隔药饼灸对慢性萎缩性胃炎大鼠PTEN-AKT抑癌途径的作用机制研究
2022-01-05郑雪萧有智吴梦蝶吴焕淦马喆王文佳黄艳李琪李璟
郑雪,萧有智,吴梦蝶,吴焕淦,马喆,王文佳,黄艳,李琪,李璟
(1.上海中医药大学,上海 201203;2.河南中医药大学第一附属医院,郑州 450000;3.上海中医药大学附属岳阳中西医结合医院,上海 200437;4.上海市针灸经络研究所,上海 200030)
慢性萎缩性胃炎(chronic atrophic gastritis,CAG)是临床上常见的一种消化系统疾病,病理表现主要为胃黏膜固有腺体的减少,伴或不伴肠上皮化生和(或)异型增生[1]。本病临床表现常轻重不一,以胃脘痛、上腹部不适、食欲不振、泛酸、饱胀、恶心呕吐等为主,伴随乏力、嗳气、抑郁、焦虑等症状[2]。流行病学研究发现,萎缩伴肠化发展为胃癌的中位时间为6.1年,低度异型增生为2.6年[3],且胃癌的发病人群主要集中在日本、中国、南美等国家[4]。积极有效地防治CAG,阻断其向胃癌发展,已成为预防胃癌的重要途径。CAG发病机制尚未明确,认为该病与幽门螺旋杆菌感染、免疫、年龄、十二指肠液返流、家族遗传史等多种因素有关[5]。CAG具有病程长、症状反复发作、迁延难愈的特点,给患者和社会造成巨大的经济负担。积极探讨其发病机制,寻求有效的治疗方法迫在眉睫。
CAG基础上伴发的肠上皮化生及不典型增生,属于胃癌前期病变,是从正常胃黏膜向胃癌转化过程中的重要阶段。磷酸酶与张力蛋白同源物(phosphatase and tension homolog deleted onchromosome ten,PTEN)是至今为止发现的首个具有脂质和蛋白磷酸酶活性的抑癌基因,其从慢性浅表性胃炎开始表达降低,在萎缩、萎缩伴肠化和异型增生至胃癌的进展过程中持续丢失[6]。研究表明,PTEN的缺失可激活PI3K/AKT信号通路,促进肿瘤的发生发展[7]。PI3K/AKT通路是一条经典的抗凋亡和促存活的信号传导通路,在介导细胞存活、促进细胞生长及增殖,抑制细胞凋亡等方面起到关键作用,该通路已被发现在多种肿瘤细胞中异常激活,与癌症的发生发展密切相关[8]。CAG是胃癌进展过程中的关键环节,积极探讨PTEN/PI3K/AKT信号通路在CAG的作用机制,逆转其发展进程对胃癌的防治具有积极意义。
目前现代医学尚缺乏有效治疗 CAG的方法,临床上主要以根除幽门螺旋杆菌(helicobacter pylori,Hp)感染、补充叶酸、保护胃黏膜、改善胃动力、消除致病因素等作为常用的治疗方法[9]。近年来,中医药尤其是针灸疗法在治疗 CAG中占据重要的地位,动物实验和临床试验均表明针灸在治疗 CAG中具有独特优势[10-11]。本课题组前期临床研究证实,艾灸能调节胃肠功能、促进胃黏膜修复、提高机体免疫[12-13]。但目前大多数研究停留在临床疗效观察阶段,艾灸治疗 CAG的作用机制尚未明确。因此,本研究通过动物实验,探讨艾灸干预 CAG的效应机制,拟证实艾灸可能通过影响CAG大鼠胃黏膜组织PTEN/PI3K/AKT通路,调控其相关基因的表达,有效控制 CAG胃黏膜损伤,达到防治CAG癌前病变的目的。
