柑橘果实大小与质量的遗传分析和数量性状位点定位
2022-01-08罗艾龚桂芝彭祝春杨程常珍珍洪棋斌
罗艾,龚桂芝,彭祝春,杨程,常珍珍,洪棋斌
(西南大学柑桔研究所/中国农业科学院柑桔研究所,重庆 400712)
柑橘(CitrusL.)属于芸香科(Rutaceae)柑橘亚科(Aurantiodeae),在我国的栽种历史久远,栽培种类众多,同时也是国内栽培面积和产量最大的水果,具有重要的经济价值[1-2]。柑橘因富含维生素C、类胡萝卜素和黄酮类化合物等功能成分而具有较高的健康属性,且储藏运输方便、加工特性突出,是全世界重要的水果商品,其生产和消费量一直在不断增加[3]。近年来,随着社会经济的快速增长和消费水平的提高,人们对柑橘产品的要求也在不断改变和提高,对柑橘育种提出了更高要求。但柑橘是木本植物,具有树体大、童期长、遗传高度杂合等生物学特性,再加上一些品种的无融合生殖特性、雌性败育和/或雄性败育的存在,影响了研究者对其性状,尤其是数量性状的遗传研究,极大阻碍了柑橘育种产业发展[4-5]。分子标记辅助选择(marker-assisted selection, MAS)以及转基因育种(genetically modified breeding, GMB)技术在植物研究中的广泛应用,为加快柑橘等长周期的果树育种带来了希望[6]。针对柑橘的品质和产量等重要农艺性状开展研究,发掘其相关的调控基因或相关标记,成为利用新技术提高柑橘育种效率的重要基础。
果实大小是柑橘的重要农艺性状,不仅影响柑橘产量和外观品质,也影响生产成本和消费者的选择,过小的果实采摘成本高,过大的果实消费者一次性食用不完,影响消费者的购买欲望,因此,日本柑橘育种者在宽皮柑橘育种中,提出了果实大小介于150~300 g 的育种目标[7]。果实大小相关性状是由多基因控制的数量性状,受到内外多种因素的影响,研究难度不小,目前关于其遗传调控的机制尚不清楚,更缺乏可供育种利用的调控基因或分子标记。因此,本研究基于培育多年的‘晚蜜2号’和‘梨橙2号’杂交的F1代分离群体,借助果实大小和质量的数量性状位点(quantitative trait locus,QTL)定位,发掘相关的调控基因或相关标记,进而为柑橘果实大小和质量的遗传改良提供分子辅助育种工具和理论参考。
1 材料与方法
1.1 试验材料
