肉类和水产制品脂质光氧化机制及其影响因素
2022-01-13顾赛麒鲍嵘斌冯媛张月婷周振毅黎君鸿朱士臣丁玉庭周绪霞
顾赛麒,鲍嵘斌,冯媛,张月婷,周振毅,黎君鸿,朱士臣,丁玉庭,周绪霞*
1(宁海县浙工大科学技术研究院,浙江 宁海,315600)2(浙江工业大学 食品科学与工程学院,浙江 杭州,310014) 3(国家远洋水产品加工技术研发分中心 (杭州),浙江 杭州,310014)
肉类和水产品在我国居民餐桌上占据重要地位,2019年全国消费总量分别达到9 420.3万t和3 285.5万t。除直接烹饪外,还可以通过腌腊、熏制、风干等手段加工成相关制品(如腊肉、腊肠、风鸭、腊鱼、鲞等),其风味独特、易于贮藏,广受消费者青睐。日晒法是上述制品常用的传统干制工艺,产品被报道拥有令人愉悦的特殊香气。
香气是食品的重要感官特征之一,产生途径较为复杂,脂质氧化降解被认为是肉类和水产制品香气生成的关键途径。甘油三酯、磷脂和游离脂肪酸是肉品中主要的脂质成分,不饱和脂肪酸(unsaturated fatty acids,UFA)是最典型的脂质氧化底物。按照反应类型分类,脂质氧化反应主要有3种:自动氧化、酶促氧化和光敏氧化。自动氧化是指活化的含烯底物与空气中的氧分子在室温下进行的氧化反应,前期反应速率较慢。酶促氧化是脂肪氧化酶(lipoxygenase,LOX)催化UFA发生的氧化反应。光敏氧化是指UFA双键与单线态氧(1O2)发生的氧化反应,需要有光敏剂(肌红蛋白等)的介导才可进行,反应速率是自动氧化的上千倍[1]。
目前,国内外学者较多关注酶促氧化反应,对脂质光氧化反应的研究报道较少,但后者可能对传统肉类和水产制品的香气形成贡献显著。张进杰[2]通过试验证实,经热风干制而成的腌腊鱼其香气感官得分均低于传统日晒品。传统肉类和水产制品一般选择在冬季晴日制作,环境温度较低且日照充足,该条件下LOX活性受到较大抑制,推测肌红蛋白等介导的脂质光氧化反应可能起主导作用。鉴于此,本文主要对脂质光氧化的基本原理、影响因素和常用测定指标进行综述,旨在为相关生产企业进行风味改良提供理论参考。
1 脂质光氧化反应的一般原理


图1 脂质光氧化的一般原理Fig.1 General principle of lipid photooxidation
两类光敏反应相互竞争,Ⅰ型取决于系统中氧溶解性和浓度;Ⅱ型取决于光敏剂和底物类型与浓度[4]。根据反应机制,光敏剂也可分为两类,第一类(底物光敏剂)对应于Ⅰ型途径;第二类光敏剂诱发的Ⅱ型反应为单线态氧和底物的协同反应,其氢过氧化位置异构体不同于自动氧化的产物,有几个不饱和碳原子便会产生几个位置异构体[4]。


图2 单线态氧在脂质光氧化中的作用机制Fig.2 Mechanism of singlet oxygen in lipid photooxidation
2 脂质光氧化反应的影响因素