1 材料与方法
1.1 实验动物
清洁级雄性 Wistar大鼠 52只,6周龄,体质量(150±30) g,由上海斯莱克实验动物有限公司提供。动物合格证编号为 2015000538649;许可证号为SCXK(沪)2012-0002。饲养环境设置为室内湿度40%~70%,室温(18~22) ℃。实验操作均在上海中医药大学实验动物管理委员会制定的实验动物使用及保护条例的指导下进行。
1.2 药物和试剂
N-甲基-N'-硝基-N-亚硝基胍(N-methyl-N'-nitro-N-nitrosoguanidine, MNNG)(日本东京株式会社);叶酸片(常州制药厂有限公司);4%多聚甲醛和二甲苯(国药集团化学试剂有限公司);苏木素和伊红(南京建成科技有限公司);Anti-PIP2抗体(Abcam,ab11039);Anti-PTEN抗体(Abcam, ab31392);AKTPhospho-S473 抗体(proteintech, 66444-1-Ig);MDM2抗体(SantaCruz, sc-13161);Anti-Caspase-9 抗体(Abcam, ab52298)。
1.3 模型制备与鉴定
大鼠适应性饲养1周后,随机分为正常组(12只)和造模组大鼠(40只)。正常大鼠予以常规饮食,造模大鼠采用 MNNG诱导剂结合夹尾刺激与饥饱失常法制备CAG大鼠模型[14],造模周期为34周。造模大鼠每日自由饮用浓度为 170 mg/L的 MNNG溶液,每日更换 1次。饱食2 d,禁食1 d,循环实施。每周用夹尾夹夹住大鼠尾部1次,持续10 min,使之保持激怒、互相攻击的状态。34周后从造模大鼠取4只大鼠进行模型鉴定,取大鼠胃窦组织并做HE染色,光学显微镜下观察大鼠胃黏膜组织病理学变化,观察到大鼠胃黏膜出现萎缩、或/和肠上皮化生病理变化,确定模型成功。
1.4 分组与干预
至34周造模组大鼠死亡6只,取4只进行模型鉴定。造模结束余正常组大鼠12只,造模大鼠30只。剩余30只造模成功大鼠随机分为模型组、隔药饼灸组和西药组,每组10只。正常组与模型组不进行任何干预,只做同隔药灸组相同的抓取和固定;西药组予叶酸悬浊液灌胃给药(每100 g体质量给药1 mL),每日1次,1周6次,共4周;隔药饼灸组取穴中脘和气海穴,将大鼠固定于自制大鼠固定架上,用调制好的药饼(直径 1 cm、厚度 0.5 cm)放置大鼠的穴区,上置 90 mg艾炷,每穴灸2壮,每日1次,1周6次,共4周。中脘穴定位为大鼠腹白线上,脐上20 mm。气海穴定位为大鼠腹白线上,脐下 12.5 mm[15]。干预期间,模型组大鼠死亡1只,剩9只。
1.5 标本采集及处理
