玉米矮秆突变体K718d的遗传鉴定
2022-01-10石海春赵长云余学杰柯永培
董 丽,石海春,,赵长云,余学杰,,柯永培,
(1.四川农业大学 农学院,四川 温江 611130;2.四川正红生物技术有限责任公司,四川 双流 610213)
在玉米生产中,高秆品种易倒伏,通常造成严重的减产;而矮秆玉米经济系数高,抗倒伏能力强,光能利用率高,利于密植高产。目前,已被定位的矮秆单基因有60多个(http://www.maizegdb.org/),包括隐性单基因br1、br2、d1、d2和d3等,显性单基因D8、D9、D11和Dt等,其中br2[1]、d3[2]、D8[3]、D9[4]和Dt[5]等基因已被克隆;分布于10条染色体上,已定位的控制玉米株高的QTL位点300余个(http://www.maizegdb.org/),但目前玉米中能够被应用的矮秆资源有限,遗传多样性低,主要集中于br2[6]。因此,为发掘新的玉米矮秆种质资源,并了解其遗传特性,对推动玉米矮化育种具有重要意义。本研究中,以自然突变所获得的矮秆突变体K718d为主要材料,研究其农艺经济性状表现和对外源激素敏感性;通过遗传交配设计,分析该矮秆性状的遗传模式,并初步定位该矮秆基因,为之后基因的精细定位、克隆及育种中的利用提供参考依据。
1 材料和方法
1.1 供试材料
野生型K718;矮秆突变体K718d(P1);5个高秆测验自交系(P2):K1208、Q3、Ly118、Na2和K338;以及K718d与测验自交系组配的正反交F1、BC1、BC2、F2等群体。5个已知矮秆基因材料114F(br2)、110K(br1)、301(cr1)、K15d(br2突变体)、K123d(br2突变体)与K718d杂交配制的F1组合。所有材料均由四川正红生物技术有限责任公司(简称公司)提供。
1.2 试验设计
1.2.1 K718d与K718形态差异比较 于2019年在公司双流区育种基地种植K718d与K718,选取其中30个有代表性的单株,对株高、穗位高等主要农艺性状进行考察;各小区中收获有代表性的10个果穗,考察主要经济性状。用t测验检测性状差异显著性,并对植株和果穗拍照对比。
1.2.2 K718d激素敏感性研究 选取K718d与K718种子浸种催芽,播于发芽盒,随机区组设计,3次重复,5个浓度处理,每处理25株,于两叶一心期用0,25,50,75,100 μg/mL的GA3和IAA溶液隔1 d浇灌1次,20 d后测量苗高及第一叶鞘长度;内源激素GA3测定参照徐敏等[7]方法测定。测定结果进行方差分析和多重比较分析(Excel 2010,Spss 20.0)。