脂质光氧化反应的影响因素可以分为直接因素和间接因素两大类,前者包括:光敏剂类型、光照条件和脂质构成等;后者包括:包装材料和方式、温度、水分、酶、盐度等。两类因素之间相互影响,在脂质光氧化反应中发挥重要作用。
2.1 直接因素
光敏剂是影响脂质光氧化反应的最重要的直接因素,其种类有:血红素类(肌红蛋白、血红蛋白、血红素、过氧化物酶、细胞色素)、叶绿素、胆红素、色氨酸以及维生素B族(核黄素、烟酸、钴胺素)[6]。除叶绿素外,光敏剂大多数存在于动物体内。肌红蛋白和血红蛋白是肉类和水产品中含量最高的光敏剂,对于屠宰放血后的动物而言,前者含量占80%以上。
2.1.1 肌红蛋白
Mb因动物种类、运动程度及肌肉部位的不同,其含量可能存在显著差异。牛羊肉中Mb含量(3.98 mg/g)显著高于猪肉(1.26 mg/g),鸡鸭等禽类肌肉中Mb含量较低(1.01 mg/g)[9]。水产动物中,洄游性鱼类Mb含量较高,如黄鳍金枪鱼肌肉中高达24.4 mg/g。高温、高压作用及食品添加剂均会影响Mb稳定性。Mb为外圆中空的分子结构,极性氨基酸几乎分布在分子表面,非极性氨基酸多分布在分子内部。血红素辅基位于疏水腔内,由位于中心的铁离子和卟啉环组成。60~70 ℃加热使中心铁原子和远端组氨之间的结合减弱,Mb三级结构发生局部变化,70~80 ℃加热严重扰乱Mb二、三级结构,发生不可逆变性,温度达80 ℃将导致铁卟啉结构被破坏[10]。高压作用破坏分子平衡力,使α-螺旋结构丧失,随机结构增加,Mb从高自旋(开放缝隙)结构转变为低自旋(闭合缝隙)结构。同时,高压离解离子对,破坏静电相互作用,引起珠蛋白变性和/或血红素移位及释放,促使血红素氧化为三价态[11]。据报道,金枪鱼肉经400 MPa处理后MetMb增加36%[12]。
为改善产品感官品质,肉类和水产制品加工过程中通常都会加入食品添加剂,这可能对光敏剂自身特性产生影响。如亚硝酸盐中的亚硝基能够置换OxyMb中的O2,生成稳定的亚硝基肌红蛋白[13]。烟酰胺可与Mb结合生成稳定的烟酰胺肌红蛋白。琥珀酸在琥珀酸脱氢酶的作用下被氧化成延胡索酸,产生的电子传递到MetMb使其被还原。在牛肉糜中添加6 mmol/L琥珀酸钠可有效抑制MetMb形成,贮存5 d后与对照组相比其含量降低了4.16%[14]。
2.1.2 核黄素(riboflavin,RF)


图3 核黄素的光化学反应机制Fig.3 Photochemical reaction mechanism of riboflavin
RF在不同动物种类和组织中含量差异较大。畜肉中RF含量约为水产品中的2倍,畜肉和淡水鱼中核黄素平均含量分别为0.17 mg/100g和0.09 mg/100g。值得注意的是,泥鳅和河蟹等水产品中RF含量较高,分别为0.33 mg/100g和0.71 mg/100g。动物内脏中RF含量尤为丰富,新鲜畜肉肝脏中含量平均达 0.45 mg/100g,猪心为0.32 mg/100g[17]。