治疗结束后,各组大鼠禁食不禁水24 h,以10%水合氯醛溶液(每 100 g体质量注射0.3 mL)麻醉,待大鼠麻醉后腹部朝上固定在解剖板上,迅速用手术剪打开大鼠腹腔,立即摘除全胃,沿胃大弯剪开,0.9%氯化钠溶液将胃内容物洗净,肉眼观察并记录胃黏膜情况;切取病变胃黏膜组织,取1/2胃窦部组织于4%多聚甲醛溶液固定,常规石蜡包埋切片与染色;另取1/2胃窦部组织,剪碎置于冻存管内,液氮冷冻 1 h后放置于﹣80 ℃冰箱保存备用。
1.6 检测指标
1.6.1 各组大鼠胃窦组织病理学观察
将切片置于二甲苯溶液Ⅰ、Ⅱ各 10 min;下行梯度100%、90%、80%、70%乙醇溶液各5 min;自来水冲洗;苏木素浸染10 min后流水冲洗10 min,1%盐酸酒精溶液分化10 s,流水冲洗至细胞核呈蓝色;伊红溶液浸染 30 s~5 min;自来水冲洗数秒钟;上行梯度70%、80%、90%、100%乙醇溶液各1~2 s;二甲苯溶液Ⅰ、Ⅱ透明各 15 min;封片后在光学显微镜下观察各组胃黏膜病变程度。
1.6.2 各组大鼠胃窦组织 PTEN、PIP2、p-mTOR、Caspase-9、MDM2蛋白检测
取大鼠胃窦组织石蜡切片,经脱蜡、水化、抗原修复、封闭后于37 ℃条件下滴加一抗,4 ℃冰箱过夜并于第2天滴加二抗孵育;PBS溶液冲洗后用DAB显色;苏木素染色,脱水,封片。对照组分别设阳性、阴性、空白对照,用已知阳性表达组织的切片作为阳性对照组,以 PBS溶液作为空白对照组。采用 LAS Version 4.30图像采集系统进行电脑图像拍片,每张组织切片要求在固定光亮度下,随机选取3个视野进行拍照。拍片结束后,采用Image J 6.0软件进行半定量分析,可得到每张照片的平均阳性面积率(%Area),最后取3张照片的平均值即为该组织切片的阳性目标的平均阳性面积率。
1.6.3 各组大鼠胃窦组织 PTEN、PI3KCA、Caspase-9、MDM2 mRNA表达检测
主要步骤为总RNA抽提、逆转录、荧光定量PCR扩增等。表达量的统计方法为采用2﹣ΔΔCt法分析数据,得到各个标本基因的 Ct值,计算△Ct,△Ct=目的基因Ct-内参基因Ct;△Ct的相反数=﹣△△Ct,2﹣ΔΔCt=﹣-△△Ct 作 2 的幂运算,即 2﹣ΔΔCt=Fold change。序列参照Gene Bank 数据库中各目的基因及内参基因的序列,引物由NCBI Primer-blast设计,由苏州金唯智合成,引物序列见表1。
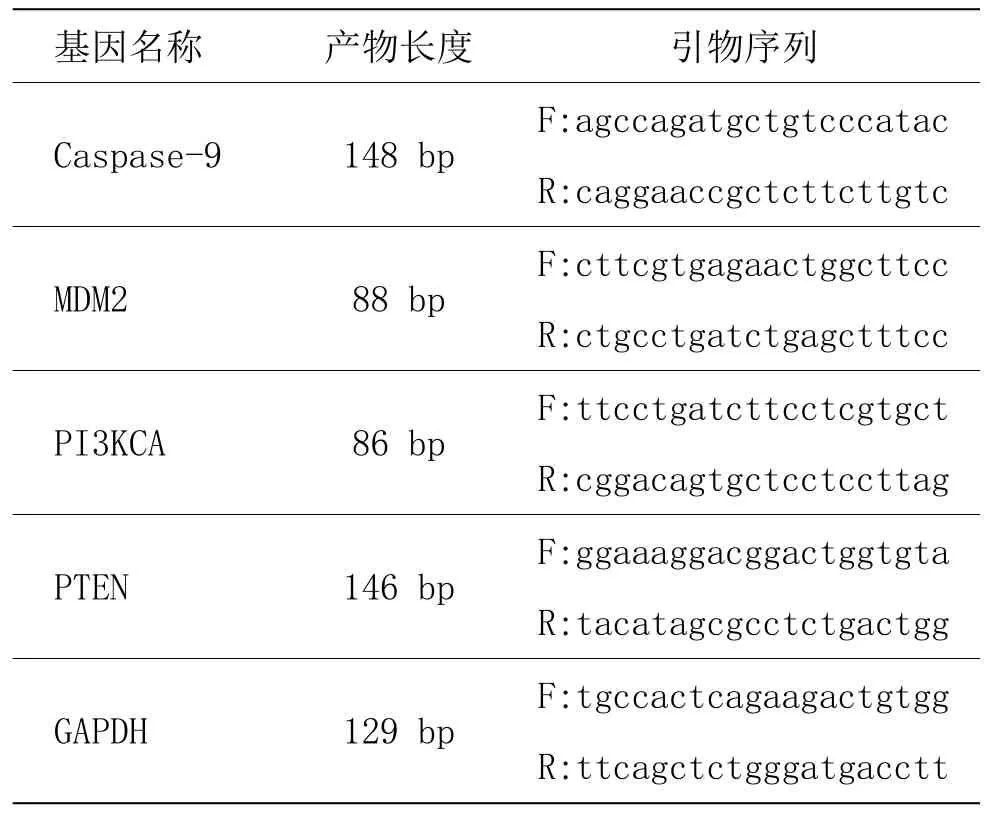
表1 各检测指标引物序列
1.7 统计学方法
采用SPSS21.0统计软件对数据进行统计分析。符合正态分布的计量资料以均数±标准差表示,不符合正态分布的数据用中位数(四分位数间距)表示,若数据方差齐则组间比较采用完全随机单因素方差分析(One-wayANOVA),进一步采用SNK-q检验、LSD-t检验行多重比较;不符合正态性的计量资料采用非参数检验(Mann-WhitneyTest)。检验水准α=0.05,以P<0.05表示差异具有统计学意义。
2 结果