分离群体是以‘晚蜜2号’[Citrus unshiu(Mark.)Marc.×C.sinensis(L.)Osb.]为母本、‘梨橙2 号’[C.sinensis(L.)Osb.]为父本杂交所得到的94株F1代材料。‘晚蜜2 号’为尾张温州蜜柑和细叶薄皮甜橙杂交所得,较抗寒,雄性部分败育,果实外观呈扁圆形,果皮橙红,种子数目较少,单胚。‘梨橙2号’为甜橙品种,果实外观呈梨形,果皮橙色,种子少,单胚。
分离群体材料和亲本均采用飞龙枳作砧木,区组重复,2012年定植在中国农业科学院柑桔研究所育种圃砧木比较试验园,到2021年完成测试时已是11 年生树,但仍有部分材料未正常结果,本研究选择结果较多的材料,总计61 株(2 株亲本和59 株杂交后代)进行果实性状的观测。
1.2 试验方法
1.2.1 分子标记分析和遗传图谱的构建
提取分离群体和亲本的DNA 以开展分子标记检测。DNA提取、保守同源序列(conserved ortholog sequence, COS)和微卫星标记即简单重复序列(simple sequence repeat, SSR)分析参考王炯[8]的方法。采用JoinMap®4.0 软件[9]进行分离群体的遗传连锁分析。
1.2.2 柑橘果实大小和质量测定及遗传分析
于2020 年柑橘果实成熟期,每株选取6 个果实,分别使用电子天平和游标卡尺对单果质量、纵横径进行测量。每个果实重复测定3 次,最后取平均值作为后续试验分析。亲本单果质量和纵横径的遗传传递力(Ta)计算公式[10]为:Ta=杂交后代性状均值/双亲性状均值×100%。
1.2.3 数量性状位点(QTL)分析
利用SPSS 22.0 软件对果实纵横径和质量数据进行统计分析,绘制频率分布直方图。基于构建的连锁图谱,采用MapQTL®6.0[11]软件对果实的单果质量和纵横径进行QTL定位分析。
2 结果与分析
2.1 柑橘遗传连锁图谱的构建结果
本试验中,杂交亲本均为遗传学上高度杂合的材料,符合JoinMap®4.0软件的“Cross Pollinator,CP”模式,CP模式杂交后代会表现出5种分离类型:ab×cd、ef×eg、hk×hk、lm×ll、nn×np。对本试验中杂交分离群体的电泳结果进行解读分析,可以比较准确地辨识出后4种分离类型,但未发现“ab×cd”类型。不同分离类型的代表性样品电泳结果如图1所示。

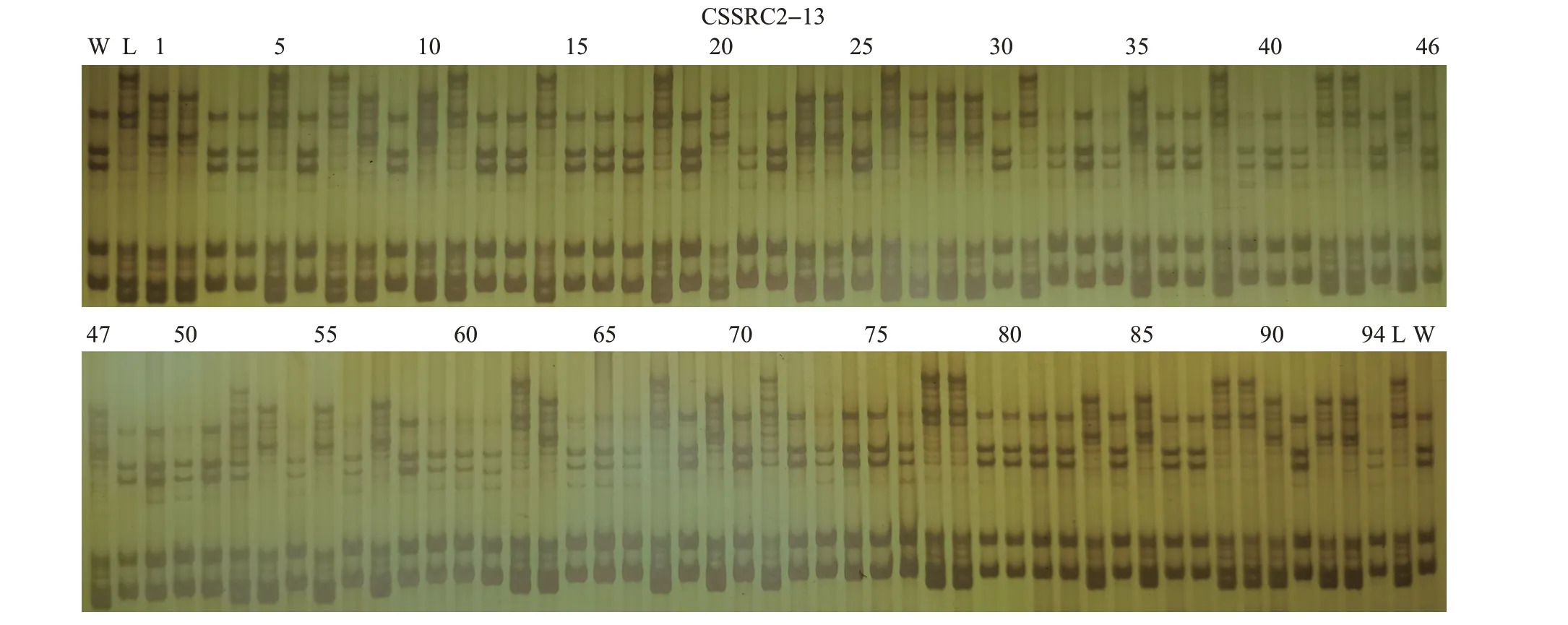
图1 不同分离类型的代表性扩增检测结果Fig.1 Representative amplification testing results of different segregation types