加工过程中,光照及酸碱度对RF稳定性影响显著,温度对其稳定性影响较小。RF在阳光下会吸收紫外线反应形成CDRF[18]。RF在酸性及中性条件下较为稳定,碱性条件对其影响较大。研究发现pH=1和pH=7条件下将10 mg/L RF溶液黑暗密封5 h后保存率分别达94%~96.4%,而pH=13条件下其保存率仅为77.9%。RF光解速率在pH为5~6时最慢(一级速率反应常数为9.8~12.0 min-1),在pH=10时达到最大值(776.2 min-1)[18]。短时间高温处理可使结合态的RF释放,其含量略微增加,而长时间加热会导致RF自身结构发生热降解,其含量显著降低。新鲜猪肉糜在100 ℃处理15 min后,RF含量由0.14 mg/100g增加至0.18 mg/100g[19];10 mg/L核黄素溶液在100、120和150 ℃加热40 min其损失率分别为4%、7%和20%[15]。
2.1.3 金属离子
金属离子多以螯合物的形式存在于肉类和水产制品中,其一方面作为光敏剂结构组分影响光敏剂催化活性,也可以向氧分子转移能量和电子使其成为激发态,因而广义上可视为一类特殊的光敏剂。对于血红素类光敏剂而言,其基态或激发态性质较大程度受中心铁离子状态的影响。当铁离子游离出来后,剩余的原卟啉结构的促单线态氧生成能力反而强于血红素,推测原因是:铁离子与原卟啉的联结降低了血红素自身光敏性。此外,金属配合物如CrⅢ、PtⅡ、OSⅡ、IrⅡ、PdⅡ在光照条件下也可生成单线态氧,加速脂质氢过氧化物的形成[20]。此外,金属离子能够直接与未氧化脂质底物发生反应,缩短其氧化诱导期,加快氧化链式反应速率。

2.1.4 光照条件
光照条件对于脂质光氧化反应至关重要,一方面光照可使氧分子活化,促进自由基的生成,加速脂肪氧化;另一方面光敏剂经过光照后,可将基态氧(3O2)转变为激发态氧(1O2),进攻脂肪不饱和双键,生成氢过氧化物。光源波长和光照强度对脂质氧化程度均有显著影响。
光源类型分为单一光源和复合光源,前者包括:紫外光(10~380 nm)、蓝光(446~464 nm)、绿光(500~578 nm)、红光(620~760 nm)等;后者包括:太阳光、日光灯、白炽灯、LED等。短波长光(如紫外光、蓝光)是激活光敏剂发生光氧化反应的重要条件,不同光敏剂对各个波长光的敏感度不同。254和280 nm波长的紫外线能量较高,使拥有卟啉结构的血红素产生1O2和·OH;50 nm波长紫外线对RF破坏作用最大。光源波长越短,光的能量越高,诱发单线态氧和自由基生成能力越强。不同波长光连续照射4 d,其促进脂质氧化能力排序如下:蓝光>绿光>红光>黄光[22]。光照强度对脂质氧化也有重要影响,光照强度低于750 lx对猪肉脂肪氧化影响较小,高于750 lx脂肪氧化程度显著加深。霍晓娜等[23]发现,冷却猪肉 350~450 lx 贮存5 d后硫代巴比妥酸值与避光条件无显著差异,750~1 150 lx贮存5 d后,硫代巴比妥酸值由0.49 mg/kg增加至0.52 mg/kg。
2.1.5 脂质底物