2.1 各组大鼠胃窦组织病理学改变
正常组大鼠胃黏膜固有腺体结构排列整齐,黏膜层少见中性粒细胞、淋巴细胞,但胃主细胞、壁细胞未见减少;模型组大鼠固有腺体结构紊乱,腺体减少,变薄,形态不一,成纤维细胞增生,可见杯状细胞,发生萎缩、肠上皮化生、假幽门腺化生和(或)异型增生,甚至个别大鼠达到低分化腺癌,黏膜层及黏膜下层有大量的中性粒细胞、淋巴细胞、单核细胞等。经4周治疗后,与模型组比较,西药组、隔药饼灸组大鼠固有腺体排列不规整,黏膜层及下层炎性细胞相对减少,固有腺体萎缩、肠上皮化生、异型增生减轻。详见图1。

图1 各组大鼠胃窦组织病理形态比较(HE染色,×200)
2.2 各组大鼠胃窦组织中 PTEN、PlP2、p-mTOR、Caspase-9和MDM2蛋白表达比较
2.2.1 各组大鼠胃窦组织PTEN蛋白表达比较
正常组胃窦组织PTEN蛋白高表达,胃黏膜固有腺体内细胞核呈大面积棕褐色颗粒,胞浆黄染,分布密集;模型组胃窦组织PTEN蛋白阳性表达较低,胃黏膜固有腺体内少量表达,染色为浅蓝;与模型组比较,隔药饼灸组和西药组胃窦组织 PTEN蛋白在固有腺体内呈散在分布黄色颗粒,主要集中在固有腺体基底部,胞浆有黄染的趋势。与正常组比较,模型组胃窦组织PTEN蛋白表达显著降低,差异有统计学意义(P<0.05);与模型组比较,西药组和隔药饼灸组胃窦组织PTEN蛋白阳性表达显著升高,差异有统计学意义(P<0.05)。详见图2、表2。
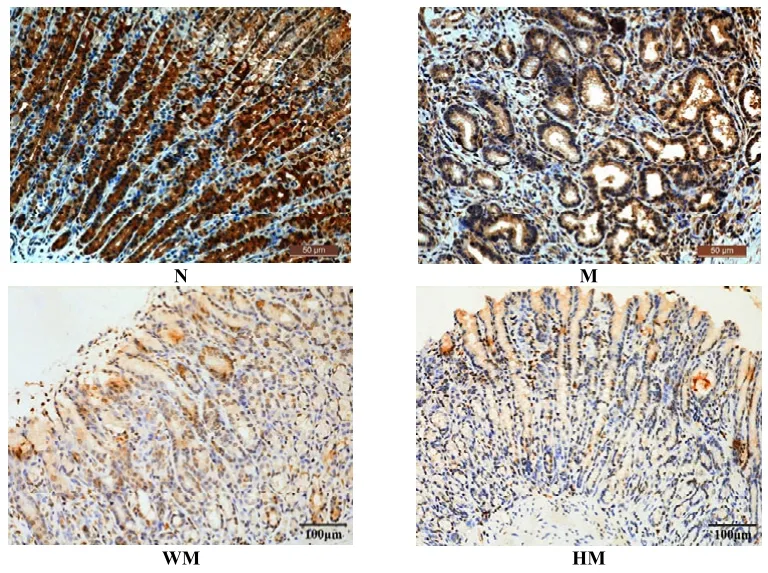
图2 各组大鼠胃窦组织PTEN的蛋白表达比较(lHC,×200)
表2 各组大鼠胃窦组织PTEN蛋白表达比较 (±s)
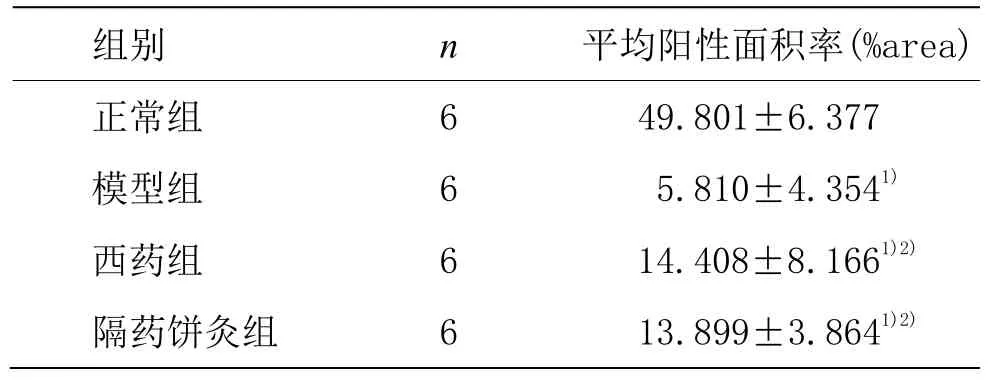
表2 各组大鼠胃窦组织PTEN蛋白表达比较 (±s)
注:与正常组比较 1)P<0.05;与模型组比较 2)P<0.05
组别 n 平均阳性面积率(%area)正常组 6 49.801±6.377模型组 6 5.810±4.3541)西药组 6 14.408±8.1661)2)隔药饼灸组 6 13.899±3.8641)2)
2.2.2 各组大鼠胃窦组织P-AKT蛋白表达比较
正常组胃窦组织 P-AKT蛋白阳性表达较少,仅见固有腺体基底部少量胞浆黄染,着色较浅;模型组胃窦组织 P-AKT蛋白阳性表达增多,胃黏膜固有腺体大面积胞浆黄染,着色较深;与模型组比较,隔药饼灸组和西药组胃窦组织 P-AKT蛋白阳性表达减少,胞浆着色变淡。与正常组比较,模型组胃窦组织P-AKT蛋白阳性表达显著上升,差异有统计学意义(P<0.05);与模型组比较,西药组和隔药饼灸组胃窦组织 P-AKT蛋白阳性表达下调,具统计学意义(P<0.05)。详见图3、表3。
表3 各组大鼠胃窦组织P-AKT蛋白表达比较 (±s)

表3 各组大鼠胃窦组织P-AKT蛋白表达比较 (±s)
注:与正常组比较 1)P<0.05;与模型组比较 2)P<0.05
?