研究发现,在所有标记中共有108个(约54%)标记表现出明显的偏分离现象,而且大多偏向母本(P>0.05,表1)。标记偏分离比率大于CURTOLO等[12]和王炯[8]构建的柑橘遗传连锁图谱的偏分离比率(43%和45%,P=0.05),小于DE OLIVEIRA等[13]构建的柑橘连锁图谱中的偏分离比率(61%,P=0.05)。
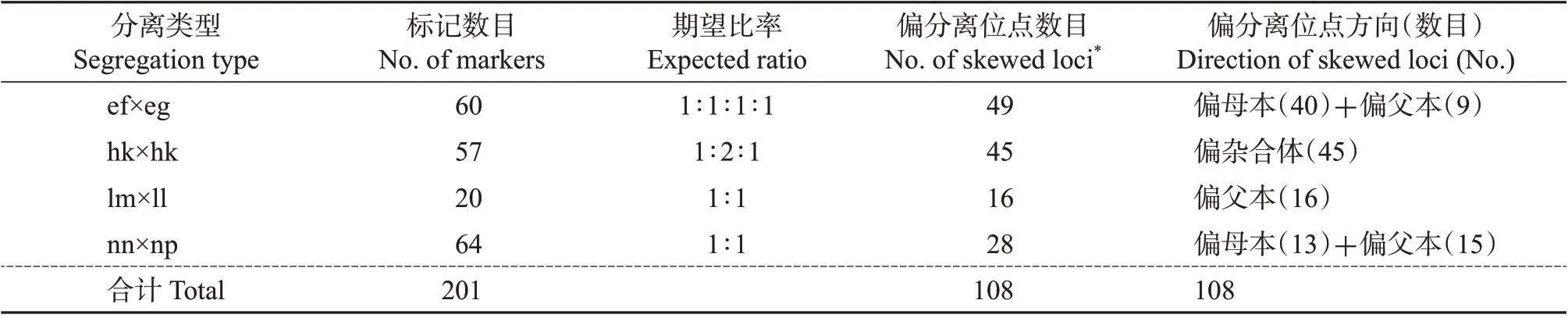
表1 标记的分离类型Table 1 Segregation types of all markers
采用比较严格的作图标准似然函数比值的对数[logarithm (base) of odds, LOD]构建遗传连锁图谱(LOD=6),并将少数未入群的标记按“最强关联组(strong cross linkage group,SCL-Group)”进行归群后,得到本研究的柑橘遗传连锁图谱。该图谱包含201 个COS 和SSR 标记、10 个连锁群,总长度为1 194.5 cM,标记间平均距离约为5.9 cM。分析发现,连锁群的主位标记与测序C.clementina(克里迈丁)的Scaffold顺序号具有一致性。以亲本‘晚蜜2号’和‘梨橙2 号’的拼音首字母缩写及测序C.clementina的Scaffold 顺序号将连锁群命名为WL1~WL9;而COSC9 系列标记在标准作图连锁过程中不能自然整合到同一连锁群,而是分成2个连锁群,本文将其分别命名为WL9-1和WL9-2(图2)。
通过与其他已发表的柑橘遗传连锁图谱[3,7-8,12]进行对比,发现分离群体、标记类型及数量对构建的图谱影响很大。RUIZ 等[14]利用随机扩增多态性DNA(random amplified polymorphic DNA,RAPD)、限制性片段长度多态性(restriction fragment length polymorphism, RFLP)和其他一些分子标记技术构建了柑橘遗传图谱,但其所构建的连锁群数与柑橘染色体数目不一致,同时还存在图距大、基因组覆盖率低等问题。近年来,研究者希望采用单核苷酸多态性(single nucleotide polymorphism, SNP)标记构建高密度遗传连锁图,也同样出现了连锁群数与染色体数不一致的情况,而且还存在分型单一、后续分析验证较难等问题。