黔北黑猪肌内脂肪沉积相关基因的筛选
2022-01-08孟金柱吴震洋安清明赵园园
孟金柱,吴震洋,安清明,赵园园*
(1.铜仁学院贵州省梵净山地区生物多样性保护与利用重点实验室,贵州铜仁 554300;2.湖南农业大学动物医学院,长沙 410128)
猪肉的品质受多方面因素的影响,如品种、营养、屠宰时的应激程度、猪肉的保鲜方式等,而猪肉本身的理化特性是决定其品质的根本因素。在很大程度上,猪肉的肌纤维类型及组成决定其理化特性,并进一步影响其品质[1]。肌肉由肌细胞组成,由于肌细胞细长,呈纤维状,因此也称为肌纤维。肌纤维由肌膜、肌浆和肌丝等构成。肌肉收缩是细肌丝和粗肌丝的相对滑动导致的,由2 条重链和多条轻链构成的肌球蛋白在该过程中承担着分子马达的作用[2-3]。哺乳动物的肌纤维根据肌球蛋白重链(myosin heavy chain, MyHC)异构体的不同分为4种类型,即慢速氧化型(Ⅰ型)、快速氧化型(ⅡA型)、中间型(ⅡX 型)和快速酵解型(ⅡB 型)。其中:Ⅰ型肌纤维因含有较多的肌红蛋白而颜色红润,同时也含有较多的线粒体,主要进行有氧代谢,为慢氧化纤维,直径较小。ⅡB型肌纤维颜色苍白,因糖原和腺苷三磷酸(adenosine triphosphate,ATP)酶系含量较高,主要依靠糖酵解供能,直径较大。ⅡA和ⅡX型肌纤维的理化特性则介于上述两者之间[4]。
肌肉中肌纤维的组成和类型对肉品质都有很大的影响,如:肌纤维横截面积越大,肉质性状越差[5];肌肉中ⅡB型肌纤维所占比例较高时,pH下降较快,易产生白肌(pale,soft and exudative,PSE)肉,而Ⅰ型肌纤维的比例与pH 呈正相关[6]。肉色与肌纤维中的肌红蛋白含量直接相关,Ⅰ型肌纤维中肌红蛋白含量最高,又被称为红肌纤维,而ⅡB型肌纤维颜色苍白,又被称为白肌纤维,因此,Ⅰ型肌纤维所占比例较高时,肉质红润,肉色评分较高[7]。肌纤维直径影响肌肉嫩度,ⅡB 型肌纤维直径和横截面积较大,在肌肉中所占比例较高时,肌肉的剪切力增加,嫩度降低[8]。肌内脂肪含量与Ⅰ型肌纤维的含量呈正相关[9],原因在于Ⅰ型肌纤维包含更多的中性脂质,同时,Ⅰ型肌纤维中含有的高水平的磷脂质与肉的多汁性呈正相关[10]。
黔北黑猪主要生长在贵州省北部,具有性成熟早、繁殖力强、耐热耐寒、抗病力强、储脂能力强、肉质好的特性[11],但由于其生长速度缓慢,经济效益低,近年来数量锐减。猪背最长肌(longissimus dorsi muscle,LDM)主要由ⅡB型肌纤维构成,腰大肌(psoas major muscle,PMM)则由较多的ⅡA 型肌纤维和ⅡX 型肌纤维及少量的ⅡB 型肌纤维构成,相比较而言,PMM 具有更好的肉品质,肉色鲜红,脂肪含量高[10]。本研究通过检测黔北黑猪LDM 和PMM中脂肪含量,结合高通量测序技术检测这2种肌肉中的差异表达基因,并利用生物信息学技术对这些基因的功能进行分析,拟筛选出与肌内脂肪沉积相关的关键基因并加以验证,旨在揭示黔北黑猪肌内脂肪沉积的分子机制,为黔北黑猪的保护、开发和利用提供实验依据。
1 材料与方法
1.1 实验动物及样品采集