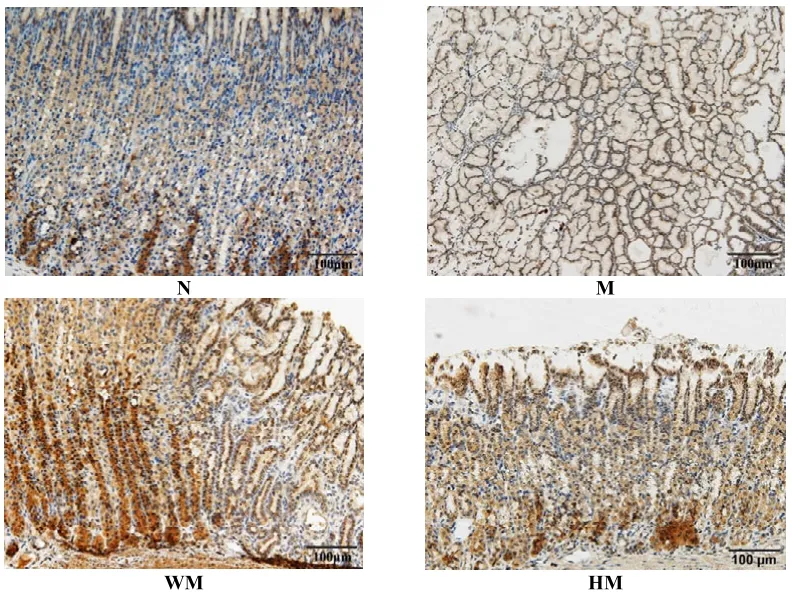
图3 各组大鼠胃窦组织P-AKT的蛋白表达比较(lHC,×200)
2.2.3 各组大鼠胃窦组织PIP2蛋白表达比较
正常组胃窦组织PIP2蛋白有少量表达;模型组大鼠胃窦组织 PIP2蛋白在腺体内高表达,染色深,棕褐色颗粒分布密集;与模型组比较,隔药饼灸组和西药组胃窦组织PIP2蛋白表达不同程度减少,散在分布黄色颗粒,染色变淡。与正常组比较,模型组胃窦组织PIP2蛋白表达显著增加,差异有统计学意义(P<0.05);与模型组比较,西药组和隔药饼灸组胃窦组织PIP2蛋白表达显著降低,均具统计学意义(P<0.05)。详见图4、表4。
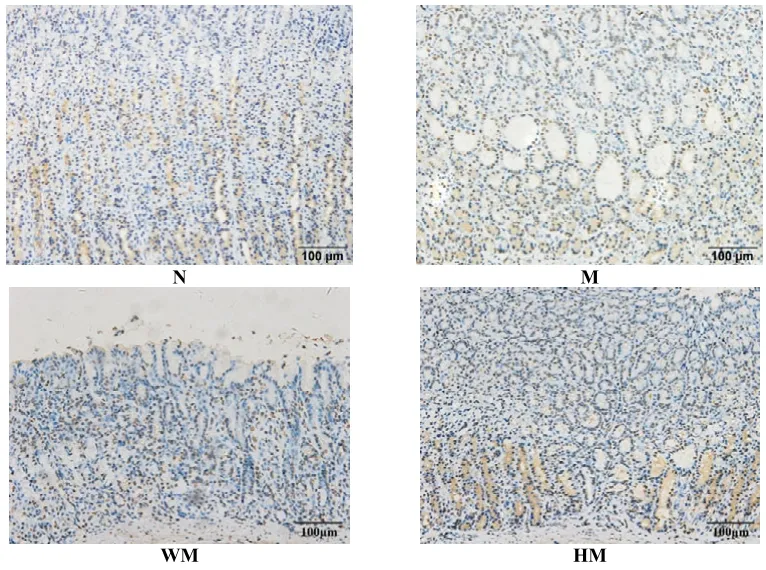
图4 各组大鼠胃窦组织PlP2的蛋白表达比较(lHC,×200)
表4 各组大鼠胃窦组织PlP2蛋白表达比较 (±s)
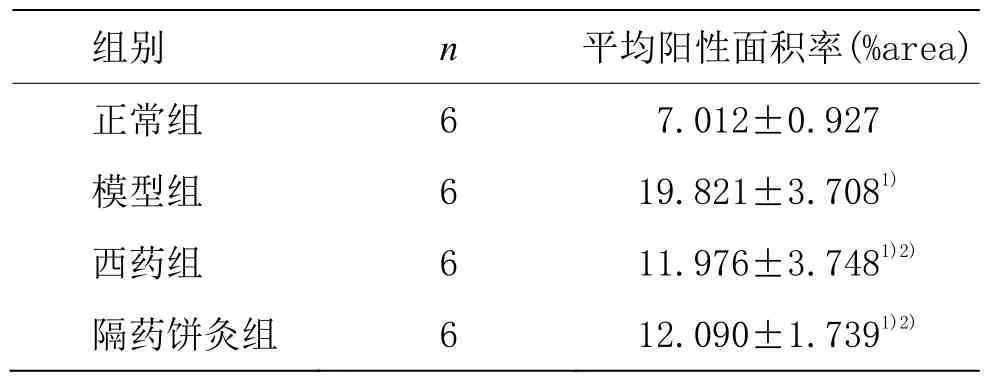
表4 各组大鼠胃窦组织PlP2蛋白表达比较 (±s)
注:与正常组比较 1)P<0.05;与模型组比较 2)P<0.05
?正常组 6 7.012±0.927模型组 6 19.821±3.7081)西药组 6 11.976±3.7481)2)隔药饼灸组 6 12.090±1.7391)2)
2.2.4 各组大鼠胃窦组织MDM2蛋白表达比较
正常组胃窦组织MDM2蛋白呈低表达,胞浆染色为浅蓝色;模型组大鼠胃窦组织MDM2在腺体内呈高表达,胞浆黄染,弥漫分布,胞核棕褐色颗粒分布密集;与模型组比较,隔药饼灸组和西药组胃窦组织MDM2蛋白阳性表达降低,胞核上的黄色颗粒变少,胞浆染色变浅。与正常组比较,模型组大鼠的 MDM2蛋白表达增加,差异有统计学意义(P<0.05);与模型组比较,西药组和隔药饼组胃窦组织MDM2蛋白表达显著降低,差异有统计学意义(P<0.05)。详见图5、表5。
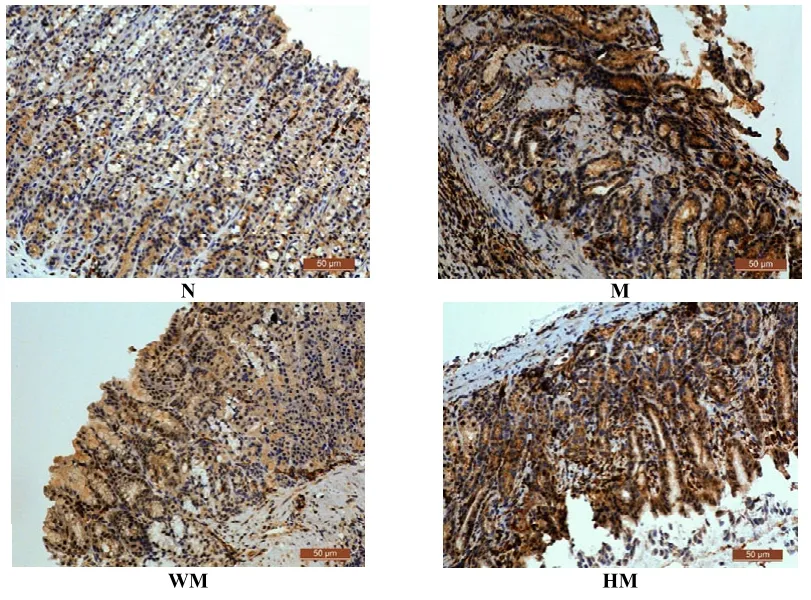
图5 各组大鼠胃窦组织MDM2的蛋白表达比较(lHC,×200)
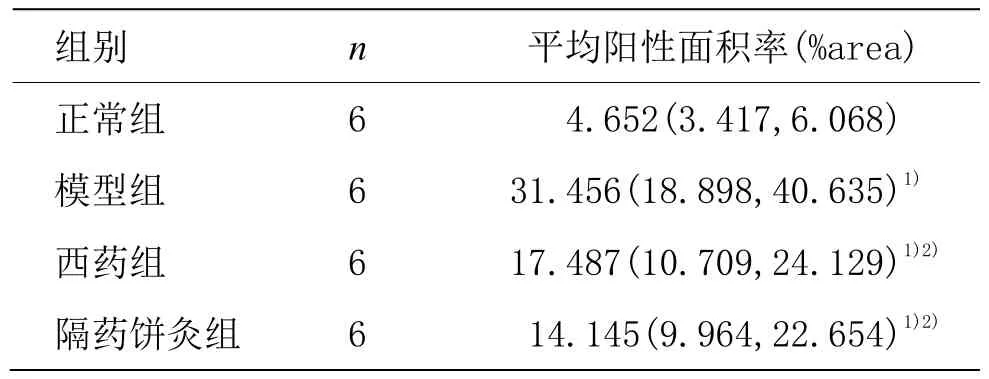
表5 各组大鼠胃窦组织MDM2蛋白表达比较 [M(P25,P75)]
2.2.5 各组大鼠胃窦组织Caspase-9蛋白表达比较