王炯[8]在2017年构建了含240个COS和SSR标记、9个连锁群,总长为1 186.4 cM的遗传连锁图谱,但其连锁群1的密度很低。本研究采用其反交材料构建图谱,构建了一张包含201个标记、10个连锁群,总长为1 194.5 cM的柑橘遗传连锁图谱,增加了连锁群1的密度,其中多数相同的标记仍然定位到同一连锁群上,少数标记在连锁群上的排列顺序有差异或者定位到了不同的连锁群上(图2)。

图2 基于COS和SSR标记构建的连锁图谱Fig.2 Linkage maps constructed by COS and SSR markers
2.2 柑橘果实大小和质量的相关性及遗传分析
对分离后代的果实质量、横纵径的均值进行比较,发现其均低于亲中值和亲本值,表现为趋小变异。亲本果实大小和质量的遗传传递力由大到小排列为横径>纵径>果实质量,其中果实质量的遗传传递力为74%,横径的遗传传递力为93%(表2)。这3 个性状的变异大,均出现明显高于或低于亲本个体的情况,表明控制柑橘果实大小和质量的增减效基因在双亲中分散分布,因此,可以通过基因重组等方式培育超亲后代。
对测试的各性状进行相关性分析发现,果实质量与横纵径均呈极显著正相关,相关系数分别为0.824 和0.835,说明果实质量受纵径影响更大。横径和纵径也呈极显著正相关,相关系数为0.756(表3)。该结果与前人研究报道[12]一致。
2.3 柑橘果实大小和质量的数量性状位点(QTL)定位分析
对分离群体的果实质量、横径、纵径数据进行分析,发现三者均为连续正态分布(图3),果实质量、横径和纵径的偏离度系数分别为1.04、0.27 和1.04,峰度系数分别为0.46、1.68 和0.65(表2),绝对值均低于2,因此,可以对其进行QTL定位。

图3 果实质量和横纵径在分离群体中的频率分布Fig.3 Frequency distributions of fruit mass,transverse and longitudinal diameters in the segregation populations
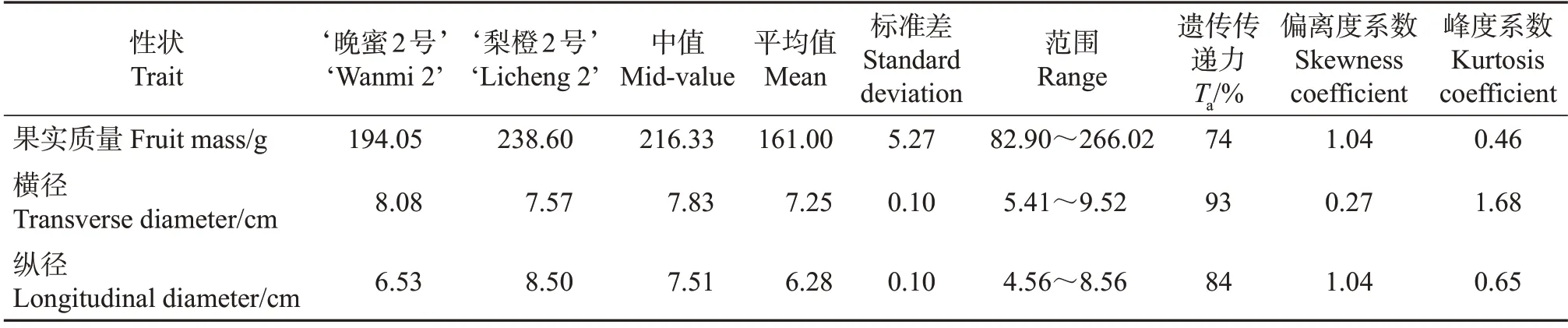
表2 亲本及分离群体的果实质量和横纵径分析Table 2 Analysis of the fruit mass,transverse and longitudinal diameters of parents and the segregation populations