1.2.3 K718d矮秆性状遗传模式分析 试验于四川崇州和雅安两生态地进行。各种植正反交F1群体84粒,BC1、BC2群体各168粒,F2群体各420粒。进行分离群体的株高鉴定,χ2检验株高分离比例,并确定遗传模式。
1.2.4 K718d矮化基因定位 定位群体为K718d×Ly118 F2,于四川崇州育种基地播种2 184粒。单株编号挂牌,调查记录高、矮植株。采用BSA-SSR法[8]进行基因定位。从定位群体中选取极高、极矮株各20株,提取DNA后等量混合,用于构建高、矮秆基因池。选取458对SSR引物于亲本K718d和Ly118间和高、矮基因池间筛选出多态性引物后,在定位群体中的所有矮秆单株间进行扩增。DNA提取用2×CTAB法,PCR体系为25 μL,用3%琼脂糖凝胶对扩增产物进行电泳检测,采用MAPMAKER 3.0、MAPDraw V2.1[9]软件进行连锁分析及遗传图谱的绘制。
1.2.5 K718d矮秆基因等位性鉴定 K718d与5个已知矮秆基因材料组配F1,并种植于公司海南陵水基地,各组合种植28粒,成熟期间观察F1株高表现,确定基因等位关系。
2 结果与分析
2.1 K718d与K718主要性状比较
K718与K718d形态对比见图1。K718d植株和穗位矮化,基部节间缩短,节间数减少,果穗变短。K718d与K718果穗均结实较好,筒型、籽粒半马齿型,但K718籽粒略长,K718d籽粒略宽。
K718d与K718主要性状差异列于表1。与K718相比,K718d生育期显著延长,株高、穗位高、节间数和节间长度分别减少48.23%,75.57%,30.83%和65.92%,差异均达极显著水平;K718d穗粗增加8.31%,穗行数和百粒质量差异不显著,但穗长和行粒数分别极显著降低28.57%,28.47%,导致单株产量极显著降低36.44%。
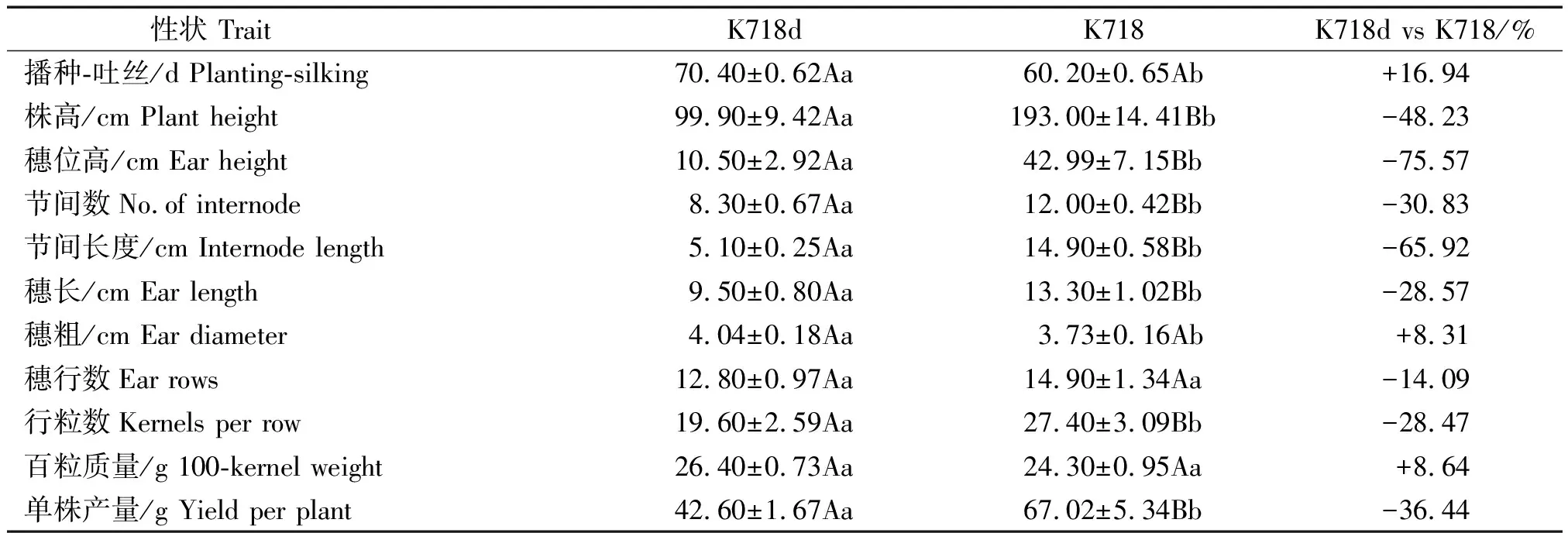
表1 K718d与K718主要农艺和经济性状比较Tab.1 Comparisons of the main agronomic and economic traits between the K718d and K718

A.植株 Bar=15 cm;B.茎秆 Bar=20 cm;C.节间 Bar=3 cm;D.果穗 Bar=3 cm;E.籽粒 Bar=1 cm。A.Plants bar=15 cm;B.Stem bar=20 cm;C.Internode bar=3 cm;D.Ear bar=3 cm;E.Kernel bar=1 cm.