脂质氧化底物主要有三类:甘油三酯、磷脂和游离脂肪酸。相对而言,游离脂肪酸最易发生氧化降解,其数值高低可以酸价表示。与甘油三酯相比,磷脂中长链多不饱和脂肪酸(polyunsaturated fatty acids,PUFA)含量更高且稳定性更低,较易发生脂质光氧化反应。不饱和度、双键位置、顺反构型、结合度均与脂肪酸氧化特性密切相关,不饱和程度越高越易氧化,共轭双键比非共轭双键易氧化,顺式比反式构型易氧化,游离脂肪酸比结合型脂肪酸易氧化[24]。
肉类和水产制品因其种类和部位不同,脂肪酸组成存在较大差异。鱼肉中不饱和脂肪酸(unsaturated fatty acid,UFA)含量(草鱼89.4%、鲤鱼76.09%)显著高于畜肉(猪肉54.9%、牛肉49.1%),其更易发生氧化降解[25]。在肝脏中,脂肪酸合成酶可将乙酰辅酶A合成棕榈酸和硬脂酸,因此肝脏中饱和脂肪酸(saturated fatty acid, SFA)含量较高。据报道,黄牛肝脏中SFA含量(88.09%)显著高于肌肉(61.06%)[26]。
高温、超高压、腌渍、超声等加工条件均会影响肉类与水产制品中脂肪酸组成。温度越高,PUFA被氧化程度越深。腌腊鱼在10 ℃和50 ℃干制时,PUFA含量分别为43.94%和40.98%[26]。超高压处理可使PUFA 氧化降解生成SFA,两者比值随着压力的增大而减少。压力从200 MPa升至600 MPa时,鸡肉早餐肠中的UFA/SFA比值从6.459降至5.627[27]。高浓度、长时间食盐腌渍可使肌内脂肪SFA增加,单不饱和脂肪酸(monounsaturated fatty acid,MUFA)和PUFA降低,这可能与食盐增强金属离子活性进而促进脂肪酸氧化有关。刘永峰等[28]等发现用10%食盐腌渍牛肉,SFA含量增加7.39%,MUFA和PUFA分别减少6.98%和0.63%。超声波的空化及热效应可以将脂肪颗粒从与肌肉组织致密结合状态中游离出来,使部分脂肪酸大分子产生聚合或解聚作用,从而改变脂肪酸组成。陈银基[29]报道,采用2%NaCl结合0.68 W/cm2超声波处理,其提升PUFA/SFA效果与6% NaCl处理基本相同。
2.2 间接因素
间接因素通过影响脂质光氧化反应直接因素,促进或抑制脂质光氧化反应速率,主要包括以下因素:包装材料和方式、温度、酶、水分活度、食盐等。
2.2.1 包装工艺
选择适宜的包装工艺(如包装材料、包装方式等)可以通过控制透光率和残氧量的方式影响脂质光氧化作用。不同包装材料对光的遮挡、反射和散射性能存在差异,使用透光率较低的包装材料,可有效抑制脂肪氧化反应。采用聚酯薄膜(polyethylene terephthalate,PET)、聚乙烯(polyethylene,PE)和铝箔作为原料,经过单一或者复合工艺制备得到的包装材料具有良好阻光性,如PET/PE在波长200~260 nm范围内光透过率为1%,而铝箔在波长200~900 nm波长范围内透光率小于0.9%[30]。