基于构建的连锁图谱,采用MapQTL®6.0 软件的区间作图法对柑橘果实的质量、横径、纵径进行QTL 定位,检测到与单果质量相关的4 个QTL(LOD≥4.39),与果实横径相关的3 个QTL(LOD≥3.08),与纵径相关的4 个QTL(LOD≥3.70)(表4)。由图4 可见:这些QTLs 均位于WL3 和WL8 连锁群上,表现出集聚性;从标记界定的QTL区间来看,果实质量与纵径在连锁群WL8 上有重叠的基因组区间(CSSRC9-18~COSC8-6),也表现出一致性。这可能与果实质量、横径、纵径三者之间具有极显著正相关关系(表3)有关。
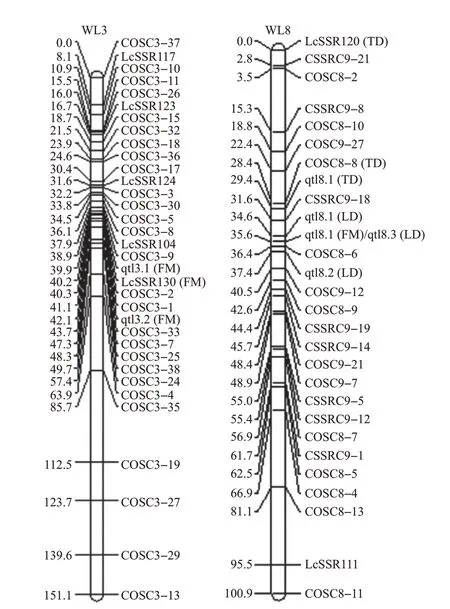
图4 柑橘果实大小和质量QTLs在连锁图谱上的定位Fig.4 QTLs locations of citrus fruit size and mass in the linkage map

表3 分离群体果实大小和质量的相关性分析Table 3 Correlation analysis of fruit size and mass in the segregation populations

表4 柑橘果实大小和质量的QTLs分析Table 4 QTLs analysis of citrus fruit size and mass
2.4 柑橘果实大小和质量QTL 区域的功能基因挖掘
以QTL所在的分子标记或区间两侧的分子标记序列选择C.clementina为参考基因组,在Phytozome(https://phytozome-next.jgi.doe.gov/pz/portal.html)网站进行Blast分析,对标记所界定的基因组区间内的编码基因进行功能注释查询,初步挖掘到一批可能与柑橘果实发育有关的功能基因,部分结果如表5 所示。进一步选择LOD 值和贡献率较高,或在果实质量和纵横径QTL 定位中均出现的候选区域开展进一步的基因挖掘分析。qtl8.1(FM)的LOD 值为4.39,qtl8.1(LD)的LOD 值为3.88,均定位于CSSRC9-18~COSC8-6 界定的区域,对该区域的序列进行分析发现:在参考基因组中临近COSC8-6引物序列的上游,有注释功能为GATA 转录因子的功能基因;在临近COSC8-6 引物序列的下游,有注释功能为生长素响应因子(auxin response factor,ARF)的功能基因。GATA为一类植物转录因子,能与靶基因启动子上的WGATAR 区域结合,激活或抑制下游靶基因的转录,从而调控植物的发育[15]。