单眼剥夺性弱视小鼠图形视觉诱发电位时间频率调制变化的研究
2022-01-04马博文吕文超王珏张伟史学锋
马博文,吕文超,王珏,张伟,史学锋
在视觉发育早期,由于异常视觉经验引起的视觉神经系统发育异常可导致弱视[1-2]。弱视在儿童中的患病率为2%~3%[3]。传统观点认为,弱视是以空间分辨力(视锐度)下降、对比敏感度降低和整体加工能力降低等为特点的空间信息处理能力缺陷为主的疾病[4]。然而,近年有研究表明,在弱视患者中,视觉相关的时间信息处理能力也会发生改变[5-6]。诸多与时间频率有关的指标,如临界融合频率、时间对比敏感度[7]、时空对比敏感度、时间超锐度等均会出现一定程度的损害。有关弱视猴的研究显示,与正常眼相比,弱视眼在高时间频率时的时间对比敏感度差异大于低时间频率时的差异[8]。因此,深入研究弱视的时间信息处理能力缺陷及其机制有重要的临床意义。视觉诱发电位(visual evoked potential,VEP)是从视皮层脑电活动中提取的视觉刺激诱发的电生理信号,是评估弱视视觉功能的一种重要的客观检查方法。本研究采用基于脑表面埋置电极记录[9]的方法观察单眼剥夺性弱视小鼠图形视觉诱发电位(pattern visual evoked potential,PVEP)的时间频率调制的变化,为深入探讨弱视的时间信息加工功能损害机制建立基础。
1 材料与方法
1.1材料 C57BL/6J雄性SPF级小鼠18只,生后26 d,体质量12~15 g,购自北京维通利华实验动物技术有限公司,屈光间质透明无病变。将小鼠按照随机数字表法分为对照组和单眼剥夺(monocular deprivation,MD)组,每组9只。动物饲养于中国医学科学院放射医学研究所实验动物中心,饲养环境通风良好,温度18~26℃,采用12 h/12 h光照/黑夜环境交替饲养,饲养条件符合医学实验动物饲养要求(伦理审查编号:IRM-DWLL-2020159)。
1.2研究方法
1.2.1单眼剥夺性弱视模型的建立 小鼠于麻醉条件下,剪去右眼的上下睑缘,采用10/0尼龙线将上下睑缘对齐缝合,在切口处涂典必殊眼膏预防感染。术后每天检查切口愈合情况,5 d后拆除缝合线,打开已闭合的眼睑,并在显微镜下观察屈光间质的透明度。如遇造模中途眼睑开裂,或造模完成后发现屈光间质混浊等异常情况,则不予入组并以新鼠替补,重新造模。见图1。
1.2.2电极埋置 采用脑表面电极埋置技术[9]。小鼠行5%水合氯醛(10µL/g)麻醉后,将其固定在立体定位仪上。使用体温维持仪保持其体温在36.5℃。双眼涂眼膏或硅油以防止其角膜干燥。碘附消毒后,剪去小鼠颅骨表面的皮毛,于记录眼对侧视皮层双眼区(后囟旁3 mm),以1.5 mm为直径开一个微小颅窗,保留硬脑膜,并将自制金属电极放置在视皮层表面硬脑膜上,将透明玻璃片盖于其上,并用生物胶固定,即完成记录电极的埋置。于记录眼同侧额骨上,用相同方法埋置参考电极。最后使用牙科水泥覆盖暴露的颅骨。待小鼠在体温维持仪上苏醒后,再将其放回笼中。

Fig.1 Monocularly deprived mouse model图1 单眼剥夺性弱视小鼠模型