正常组胃窦组织 Caspase-9蛋白高表达,胃黏膜腺体内大面积棕褐色颗粒,胞浆弥漫性黄染;模型组胃窦组织 Caspase-9蛋白低表达,胃黏膜腺体内少量表达,染色为浅蓝;与模型组比较,隔药饼灸组和西药组胃窦组织 Caspase-9蛋白阳性表达有不同程度增加,主要表现在胃黏膜腺体基底部的黄色颗粒增加,胞浆呈淡黄色。与正常组比较,模型组胃窦组织Caspase-9蛋白阳性表达明显降低,差异有统计学意义(P<0.05);与模型组比较,西药组和隔药饼灸组胃窦组织Caspase-9蛋白表达量增加,均具统计学意义(P<0.05)。详见图6、表6。
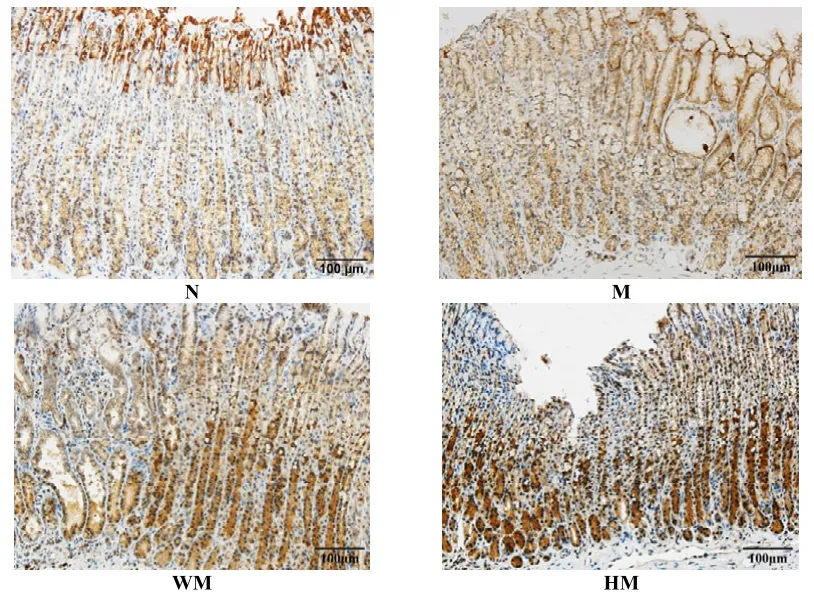
图6 各组大鼠胃窦组织Caspase-9的蛋白表达比较(lHC,×200)
表6 各组大鼠胃窦组织Caspase-9蛋白表达比较(±s)

表6 各组大鼠胃窦组织Caspase-9蛋白表达比较(±s)
注:与正常组比较 1)P<0.05;与模型组比较 2)P<0.05
组别 n 平均阳性面积率(%area)正常组 6 29.700±5.373模型组 6 8.009±2.8971)西药组 6 16.057±3.3541)2)隔药饼灸组 6 14.622±4.4021)2)
2.3 各组大鼠胃窦组织 PTEN、Pl3KCA、Caspase-9和P53 mRNA表达比较
与正常组比较,模型组胃窦组织 PTEN、Caspase-9 mRNA表达均降低,差异有统计学意义(P<0.05)。与模型组比较,隔药饼灸组胃窦组织Caspase-9、PI3KCA mRNA表达均升高,差异有统计学意义(P<0.05);西药组胃窦组织 PTEN、Caspase-9、PI3KCA mRNA表达均升高,差异均有统计学意义(P<0.05)。详见表 7。

表7 各组大鼠胃窦组织PTEN、Pl3KCA、Caspase-9和P53 mRNA的表达比较
4 讨论
慢性萎缩性胃炎(chronic atrophic gastritis,CAG)是消化系统的常见病、多发病,本病迁延不愈发展易发生癌变。CAG到胃癌的演变是多因素、多阶段、多步骤的综合结果,目前公认的胃癌发生模式为正常胃黏膜、慢性胃炎、萎缩性胃炎、肠上皮化生、不典型增生最终发展为胃癌(gastric cancer, GC)[16]。本研究采用 MNNG诱导联合夹尾刺激加饥饿失常的综合方法复制了 CAG大鼠模型,其出现组织病理学变化与临床CAG的组织形态比较接近,可见炎症、萎缩、肠化和异型增生,甚至出现腺癌的变化。提示MNNG诱导的CAG动物模型比较适合用于基础研究[14,17]。