真空包装和气调包装是肉类和水产制品常用的包装方式,可以通过控制残氧量的方式抑制脂质氧化反应。研究发现,在残氧量大于0.5%的条件下,光照下会加速肉制品与水产制品脂质光氧化反应[31]。气调包装使用过程中采用特定比例的混合气体(CO2、O2、N2等)代替原包装中的气体环境,可起到抑制脂肪氧化的作用。CACHALDORA等[32]报道,西班牙血肠采用气调包装(40%N2-60%CO2)贮存8周后,硫代巴比妥酸值比真空包装处理样品降低0.51 mg/kg,表明气调包装可有效抑制脂肪氧化反应。
2.2.2 温度
温度对脂质构成、光敏剂结构和酶活性均有一定影响,高温可以降低形成过氧自由基(ROO·)所需活化能,有利于脂质氧化发生[33]。热加工阶段,PUFA最先被氧化降解,MUFA次之,SFA相对稳定。油酸酯、亚油酸酯、亚麻酸酯在24 ℃时氧化速率之比为1∶12.5∶25,在37 ℃时三者氧化速率之比为 1∶27∶77[34]。牛肉在55℃热风干燥至水分含量为50%过程中,UFA含量下降21.70%(MUFA和PUFA分别下降41.49%和11.81%),而SFA含量从40%显著增加至58%[35]。此外,热加工可以改变肌红蛋白等光敏剂结构:100 ℃高温加热,Mb自身二、三级结构发生剧烈变化,结构完整性被破坏,释放出铁离子促进脂质氧化[10]。高温也可造成高铁肌红蛋白还原酶全部或部分失活,导致MetMb无法及时被还原而大量积累,间接促进脂质氧化[10]。
2.2.3 酶
某些酶类可以作用脂质底物和肌红蛋白,间接影响脂质氧化。具体而言,脂肪酶可以催化甘油酯水解为游离脂肪酸、甘油及其他不完全降解产物[3]。添加0.02%脂肪酶,烤肠中游离脂肪酸含量增加,PUFA含量降低28.74%。添加0.02%~0.08%脂肪酶,烤肠脂质氧化程度相当于贮藏30 d的广式腊肠,添加0.10%脂肪酶相当于贮藏90 d的广式腊肠[36]。LOX属于非血红素铁双加氧蛋白酶,专一催化含有顺, 顺-1,4-戊二烯结构的多不饱和脂肪酸。当加工温度达到其最适温度(25~30 ℃)时,LOX具有较高的催化活性,迅速氧化PUFA使之生成含共轭双键的ROOH。高铁肌红蛋白还原酶是MetMb还原系统中的关键酶,在消耗ATP条件下可使MetMb还原为DeoMb,若该酶失活会使MetMb大量积累,促进脂质氧化反应。
2.2.4 水分活度(water activity,Aw)
Aw通过影响金属离子催化特性,间接影响脂质氧化。Aw在0.33时脂质氧化速率最慢;0~0.33范围内,随着Aw升高,脂质氧化速率降低,分析原因是:高度干燥样品中添加微量水分子,既能与金属离子水合降低其催化能力,又能与氢过氧化物结合阻止其降解。0.33~0.73范围内,随着Aw升高,金属离子流动性提高,水中溶解氧增多,分子溶胀且暴露出更多催化位点,最终导致脂质氧化速率提高。水分含量可能对脂肪氧化有一定促进作用,与干腌腊肉相比,湿腌法加工腊肉其酸价和过氧化值分别提高29.92%和7.14%[37]。当Aw>0.73时,含水量大幅增加,金属离子浓度被稀释,脂质氧化速率降低。Aw对酶活性也有显著影响,当Aw<0.8时,大多数酶活被抑制;Aw在0.35~0.30时,酸性脂酶活力提高,其他酶活力降低。