从贵州省德江县农祥牧业发展有限公司选取5头健康的12 月龄黔北黑猪,屠宰后分别取2 cm×1 cm×1 cm 大小的位于最后肋附近的LDM 和倒数第4 肋附近的PMM 各5 块,放入1.5 mL 离心管,然后置于液氮中进行冷冻保存。本研究中所涉及的动物实验都遵循国家有关法律和法规中规定的动物福利和伦理原则。
1.2 实验方法
1.2.1 肌肉组织中脂肪含量测定
采用索氏抽提法,称取5 g肉样放入滤纸筒,然后将滤纸筒放入索氏抽提器的抽提筒内,连接已干燥至恒量的接收瓶,从抽提器冷凝管上端加入无水乙醚,水浴加热,使无水乙醚不断回流抽提10 h。取下接收瓶,回收无水乙醚,待接收瓶内溶剂剩余1~2 mL时在水浴锅上蒸干,再于(100±5)℃条件下干燥1 h,放入干燥器内冷却0.5 h 后称量。重复以上操作直至恒量,计算脂肪含量。
1.2.2 总RNA 提取、纯化、建库和测序
黔北黑猪LDM 和PMM 中的总RNA 采用购自美国Invitrogen 公司的Trizol(货号:15596018)提取,随后用购自德国Qiagen 公司的RNeasy Mini 试剂盒(货号:74104)纯化,利用琼脂糖凝胶电泳检测RNA的完整性,最后,采用购自美国Thermo公司的NanoDrop 2000分光光度计测量其浓度。每个样本取3 μg 总RNA 用于建库和测序(n=5),委托北京诺禾致源生物信息科技有限公司完成,其中测序采用Illumina Hiseq 2500平台进行。
1.2.3 数据处理及分析
测序获得的原始数据依次通过FastQC v0.10.8软件进行质量评估并获得有效读长(clean reads),经Trinity v0.1.0-alpha 软件重新组装后,得到重叠群。使用CLC Genomics Workbench v21.0.2 软件将重叠群与猪RefSeq 数据库进行比对,获得注释基因。利用DESeq2 1.26.0 软件分析差异表达的mRNA,并用GOseq 1.15.5 软件和KOBAS 3.0 软件对差异表达的mRNA 进行基因本体(gene ontology,GO)以及京都基因和基因组百科全书(Kyoto encyclopedia of genes and genomes,KEGG)信号通路分析。
1.2.4 脂肪沉积相关基因的筛选
根据GeneCards 数据库(https://www.genecards.org/)和NCBI网站(https://www.ncbi.nlm.nih.gov/)中对差异表达的mRNA功能的报道,筛选脂肪沉积相关基因。
1.2.5 实时荧光定量聚合酶链反应(quantitative real-time polymerase chain reaction,qRT-PCR)验证
为验证高通量测序结果的准确性,对1.2.2节提取的总RNA 进行反转录,合成cDNA,然后运用NCBI 网站(https://www.ncbi.nlm.nih.gov/)中的Pick Primer程序设计筛选的脂肪沉积相关基因的特异性引物(表1),交由华大基因公司合成。qRT-PCR 采用北京全式金生物技术有限公司的EasyScript®Allin-One First-Strand cDNA Synthesis SuperMix for qPCR(One-Step gDNA Removal)试剂盒(货号:AE341-02)建立20 μL 反应体系:2×TransStart Tip Green qPCR SuperMix 10 μL,上、下游引物(8 pmol)共0.8 μL,cDNA(100 ng)4 μL 以及ddH2O 5.2 μL。