针灸在治疗慢性萎缩性胃炎中发挥了免疫调节作用。据报道艾灸可通过调节细胞内外信号分子的表达,抑制Hp对胃黏膜的炎性损伤[18]。笔者团队前期和本次研究结果均提示,隔药饼灸CAG大鼠中脘、气海两穴干预后,能观察到艾灸能改善 CAG大鼠的胃窦组织形态学的变化,减轻胃黏膜炎症反应,降低腺体萎缩、肠化和异型增生的程度[19]。中脘和气海皆为任脉要穴,中脘为胃之募穴,气海为肓之原,是目前灸法治疗CAG及调节免疫功能常用取穴思路[20-21]。
艾灸可通过多途径、多环节、多靶点治疗CAG大鼠。早期发现PTEN的杂合性缺失发生率随CAG到GC病理进展增加[22];临床胃黏膜组织PTEN表达随着病灶的加重而降低[23],相比于癌旁正常胃黏膜,异形增生和胃癌患者的胃黏膜PTEN表达具显著差异。胃癌患者癌组织发现多种形式的 PTEN的失活[24],除了胃癌组织外,PTEN在多种瘤组织或细胞内表达异常[25-28]。本研究发现,隔药饼灸干预CAG大鼠中脘和气海穴后,胃黏膜组织的PTEN蛋白表达明显增加,提示艾灸能促进抑癌基因 PTEN的蛋白及 mRNA表达,可能是艾灸抑制 CAG大鼠胃黏膜组织炎-癌转变的作用基础。
胃肠道肿瘤发病机制与抑癌基因丢失和癌基因活化导致信号通路的异常激活密切相关[6]。单核苷酸多态性研究指出PTEN、PI3KCA、AKT的遗传变异与胃癌的复发和预后相关[29]。CAG胃黏膜病灶病理程度升高,P-AKT阳性表达亦高[30]。对 1 275例 GC患者和1 436例对照病例进行对照研究指出PI3K/AKT通路参与胃癌的发生[31]。其中 P-AKT阳性表达高的 AKT rs1130233多态性与Hp阴性感染者的CAG风险增加有关,与Hp感染者的CAG到胃癌级联风险有关。本研究结果发现,艾灸能上调PI3KCA的mRNA表达水平,抑制PIP2蛋白表达,抑制AKT磷酸化,提示艾灸发挥抑癌作用可能是通过PTEN-AKT途径发挥作用。PI3K通过磷酸化磷脂酰肌醇 4,5-双磷酸[PI(4,5)P2]累积磷脂酰肌醇-3,4,5-三磷酸[PI(3,4,5)P3]活化 AKT,PTEN 可水解 PI(3,4,5)P3 和磷脂酰肌醇 3,4-双磷酸[PI(3,4)P2]的D3位磷酸拮抗PI3K/AKT信号[32],抑制细胞过度的生长与增殖,降低肿瘤发生可能。