2.2.5 食盐
食盐是食品加工中最常用的添加剂之一,腌干制品中食盐含量甚至高达15%以上。高浓度食盐可以破坏细胞膜结构,促进UFA与各类氧化因子间发生相互作用。此外,高浓度食盐能够促进Mb向MetMb发生转变,加速铁离子从肌红蛋白中释放,也可以抑制肌肉中抗氧化酶活性,间接起到促进脂质氧化的作用。食盐对中性脂酶活性具有促进作用,而对酸性脂酶和磷脂酶活性具有抑制作用。食盐对LOX活性具有双重作用:低盐浓度下轻微促进LOX活性,高浓度下显著抑制LOX活性。此外,在高浓度条件下,食盐可以通过提高渗透压使细胞脱水,降低Aw,间接影响脂质光氧化。JIN等[38]报道,添加0.5%和4.0% NaCl的猪肉糜贮存6 d后,后者样品中硫代巴比妥酸值比前者提高近50%。
3 脂质光氧化反应的常用检测指标
根据光氧化反应的基本原理,本文从光敏剂、活性氧、脂质底物、氧化指标等方面,归纳总结了脂质光氧化反应的常用检测指标(表1),以期为脂质光氧化反应机制解析提供方法借鉴。
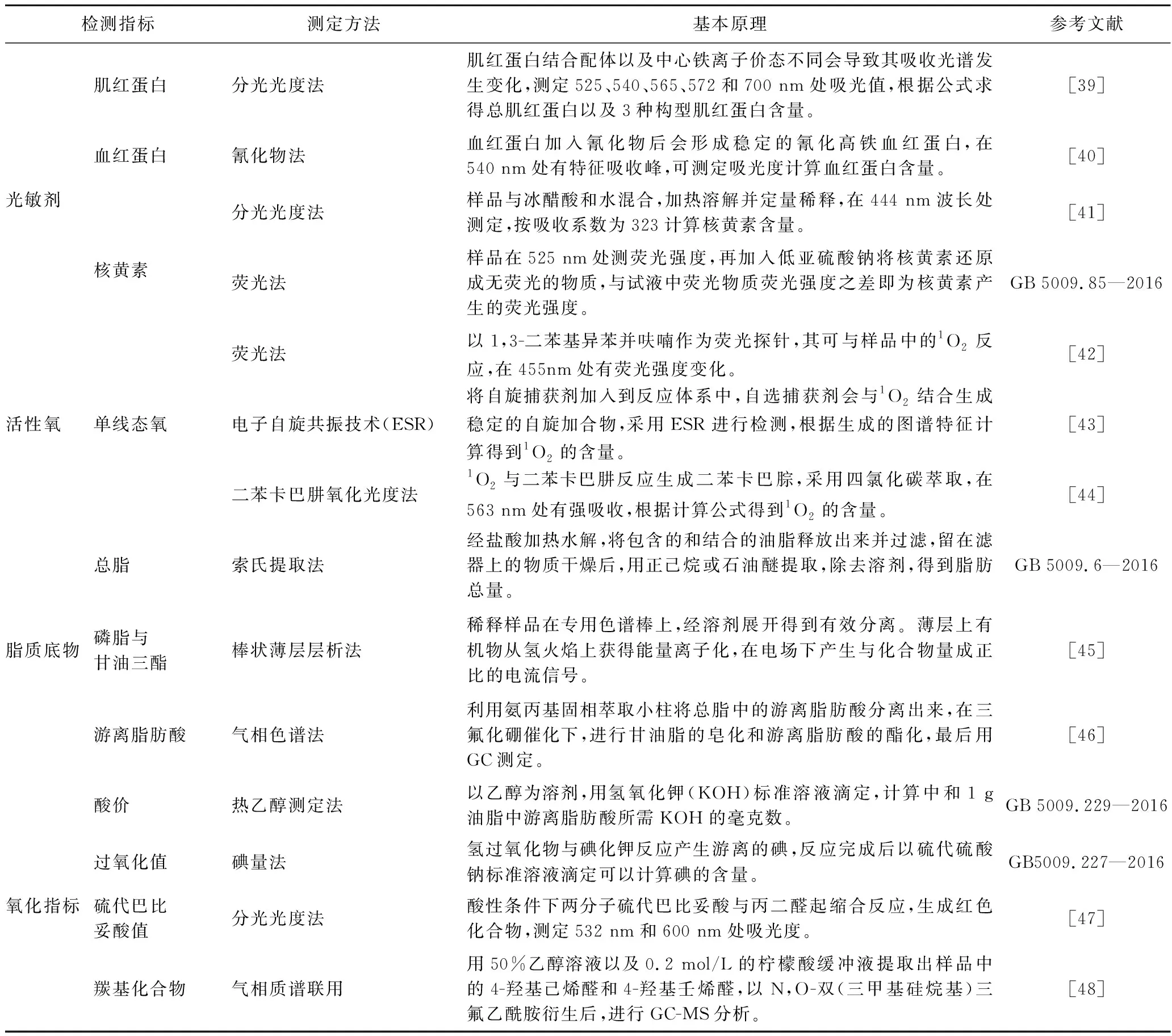
表1 脂质光氧化反应常用检测指标Table 1 Common detection indexes of lipid photooxidation reaction
4 展望
脂质光氧化反应对传统日晒品的优良风味贡献显著,本文对脂质光氧化反应的基本原理、影响因素及常用测定指标进行了综述。肉类和水产制品在实际加工过程中,除了光氧化反应,同时还会发生脂质的自动氧化和酶促氧化等反应。此外,肉类和水产制品中往往存在多种光敏剂(如肌红蛋白、血红蛋白、核黄素等),脂类底物构成十分复杂,不同脂类成分氧化特性存在差异,以上因素均可能对制品的最终香气产生影响。
对此,可以采用去繁从简的思路,构建样品模拟反应体系来解析脂质光氧化反应机制:基于嗅辨仪和电子鼻等分子感官技术探明特征日晒风味物身份,选择抑酶后的白色肉(不含肌红蛋白)作为空白样品,添加不同种类、不同浓度的光敏剂构建样品模拟反应体系,设置不同光强、光照温度和光照时间等光照条件,分析上述过程中光敏剂自身含量与结构变化规律,结合1O2等指标评估其促脂质氧化能力,探究脂质成分降解与优良风味物生成的关联性,最终解析脂质光氧化的具体作用机制。
对于产业化而言,在确认特征性日晒风味物身份并探明其风味形成机制基础上,可以集成光谱、频谱和温度、湿度、风速等多因素的动态调控法来模拟现实日晒条件,研发出一种可控模拟日晒风味的新型干制加工技术。将该项新技术应用于现代化肉制品和水产品加工企业的工业化生产,其产品可最大程度上保持传统日晒制品的典型优良风味,且较传统日晒制品相比质量稳定性更好、食用安全性更高,具有良好的应用推广前景。
