染路径也已被解析[8]。稻瘟病菌是半活体的营养型真菌,其侵染过程有2个阶段:在侵染水稻的初期处于活体营养阶段,在这一时期主要是抑制寄主的免疫反应,进而定植在寄主细胞内;接着进入死体营养阶段,在这一时期会导致寄主细胞坏死。
1.2 水稻稻瘟病防御机制
在协同进化过程中,水稻也同样进化出复杂的防卫机制来应对稻瘟病菌的侵染,其主要有2层先天免疫系统,分别是病原菌相关模式分子诱发的免疫机制(PAMP-triggered immunity,PTI)与病原菌效应分子诱发的免疫机制(effector-triggered immunity,ETI)[9]。这2种免疫系统均能够诱导水稻产生稻瘟病抗病性,且这一免疫过程又被细分为3个步骤:首先是信号感知,病原菌中的PAMPs(pathogenassociated molecular patterns)或效应分子被各种受体识别;第2阶段是通过G蛋白、Ca2+流等将信号传递并放大之后,能够进一步激活丝裂原活化蛋白激酶和NADPH(还原性辅酶Ⅱ)氧化酶,进而释放活性氧;第3阶段是诱导防卫基因表达,并且积累抗病原物的次级代谢产物,使得细胞壁加厚,最终导致侵入位点的细胞程序性死亡等[10-12]。
1.2.1 稻瘟病PTI天然免疫系统。PTI天然免疫系统又被称为基础抗性[13],稻瘟病菌分泌的PAMPs主要是被水稻细胞表面的模式识别受体(patternrecognition receptors,PRRs)识别。目前在拟南芥中,对于PTI防御机制已经有了较为系统的研究。近年来研究人员对于水稻PTI机制的研究也取得了极大的突破。如flg22与flg22的受体OsFLS2,包括其介导的内部信号通路等方面的研究均有所突破。flg22是一种细菌鞭毛蛋白,它的蛋白终端是由22个氨基酸组成的保守多肽,这一肽段具有很强的抗菌性,是鞭毛蛋白诱导植物产生抗性的重要结构[14]。flg22与其受体OsFLS2结合之后,能够有效地激活抗病结构,这不仅提高了水稻的抗病性,同样也能够从根本上减少水稻稻瘟病的发生[15]。
1.2.2 稻瘟病ETI天然免疫系统。这一系统被分为2个部分,一部分是稻瘟病抗性基因的有效克隆,另一部分是抗性基因R与无毒蛋白Avr作为介质的ETI激活机制。将已克隆的抗性基因与无毒基因组合在一起后,能够不断调整ETI防御机制[16],不仅能够使得水稻稻瘟病防御机制更好地发生,还能有效减少病虫害的发生,提高稻米产量。阐述稻瘟病抗性基因与其对应的无毒基因间的互作关系,是解析稻瘟病ETI防御机制的关键。
2 水稻稻瘟病抗性基因
近年来,国内外研究人员通过对水稻稻瘟病抗性基因进行大量系统化的研究,使用分子标记等方法定位了100余个稻瘟病主效抗性基因,目前已经被克隆的基因有36个,如表1所示。其中已克隆的稻瘟病抗性基因大部分位于第6和第11号染色体上,少量位于第1、第9、第12染色体上,同时第2、第4、第8号染色体上还各有1个抗性基因。第6号染色体上的抗性基因主要有Piz、Pi2、Pi9、Pizt、Pid2、Pid3、pigm、Pi5、Pi26等[17],第11号染色体上的抗性基因主要有Pia、Pik、Pi1、Pikm、Pikp、Pb1等。