反应设置5个样本重复,4个技术重复,通过美国罗氏Light Cycler 480 荧光定量仪进行反应。以β-Actin作为内参基因,所有基因的表达水平用2-ΔΔCT法计算,并使用SPSS 17.0软件进行差异显著性分析。

表1 供试引物序列Table 1 Tested primer sequences
2 结果与分析
2.1 黔北黑猪LDM 和PMM 的表型及脂肪含量
通过索氏抽提法检测黔北黑猪LDM 和PMM中的总脂肪含量。结果表明,PMM 中的总脂肪含量极显著高于LDM(P<0.01),LDM 的颜色较为明亮,而PMM呈暗红色(图1)。
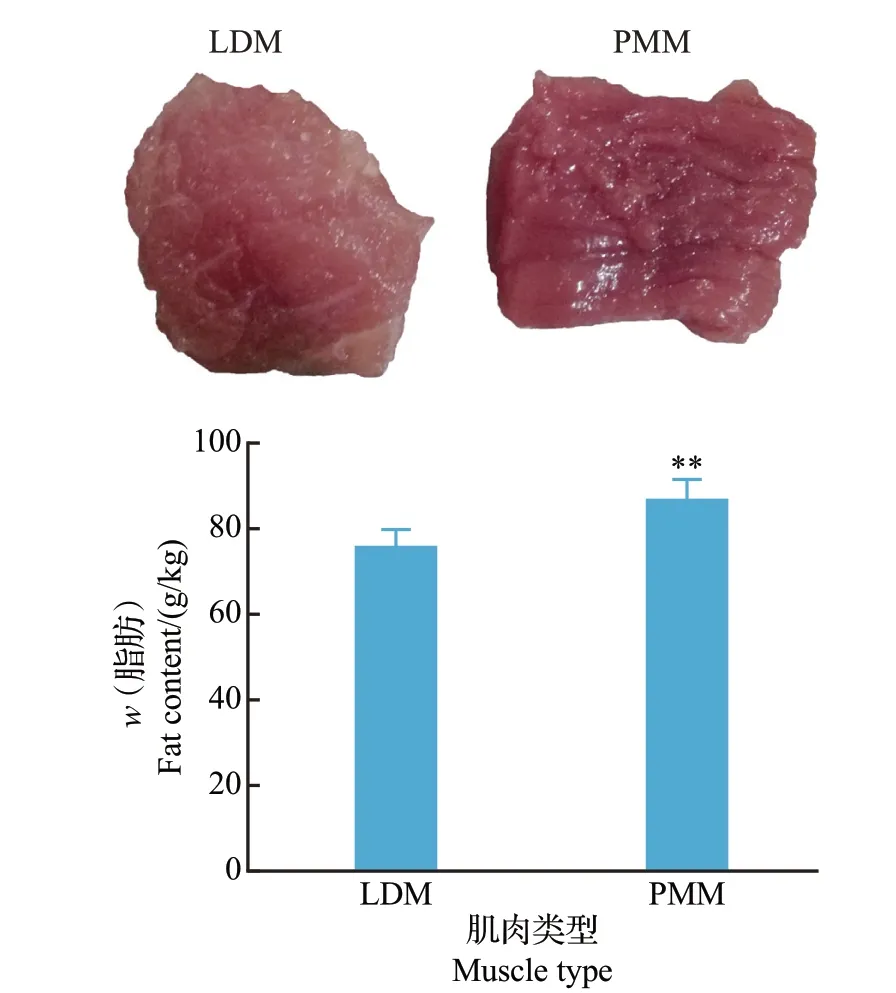
图1 黔北黑猪LDM和PMM的表型及脂肪含量Fig.1 Phenotypic traits and fat contents of LDM and PMM of Qianbei black pigs
2.2 黔北黑猪LDM 和PMM 中的差异表达基因
对高通量测序数据进行去杂、注释和差异表达分析后,按每千个碱基的转录中每百万映射读取的片段数(fragments per kilobase of transcript per million mapped reads, FPKM)>0.5,|log2(差异倍数)|>1 及校正后的P值(q值)<0.05 的标准进行筛选,共获得162 个差异表达基因(附表1,http://www.zjujournals.com/agr/CN/10.3785/j.issn.1008-9209.2021.01.191),其中96个基因表达下调,66个基因表达上调(图2)。

图2 黔北黑猪LDM和PMM差异表达基因火山图Fig.2 Volcano plot of differentially expressed genes in LDM and PMM of Qianbei black pigs
2.3 黔北黑猪LDM 和PMM 中差异表达基因的GO 功能富集
对162个差异表达基因进行GO功能富集分析,结果共分为3 大类,其中,生物学过程(biological process)11组,细胞组分(cellular component)7组,分子功能(molecular function)8 组。在生物学过程中富集最多基因的是心脏肌肉收缩和细胞钙离子稳态,在细胞组分中富集最多基因的是胞质和线粒体内膜,在分子功能中富集最多基因的是运输活性和血红素结合(图3)。

图3 黔北黑猪LDM与PMM差异表达基因GO分析Fig.3 GO analysis of differentially expressed genes between LDM and PMM of Qianbei black pigs
2.4 黔北黑猪LDM 和PMM 中差异表达基因的KEGG 信号通路分析