ARF 是由ULMASOV 等[16]新发现的一类调控生长素响应基因表达的转录因子家族,能够特异地与生长素响应基因启动子区域的生长素响应元件结合。梅丽华[17]研究发现,番茄ARF 家族成员之一的SlARF10通过控制淀粉合成相关酶的表达来促进糖类物质的积累,在番茄果实发育过程中发挥着重要作用。说明ARF 可能具有一因多效的性质,对果实质量、横纵径有共同调控作用。本试验中,LcSSR130(FM)的LOD 值为5.88,LcSSR130 是基于表达序列标签(expressed sequence tag,EST)开发的引物,该EST 所在的基因功能注释为GDSL 酯酶(GDSL esterase)。有研究报道,来自水稻脆叶鞘1突变体BS1的GDSL 酯酶成员是一种次级壁乙酰-木聚糖酯酶,能调控木聚糖的脱乙酰化,进而导致次生壁发育受损和异常的多变性表型[18]。柑橘果实中的GDSL酯酶成员也很可能通过动态微调多糖的乙酰化结构来发挥结构调控作用,从而调节细胞壁结构,进而对果实的大小和质量产生影响。
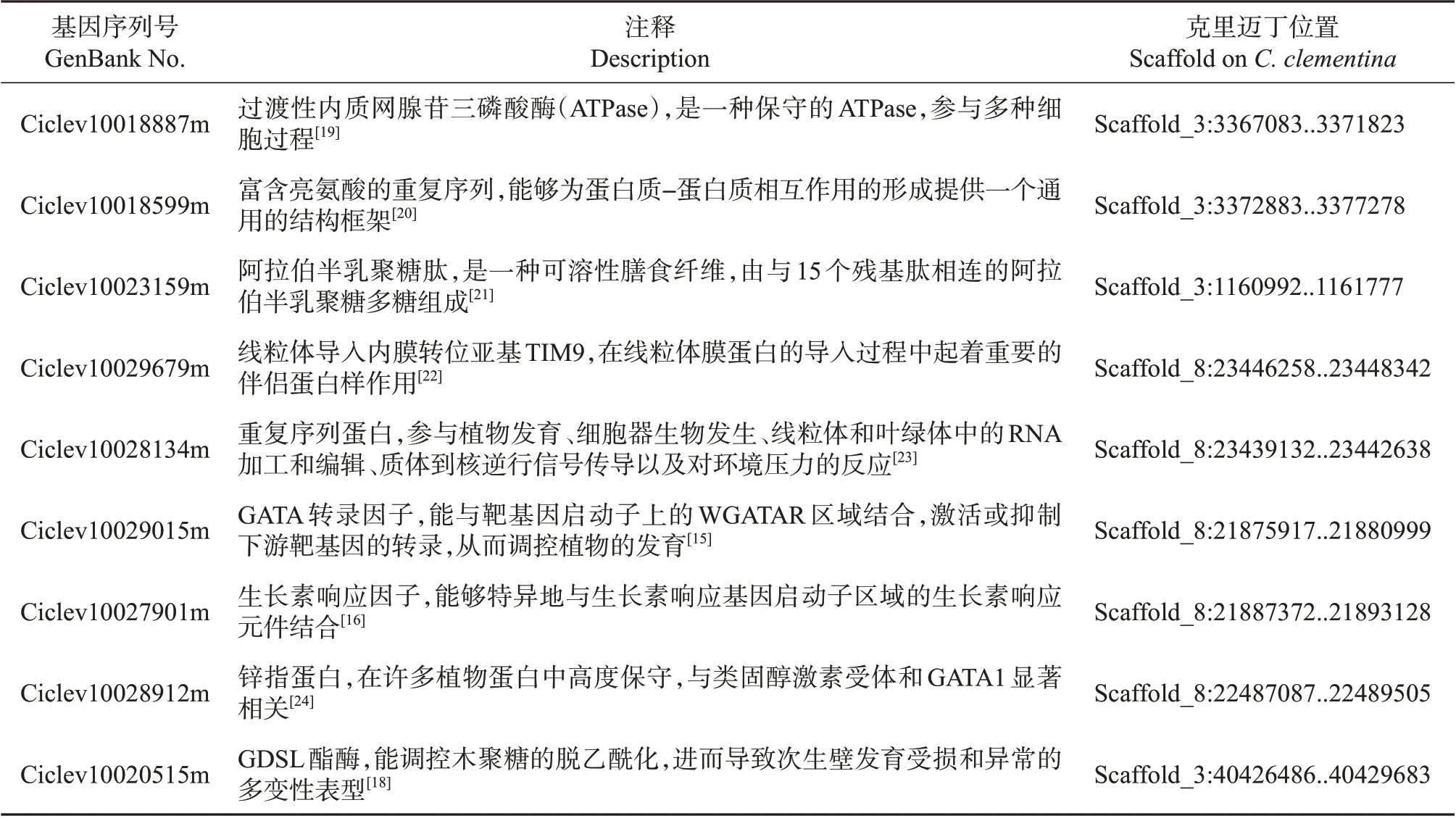
表5 与柑橘果实发育可能相关的基因Table 5 Genes putatively associated with citrus fruit development
3 讨论与结论
3.1 偏分离标记的作图应用
孟德尔分离定律是经典遗传学的基本原则,能帮助遗传学家预测简单遗传性状的表达情况。人们从20 世纪初就开始研究偏离孟德尔分离预期的现象,即偏分离现象,主要是配子或合子选择、种间或种内杂交引起染色体重排而导致的。在物种的大多数遗传作图研究中都观察到了偏分离标记[8,10],其可能会使标记之间遗传距离的估计出现偏差,并可能影响标记之间的顺序,从而影响表型性状QTL定位的准确性。因此,在遗传作图中常有将偏分离标记舍弃后再进行图谱构建的报道[3,7]。DE OLIVEIRA等[25]通过对仅包含正常分离标记图谱和包含偏分离标记的图谱进行比较发现,偏分离标记一般位于染色体的末端部位,并未显著改变标记的距离。同时,随着对偏分离的深入研究,人们发现偏分离标记对QTL定位中的加性效应影响较小,并且偏分离标记可能与含生物关键遗传特性的染色体区域相关[26],如果将其排除在遗传图谱之外,则无法检测到一些与生长和成熟相关的生物学功能。