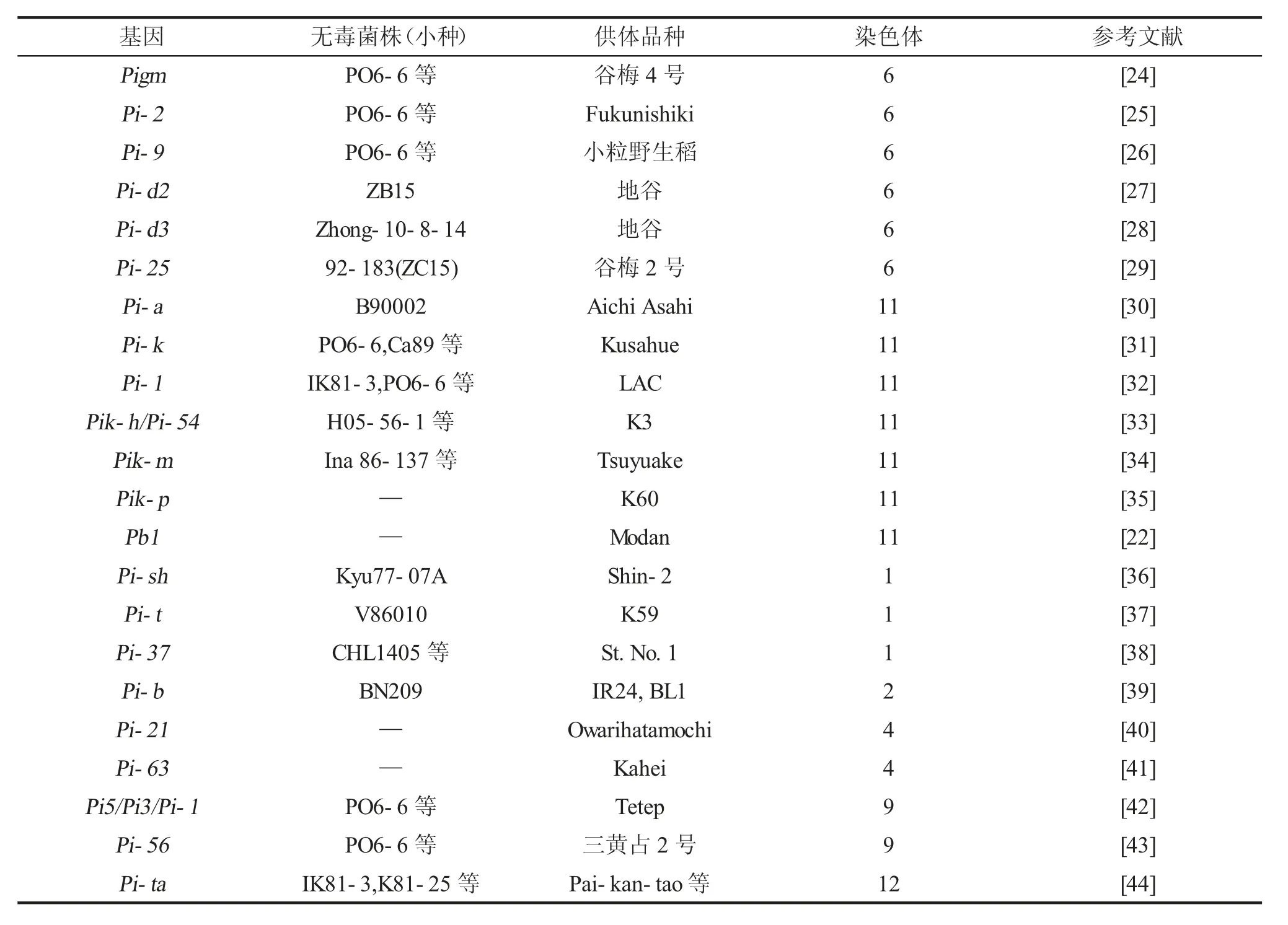
表1 已克隆的抗稻瘟病基因信息
这些抗性基因不仅表型特征不同,其来源与抗病地区也不同,不同的抗性基因抗性谱也不同。而稻瘟病菌的高度多样性和变异性等特点,也是抗性基因多样性的原因。Pigm是一个位于第6号染色体的广谱性稻瘟病抗性基因,供体亲本为谷梅4号,其与Pi2、Pi9、Piz-t、Pi26等基因同属于一个基因家族或是互为复等位基因,但彼此之间的抗谱具有明显差异。Pigm基因在粳稻中基本不存在,但对江苏、安徽、湖北、广东等地区的代表性稻瘟病菌种均有较好的抗性,统计表明,其对于来自世界各地的50余种菌株均表现出抗性[18]。Pi2抗菌谱也相当广,对来自我国13个地区的790余种菌株均表现出抗性[19]。Pi9同样也是一个稻瘟病广谱抗性基因,对21个稻瘟病小种具有抗性[20]。
穗颈瘟对于水稻危害最为严重,而Pb1所介导的抗性是持久性的,并且是成株期抗性和穗颈瘟抗性[21],这种抗性已被用于许多优良品种中[22]。研究表明,在育种工作当中,可通过选择合适的基因组合来更好地达到对稻瘟病的抗性作用,如Pita+Pia或Pita+Pi3/5/i双基因聚合的水稻品种均有高抗病的特性[4]。
目前已克隆的36个稻瘟病抗性基因对于稻瘟病产生抗性的生物学过程各有不同,根据蛋白结构的不同被细分为4类:一是NBS-LRR(nucleotidebinding site and leucin-rich repeat)类蛋白,如Pi1、Pib、Pi9等29个抗性基因;二是RLK(receptor like kinases)类蛋白,如Pid2,主要通过单个氨基酸差异来区别抗感病基因;三是富含脯氨酸结构域蛋白,如Pi21,主要通过蛋白功能的失活来产生抗性;四是富含ARM重复序列蛋白,如Ptr,其抗性与Pita相关,且是单子叶植物所特有的基因[23]。
3 水稻抗性蛋白与稻瘟病菌无毒蛋白间的互作机制
无毒基因作为一类能够诱导植物产生抗病性的病原物遗传因子,在稻瘟病菌与水稻植株产生相互作用的过程中,能够转录翻译成无毒蛋白,接着水稻中的R基因能够识别相对应的无毒蛋白,进而产生抗性[45]。目前从稻瘟病菌中鉴定并克隆的无毒基因有12个,分别是AvrPita、PWL1、PWL2、ACE1、AvrPiz-t、Avr-Pik/km/kp、Avrpi9、Avr-Pii、Avr-Pia、Avr-Pib、AvrPi54、AvrPi-CO39。已被克隆的无毒基因中,除ACE1外都编码分泌蛋白,而ACE1不分泌但编码聚酮合成酶[46]。