2.2 突变体K718d对GA3和IAA的敏感性
经2种外源激素处理后方差分析结果,除内源GA3在材料间差异不显著外,其余性状在材料间和浓度间差异均达极显著水平,材料与浓度互作间差异不显著。K718d与K718比较,低浓度GA3处理下第一叶鞘长度差异不显著,高浓度下差异极显著,而各浓度处理苗高均达极显著差异,但内源赤霉素含量(以鲜质量计)差异不显著(表2);不同IAA浓度处理,2个材料间第一叶鞘长度和苗高均达极显著差异(表3)。2种激素处理,均不能使突变体苗高恢复至野生型水平,说明该突变体对GA3和IAA不敏感,但K718d能够正常合成赤霉素,且转运GA3途径正常。

表2 K718d和K718第一叶鞘长度、苗高、内源赤霉素含量的多重比较 (GA3)Tab.2 Multiple comparisons of the first sheath length,seeding height,concentrations of endogenous GA3 between K718d and K718
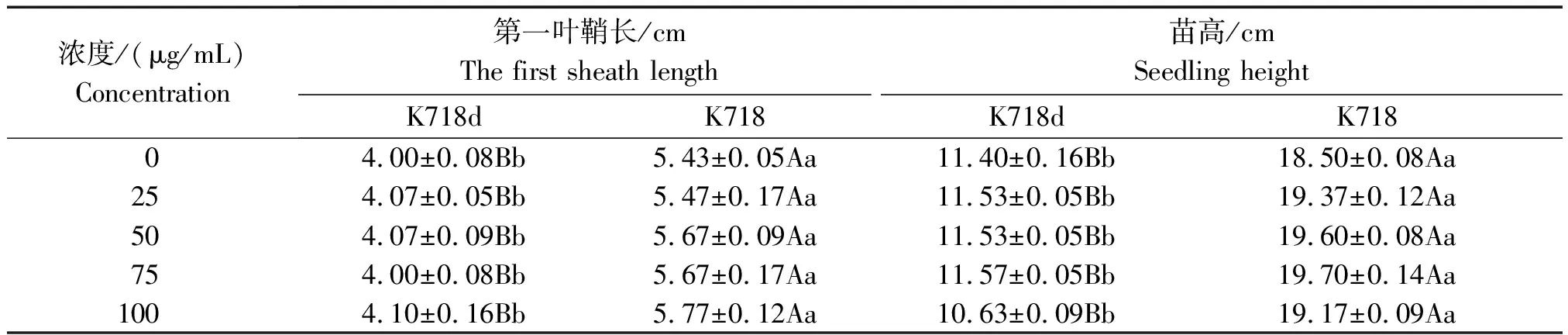
表3 K718d和K718第一叶鞘长度和苗高的多重比较(IAA)Tab.3 Multiple comparisons of the first sheath length and seeding height between k718d and K718(IAA)
2.3 突变体K718d株高遗传模式
2.3.1 正反交F1群体株高表现 将K718d配制的5个正反交群体平均株高列于表4。所有正反交F1在两试验点均表现为高秆,经t检验正反交F1群体平均株高差异不显著,初步说明株高无细胞质效应。

表4 正反交F1群体株高Tab.4 Plant height of reciprocal F1 populations