偏分离标记大部分是聚集的,并且每个聚集内的标记具有相同的偏斜方向,比如多数位点偏向于同一亲本或杂合体,这表明偏分离标记是由生物因素引起的[27]。本研究的偏分离标记大多偏向于母本,说明来自母本基因型的配子在形成合子时占主导地位。同时,本研究所定位到的QTL 相关的分子标记,比如LcSSR130(FM)、LcSSR120(TD)和COSC8-8(TD)等大多是偏分离标记,因此将其用于图谱的构建必不可少。目前,已有关于偏分离分析模型与软件的报道,说明大部分的偏分离标记确实可以用于遗传图谱构建,只是需根据实际情况进行评估和调整[27]。
3.2 柑橘果实大小和质量的QTL 比较
本研究采用的COS 标记是根据测序克里迈丁从头(de novo)组装结果(https://phytozome.jgi.doe.gov/pz/portal.html)开发的。因此,得到的连锁群与克里迈丁参考基因组的染色体顺序具有一致性,可以比较全面地与其他研究者基于一致的连锁群定义而开展的柑橘果实大小和质量的QTL 定位进行比较分析。以克里迈丁为参考基因组,GARCÍA等[28]检测到与平均果实大小相关的3个QTLs,分别分布在V2、V7、V8 连锁群上;检测到与平均单果质量相关的2个QTLs,分布在V2和V7连锁群上。但其报道中并未表明这些位点的LOD 值和贡献率。YU 等[3]结合‘Fortune’בMurcott’(Citrus reticulataBlanco)的F1代检测到3 个与柑橘横径相关的QTLs,3 个与单果质量相关的QTLs,这些位点分布在MUR4.2、MUR8、FOR5.1和FOR9.3连锁群上,其中只有位于连锁群FOR5.1 上的FM5.1 和位于MUR8 上的FM8 在2012 年和2013 年均有被检测到。IMAI 等[7]定位到4 个与柑橘单果质量相关的QTLs,分布在LG3、LG4和LG7连锁群上,其中只有位于LG4 上的位点FMq3 在连续2 年里被检测到。本研究定位到的与果实大小相关的QTLs 分布在WL3和WL8连锁群上。综上所述,在柑橘第3、4和8染色体上可能存在控制果实大小的QTLs,而在第8 染色体上可能存在控制柑橘果实大小的相关基因,在第3 染色体上可能主要存在与果实质量相关的功能基因。