对差异表达基因进行KEGG信号通路分析,结果发现14 条信号通路(图4),其中参与基因最多的是代谢途径(15.54%),肌内脂肪沉积相关的通路包括过氧化物酶体增殖物激活受体(peroxisome proliferator-activated receptor, PPAR)信号通路(6.76%)、脂肪酸代谢(4.05%)、脂肪酸降解(3.38%)、脂肪细胞因子信号通路(3.38%)、非酒精性脂肪肝病(4.05%),参与这些通路的基因包括SLC27A1、FABP3、CPT2、ACSL1、SCD、LPL、NR1H3、PPARA、CPT1B、RXRG、ACADVL、ACAA2、PRKAG2、COX2、COX1、SDHD。

图4 黔北黑猪LDM与PMM差异表达基因KEGG通路分析Fig.4 KEGG pathway analysis of differentially expressed genes between LDM and PPM of Qianbei black pigs
2.5 qRT-PCR 验证分析
利用NCBI 和GeneCards 数据库对上述脂肪沉积相关基因的功能进行分析,结果筛选出6 个脂肪沉积相关基因,即LPL、SDHD、PPARA、COX2、PRKAG2和SCD。通过qRT-PCR对这6个基因表达水平进行检测,以验证高通量测序结果的准确性。结果(图5)表明,这6个基因的表达趋势跟高通量测序结果一致:LPL、PPARA和COX2在LDM 中的表达量显著或极显著低于PMM(P<0.05或P<0.01),SDHD的表达量也呈现相同的趋势,但差异不显著;PRKAG2和SCD在LDM 中的表达量显著或极显著高于PMM(P<0.05或P<0.01)。

图5 肌内脂肪沉积相关基因在黔北黑猪LDM与PMM中的相对表达情况Fig.5 Relative expression levels of intramuscular fat deposition related genes in LDM and PMM of Qianbei black pigs
3 讨论
与大白猪和长白猪等引进品种相比,我国本地猪种肉质好,主要表现为颜色鲜红、无PSE肉,保水力好,肌纤维直径较低、密度大,肌内脂肪含量高[12]。肌内脂肪含量是猪肉嫩度、大理石纹、多汁性、风味等的重要物质基础,并且肌内脂肪含量与肌纤维类型和组成紧密相关。Ⅰ型肌纤维中脂肪含量是ⅡB型肌纤维的3倍[13]。以Ⅰ型肌纤维为主的半膜肌比以ⅡB型肌纤维为主的背最长肌含有更高水平的总脂肪、甘油三酯和磷酸[14]。本研究中,对LDM 和PMM 的脂肪含量进行检测也发现,PMM 中的脂肪含量极显著高于LDM。这可能是由于PMM由较多的ⅡA型肌纤维和ⅡX型肌纤维及少量的ⅡB型肌纤维构成,而LDM主要由ⅡB型肌纤维组成。
石岗[12]对深县猪和杜洛克猪的LDM 进行高通量测序,结果获得282个差异表达基因,其中170个上调,112个下调。王志秀[15]对滇南小耳猪、藏猪、长白猪和杜洛克猪进行高通量测序,结果在滇南小耳猪-藏猪组和长白猪-杜洛克猪组中共筛选出差异表达基因315 个,其中上调基因140 个,下调基因175个,并鉴定出27个与脂肪沉积相关的基因,主要参与脂肪代谢、脂肪酸合成等过程。庞喆[16]对高脂肪组和低脂肪组延边黄牛的LDM 进行高通量测序,结果共获得38 个差异表达基因,其中上调基因有25个,下调基因有13个。