2.3.2 回交群体株高分离比例 以K718d为回交亲本构建的BC1回交群体,在两生态点高秆与矮秆植株分离比例经卡方检验均符合1∶1(表5);而以5个测验种为回交亲本的BC2回交群体在两试验点均表现为高秆,表明K718d矮秆性状可能由1对隐性核基因控制。
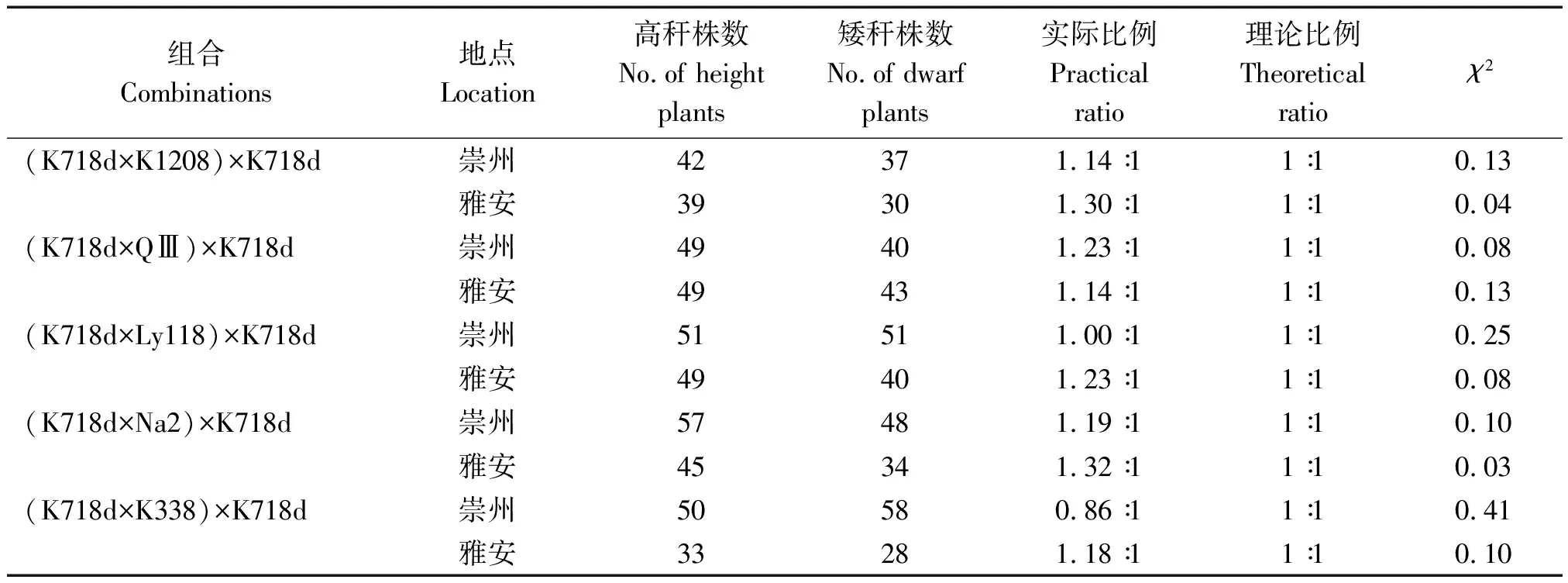
表5 BC1分离群体株高χ2检验Tab.5 The χ2 test of plant height of BC1 segregation populations
2.3.3 F2群体株高分离比例 在2个生态点所有F2群体高秆与矮秆植株分离比例,经卡方测验都符合3∶1,进一步证明K718d的矮秆性状由1对隐性核基因控制(表6)。
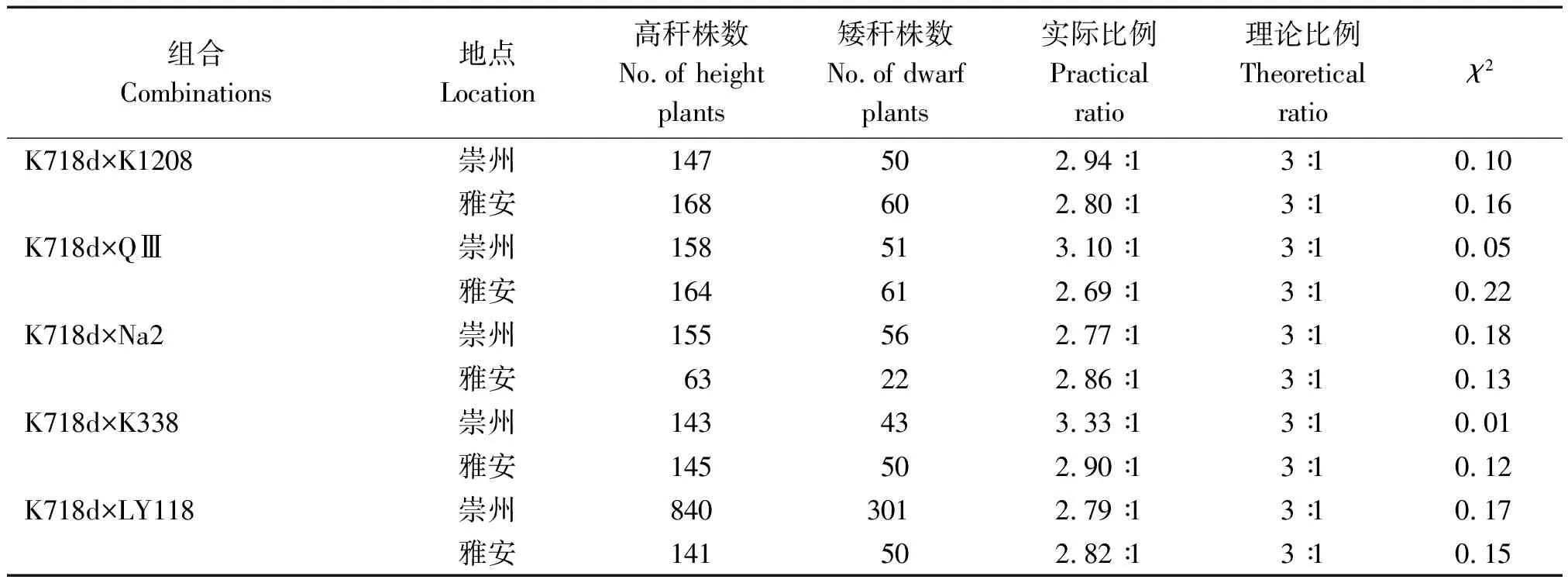
表6 F2分离群体株高χ2检验Tab.6 The χ2 test of plant height of F2 segregation populations
综上所述,K718d与5个测验系组配的正反交F1在崇州和雅安两地均表现为高秆,其正反交组合株高差异不显著,无细胞质效应,受环境影响较小;K718d与F1构建的BC1群体高矮秆植株分离比符合1∶1,5个测验系与F1构建的BC2群体都表现为高秆,F2群体高矮秆植株分离比符合3∶1,表明K718d矮秆性状受一对细胞核隐性基因控制,暂将其命名为d718。
2.4 矮秆基因d718定位
定位群体为K718d×Ly118 F2,选取458对均匀覆盖于玉米10条染色体上的SSR引物,在双亲之间筛选出了170对多态性较好的引物后,继续在高、矮秆基因池间筛选,筛出1对多态性引物umc1446;以该引物为参考,继续合成1号染色体Bin1.06~1.09区段的45对引物进行多态性筛选,筛选出了6对多态性引物,至此共筛选出7对多态性引物(表7)。图2为7对多态性引物在F2群体高、矮秆基因池间的扩增结果。

表7 7对SSR引物序列信息Tab.7 The sequence information of 7 pairs of SSR primers