目前已成对的抗性基因和无毒基因有Pi-ta/AVR-Pita[47]、Piz-t/AvrPiz-t、Pi9/Avrpi9、Pi-33/ACE1、Pia/Avr-Pia、Pib/Avr-Pib、Pii/Avr-Pii、Pik/km/kp/Avr-Pik/km/kp、Pi-CO39/AvrPi-CO39、Pi54/AvrPi54。除Pi9/Avrpi9、Pi-33/ACE1、Pib/Avr-Pib互作关系未知外,绝大部分水稻稻瘟病抗性蛋白与无毒蛋白间的互作关系已被解析,可分为直接互作与间接互作。Piz-t/AvrPiz-t、Pii/Avr-Pii属于间接互作,Pi-ta/AVR-Pita、Pia/Avr-Pia、Pik/km/kp/Avr-Pik/km/kp、Pi-CO39/AvrPi-CO39、Pi54/AvrPi54则属于直接互作。
抗性蛋白与无毒蛋白直接互作有3种方式:一是符合经典“基因对基因学说”的一个抗病蛋白对应一种无毒蛋白,Pi-ta与AVR-Pita[48]对应这种互作关系;二是2种抗病蛋白成对出现,且只对应一种无毒蛋白,Pik-1和Pik-2与AVR-Pik之间对应这一互作关系;三是2种抗病蛋白成对出现,与多个无毒蛋白互作,如Os11gRGA4和Os11gRGA5对Pia的抗性都是必需的[49],但在防御反应的过程中,仅RGA5-A与AVR-Pia或AVR1-CO39互作,并起到重要作用。
4 展望
近些年来,由于分子生物学与生物信息学这2个学科的急速发展,水稻抗性基因的发掘鉴定、克隆及互作机制等方面的研究得到巨大进步。通过分子标记育种技术育成的抗病品种已经成功应用于农业生产当中。但是,随着病原菌生理小种的不断变异和种植环境的变化,单一稻瘟病抗性品种不足以应对这些问题,因此需要不断培育新的水稻稻瘟病抗性品种。基于以上问题,稻瘟病新基因的挖掘仍是水稻稻瘟病抗性分子研究的重要基础。基于目前的研究和育种现状,笔者认为还存在5个方面需要更深入的研究。1)分子育种缺乏广泛的有效抵抗多种稻瘟病菌的抗性基因。虽然近年来分子育种发展迅速,已经发现并克隆了大量的抗性基因,但这些基因在田间的抗性作用难以预测,导致在育种过程中,难以将这些抗性基因运用到相对应的致病菌株上。2)还需要开发创制更高效、且能够与抗性基因紧密连锁的分子标记,这对于育种工作中进行抗性基因的应用具有极大的帮助,能够将分子育种真正运用到实际育种工作当中。3)进一步开展抗性基因与无毒基因互作的分子机理研究,挖掘新的抗稻瘟病基因,并通过转基因技术、基因编辑技术、单倍体技术等作物育种技术,培育具有广谱抗性的水稻新品种。4)对调控水稻稻瘟病抗性的不同路径进行深入研究,寻找更多安全有效的防治方法。5)在育种工作当中,可尝试选择合适的基因组合进行应用,以达到事半功倍的效果,但对其抗病的广谱性与持久性仍需要进一步研究。