石斌刚[17]在对不同月龄天祝白牦牛LDM的高通量测序结果中发现10个脂肪沉积相关的候选基因,其中4 个上调,6 个下调。本研究运用高通量测序技术和生物信息学技术,在黔北黑猪LDM 和PMM 中共发现162 个差异表达基因,其中96 个基因表达下调,66 个基因表达上调,最终筛选出6 个与脂肪沉积相关的候选基因。qRT-PCR 结果表明,LPL、PPARA、COX2在LDM 中的表达量显著或极显著低于PMM(P<0.05 或P<0.01);PRKAG2和SCD在LDM 中的表达量显著或极显著高于PMM(P<0.05或P<0.01)。
脂蛋白脂肪酶(lipoprotein lipase, LPL)又被称为清除因子脂肪酶[18]。研究表明,在猪中LPLmRNA 表达量与肌内脂肪含量呈不同程度的正相关[19],其通过水解脂蛋白中的甘油三酯,为组织利用和储存提供游离脂肪酸,同时,通过调控肌肉和脂肪中的血浆甘油三酯的表达来决定脂肪沉积的程度[20]。过氧化物酶体增殖物激活受体α(peroxisome proliferator-activated receptor alpha, PPARA)通过抑制1-氨基环丙烷-1-羧酸(1-aminocyclopropane-1-carboxylic acid,ACC)、肿瘤坏死因子受体超家族成员6(tumor necrosis factor receptor superfamily member 6,TNFRSF6/FAS)、硬脂酰辅酶A去饱和酶(stearoyl-coenzyme A desaturase, SCD)等脂肪酸从头合成基因的表达,从而抑制甘油三酯的合成,防止甘油三酯的积累[21]。活化后的PPARA,通过调节脂联素的活性使胰岛素的敏感性增强,进一步促进脂肪酸氧化酶mRNA的表达,最终促进脂质分解代谢[22]。蛋白激酶腺嘌呤核糖核苷酸激活的非催化亚基γ2(protein kinase AMP-activated non-catalytic subunit gamma 2,PRKAG2)作为腺嘌呤核糖核苷酸活化蛋白激酶(AMP-activated protein kinase,AMPK)家族重要的成员,在肌肉脂肪代谢、能量代谢等方面发挥着巨大作用[23]。JING 等[24]通过对高、低剩余采食量的猪骨骼肌进行高通量测序发现,PRKAG2 在骨骼肌能量代谢过程中起着至关重要的作用。SCD 在动物肌肉组织中大量表达并参与单不饱和脂肪酸的形成,SCD基因的过表达能够促进甘油三酯和脂肪酸的合成[25]。钙离子浓度对脂质含量具有一定的调节作用,其浓度的升高会促进SCD基因的表达,使脂肪酸的合成量进一步增加[26]。
4 结论
本研究通过检测黔北黑猪LDM 与PMM 中的总脂肪含量,发现PMM 中的脂肪含量极显著高于LDM。利用高通量测序在2种肌肉中获得162个差异表达基因。通过GO、KEGG 分析并结合功能查找,共筛选出6个与脂肪沉积相关的差异表达基因,其中LPL、PPARA、COX2、PRKAG2和SCD在LDM和PMM 中的表达情况存在显著差异,推测这些基因能够促进或抑制肌内脂肪沉积。本研究结果为深入探究黔北黑猪肌内脂肪沉积的分子机制以及对黔北黑猪的保护、开发和利用提供了实验依据。