D.高秆基因池;d.矮秆基因池。D.High stalk gene pools;d.Dwarf gene pools.
用上述7对引物在K718d×Ly118的F2群体的316个矮秆单株分别进行PCR扩增,用3%琼脂糖凝胶进行扩增产物的电泳检测。用上述作图软件进行扩增结果连锁分析后,绘制出遗传连锁图谱。最终将该矮秆基因d718,初步定位于玉米1号染色体长臂上,位于SSR分子标记umc1128与umc1278之间,遗传距离分别为1.0,2.5 cM(图3)。
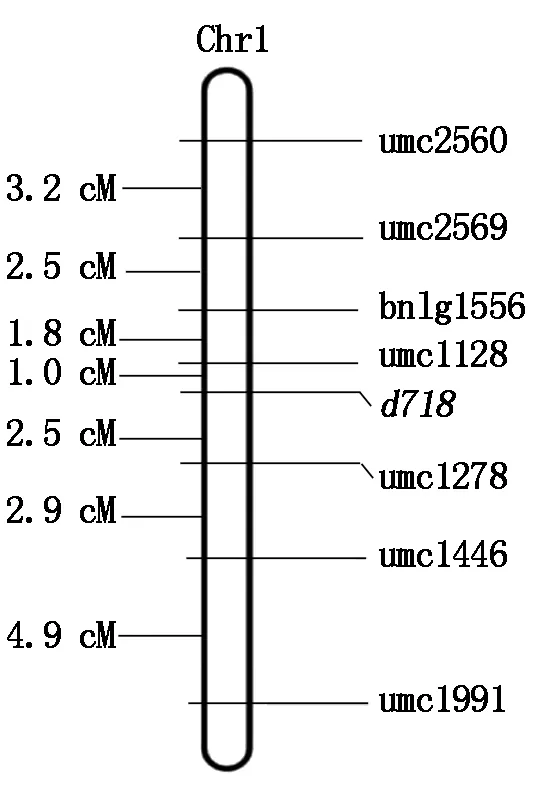
图3 矮秆基因d718遗传连锁图谱Fig.3 Genetic linkage map of dwarf gene d718
2.5 d718基因的等位性鉴定
由于矮秆基因d718在染色体上与br1(1.07)、br2(1.06)等已知矮秆基因相近,有必要对该基因进行等位性鉴定。K718d与5个矮秆测验系组配的F1群体株高统计发现,除K718d×110K株高表现为矮秆外,其他组合均表现为高秆(表8)。在5个矮秆测验系中,已知110K为br1材料,301为cr1材料,其余3个均为br2突变体,试验结果只有K718d与110K杂交F1群体植株为矮秆,由此表明d718为br1的等位基因。
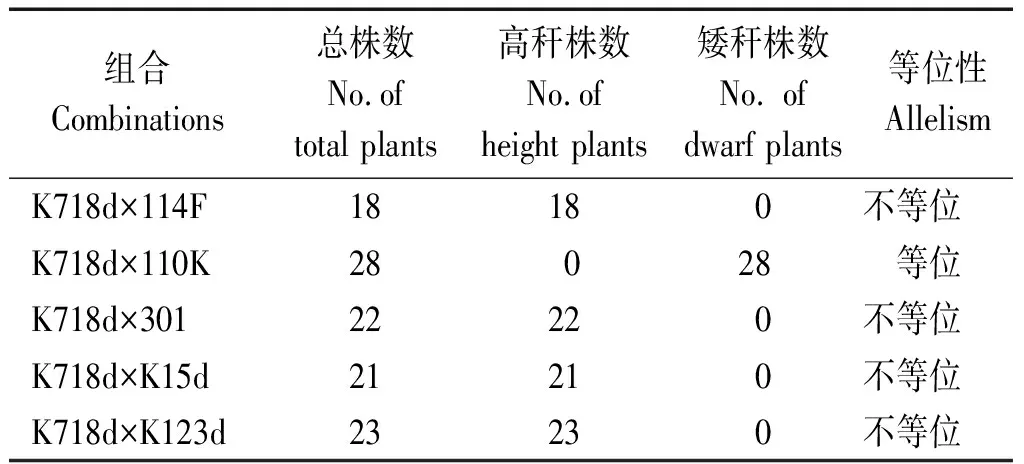
表8 F1群体株高Tab.8 Plant height of F1 populations
3 讨论与结论
3.1 矮秆突变体K718d表型特征
玉米茎秆节间数减少,节间长度变短可以造成株高的矮化,植物内源激素通过促进、抑制或改变生理活动,调控植物生长发育进程,参与植物株高建成[10-12]。前人研究表明,突变体A2节间数减少,节间长度缩短,dm676节间极显著缩短,从而导致植株矮化[13-14],br1类矮秆突变体节间显著缩短,平均单株产量只有野生型的2/3[15],且br1对GA3不敏感,而突变体523333[16]、d0掖(478)、d0(齐319)和d0(PH4CV)[17]对GA3敏感。