除了PI(3,4,5)P3之外,PI(3,4)P2与AKT活性直接相关[33],表明PIP2的生成或降解作为PI3K和PTEN的第二信使参与此促癌通路。CAG患者胃黏膜病理程度升高,P-AKT阳性率亦高[31]。磷脂酰肌醇3激酶(PI3K)是PTEN/PI3K/AKT通路的重要协调因子,PI3Ks分3类,其中I类PI3Ks依据调控模式分IA类和IB类,PIK3CA作为同源类IA催化亚型于肿瘤中广泛表达,其基因突变或异常表达是导致 PI3K/AKT信号通路过度激活的关键因素[34-36]。
本研究结果还发现,隔药饼灸可能通过上调PTEN-AKT抑癌途径,抑制 MDM2蛋白活性,促进Caspase-9凋亡蛋白的表达,修复损伤的胃黏膜,改善CAG大鼠胃黏膜的萎缩、肠化生和异型增生的病理状态,防治CAG发生癌前病变。PI3K/AKT信号通路中的AKT活化可磷酸化MDM2或Caspase-9。在CAG到GC的病理进程,随着程度加重,胃黏膜病变组织 MDM2蛋白阳性表达也逐增[37],表明MDM2参与胃黏膜病变的发生发展。据报道 PTEN的过表达抑制 AKT活化,激活Caspase-9从而促进细胞凋亡[38];AKT活化诱导Caspase-9的前体(pro caspase-9)第196位Ser磷酸化,阻断其与凋亡活化因子结合,抑制Caspase-9促凋亡信号[39-41],抑制胃癌前病变细胞凋亡,促进CAG到GC的级联进程。
综上所述,艾灸有调节免疫、改善血液循环、抗癌及抗衰老等作用[42]。CAG伴有胃腺部分萎缩、黏膜受损和炎症,艾灸对 CAG临床治疗有改善胃部不适等症状,也能减轻胃黏膜炎症。代谢组学及外周血差异表达基因研究,说明艾灸治疗 CAG大鼠效应机制涉及信号转导、癌症、免疫疾病、氨基酸代谢、能量代谢、脂质代谢等通路[19,43]。本研究从CAG胃黏膜的PTEN表达降低,伴随AKT的过度激活的角度,研究具有脂质和蛋白磷酸酶活性的抑癌基因 PTEN,作用于 PIP2,通过PI3K/AKT的信号通路,介导了MDM2和Caspase-9的生物学功能。但也有研究发现MNNG造模的CAG大鼠胃黏膜PTEN mRNA和蛋白的表达降低,而PI3K、AKT mRNA和蛋白表达升高[44]。通过本研究,艾灸中脘、气海穴干预 CAG大鼠的胃黏膜保护效应可能是通过 PTEN-AKT抑癌途径,影响MDM2和Caspase-9的生物学功能而发挥作用的。