YU等[3]和IMAI等[7]的研究中,与果实质量和横径相关的QTLs 的LOD 值在2.69~3.68 之间,贡献率在14.9%~33.74%之间;本研究中与果实大小和质量相关的QTLs 的LOD 值在3.08~5.94 之间,贡献率在19.6%~34.4%之间,研究结果与前人相近。CURTOLO 等[12]研究发现,与柑橘果实横纵径相关的QTLs 的LOD 值虽然都大于5.94,但是其表型变异率最高只有9.7%;GARCÍA 等[28]却没有报道其定位到的QTLs 的LOD 值和解释表型变异的决定系数,因此,其定位到的QTLs还有待考证。BUDAHN等[29]研究表明,超过45%的表型变异是由主要的QTLs 效应造成的,而这在本文和以上研究中均没有观察到。但也有报道指出贡献率大于30%即可认为是主效QTLs[30]。因此,对于主效QTLs 的判断标准还有待进一步探讨,但这也说明果实大小和质量是由多基因共同控制的。
基因型×环境互作效应对柑橘果实品质性状起着重要作用,基因型×取样日期对所有柑橘果实品质性状也有显著影响,特别是柑橘的外部特征比内部特征波动更大。YU等[3]使用2年的表型数据检测到与果实直径和果实质量相关的QTLs有6个,在该研究中,2年里取样时间和表型材料并不完全相同,所以只有关于果实质量的2 个QTLs 在2 年里被检测到,而与果实直径相关的QTLs 在2 年里均不相同。IMAI等[7]采用3年的表型数据定位到与果实质量相关的QTLs 位点有4 个,且只有1 个位点在2 年里被连续检测到。CURTOLO 等[12]结合1 年的表型数据,只检测到3个与果实纵横径相关的QTLs。与以上研究比较,我们对分离群体的果实发育全程进行了分析测试,发现到发育后期,即开花后246—274 d,果实的生长量变化很小,果实大小总体已进入比较稳定的状态;此外,本研究仅选用果实发育停滞时段的测试数据进行分析,减少了数据波动对QTL 定位的影响。本研究定位到了与果实大小相关的10个QTLs,数量较多,这可能与我们所研究的分离群体果实大小和质量分离比较大有关,如在本研究中单果质量的变化范围为82.90~266.02 g,变异范围较大。分离群体中尚有部分未结果,连锁图谱存在标记数目不足,连锁群间和群内分布不均匀等,都对本研究的QTL数量确定和精准鉴定等带来了较大影响。后续可通过开发和应用更多分子标记优化遗传图谱,对更多结果个体进行分析以得到更好的QTL 鉴定结果。在对LOD 值和贡献率较高,或在果实质量和纵横径QTL定位中均出现的候选区域进行基因挖掘时,发现本研究所定位到的QTL 所在的基因组区间确实存在对果实发育和大小产生影响的基因,这些基因对柑橘果实大小和质量的遗传和育种的深入研究和应用探索具有一定的参考价值。