本研究发现,与野生型K718比较,K718d节间数减少近1/3,节间长度缩短近2/3,株高降低近1/2,平均单株产量降低36.44%,但穗粗增加8.31%,可用来改良玉米品种株高和穗型等性状,具有一定的应用价值。经不同浓度的GA3和IAA处理发现,该突变体为赤霉素与生长素钝感型,与矮秆突变体br1和br2相似,但赤霉素合成及转运途径正常。推测造成突变体K718d矮化的原因,可能是植株生长过程中某一特定时期激素合成或运输有关,有待进一步研究。
3.2 矮秆突变体K718d株高遗传特性
现有研究发现,玉米株高分为单基因遗传和多基因遗传2种模式,其中单基因遗传又分为显性和隐性2种[18-19]。王立静等[20]和戚洪源等[21]利用矮秆突变体与普通玉米自交系构建F1、BC和F2群体的方法,分析明确了矮生性状的遗传模式。王益军等[22]发现了来自玉米自交系Mo17的一个显性矮秆突变基因D*-10,并采用 SSR分子标记技术,将该基因定位在玉米2号染色体上。王立静等[20]把121C(D8)和502C(D9)的花粉授予玉米矮秆显性突变体52333,取30株F1的花粉授予高秆自交系Lx9801,根据后代高矮秆植株分离比例,确定了Dt与D8和D9为非等位基因。
本研究用K718d与5个高秆自交系组配正反交F1、BC1、BC2和F2群体,分析矮秆性状的遗传模式,结果发现,K718d矮秆性状由1对隐性核基因控制;并利用BSA-SSR分子标记技术,将矮秆基因d718定位于1号染色体长臂上,位于分子标记umc1128与umc1278之间,遗传距离分别为1.0,2.5 cM;等位性鉴定发现,K718d与110K(br1)组配的F1群体株高表现为矮秆,表明d718与br1等位。后续将进一步对d718进行精细定位和克隆等研究,为育种应用提供技术支撑。