1.2.3数据采集 于电极埋置术后第1天,将小鼠麻醉后,待其尾部夹捏反应消失,将其置于视觉刺激器屏幕正前方15 cm处。将记录电极和参考电极与前置放大器的电极接头相连。记录过程中若小鼠出现夹捏反应,则追加少量麻醉药。测试过程中使用的视觉刺激采用自编的Matlab程序控制。采用垂直光栅图形刺激,光栅空间频率一致,均为0.02周/度,叠加次数240次,屏幕亮度20坎德拉/平方米(cd/m2),对比度为80%。按照事先设计的伪随机序列给予小鼠6种不同时间频率的视觉刺激。6种时间频率分别为2.50、1.25、1.00、0.75、0.50和0.25 Hz。相邻2次刺激间在空间相位上以180°翻转,后一次刺激发生的时间间隔为伪随机序列确定的相应时间频率的倒数。所有数据采集过程均在暗环境中进行。记录到的视觉诱发电位信号通过CED1401数据采集系统(英国Cambridge Electronic Design公司)采集并通过Spike2软件(英国Cambridge Electronic Design公司)保存在电脑中。将采集到的数据导入Matlab软件,用自编的程序对数据进行叠加平均处理,分析不同刺激条件下得到的PVEP P100波振幅。
1.3统计学方法 采用GraphPad Prism 9.1.1统计软件(美国GraphPad Software公司)进行数据分析。数据均进行正态分布检验,采用±s形式表示。多组间比较采用单因素方差分析,组间多重比较采用Tukey法。2组间相同时间频率条件下PVEP测量结果的比较采用独立样本t检验。以P<0.05为差异有统计学意义。
2 结果
2.1 MD小鼠PVEP反应的时间频率调制特性的改变 对照组小鼠对所有6种时间频率的PVEP P100波反应幅值差异无统计学意义(F=2.214,P>0.05),见图2。MD组小鼠对6种时间频率的PVEP P100波反应幅值差异有统计学意义(F=6.588,P<0.01),其时间频率调制曲线表现为低通特性,在高时间频率刺激条件下的反应显著低于低时间频率刺激条件下反应,见图3。
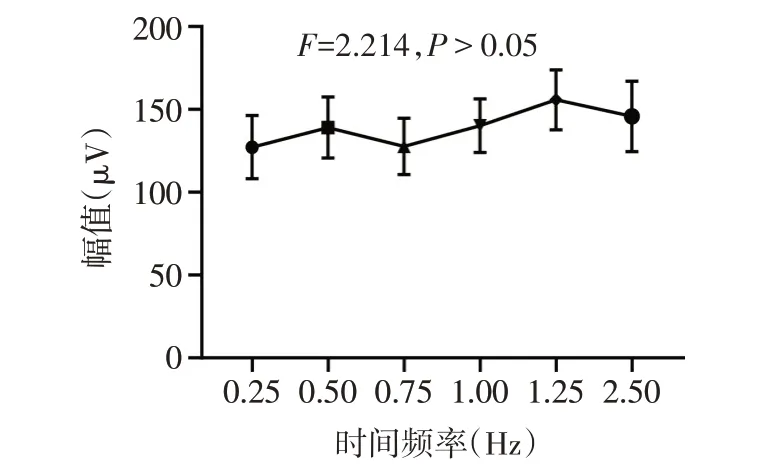
Fig.2 Temporal frequency tuning curve of PVEP responses in the Ctrl group图2 对照组小鼠PVEP反应的时间频率调制曲线
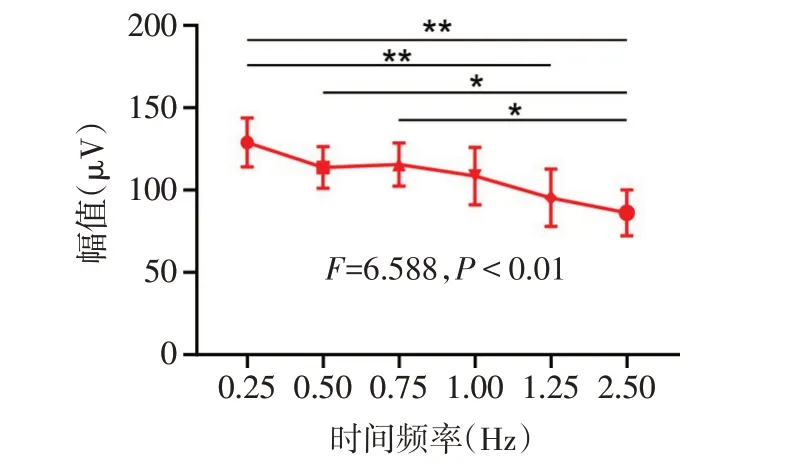
Fig.3 Temporal frequency tuning curve of PVEP responses in the MD group图3 MD组小鼠PVEP反应的时间频率调制曲线
2.2 不同时间频率刺激条件下2组小鼠PVEP反应幅值的比较 高时间频率(2.50和1.25 Hz)视觉刺激条件下,与对照组(145.7µV±63.8µV,155.7µV±54.2µV)相比,MD组(85.7µV±41.6µV,94.8µV±52.3µV)P100波幅值显著降低(t分别为2.362和2.425,P<0.05)。在中低时间频率(1.00 Hz、0.75 Hz、0.50 Hz、0.25 Hz)刺激条件下,2组间P100波反应幅值差异无统计学意义(t分别为1.347、0.583、1.155和0.054,均P>0.05)。
3 讨论
弱视是指在儿童视觉发育早期由于单眼斜视、未矫正的屈光参差、高度屈光不正及形觉剥夺等异常视觉经验而导致的单眼或双眼视力受损的一种神经发育异常性疾病[10-11]。弱视引起的视觉损伤可伴随终身且较明显,是一个重要的公共健康问题,其中单眼形觉剥夺性弱视较双眼弱视后果更为严重。在视觉发育关键期,视觉神经系统的发育非常迅速,这一时期各种异常视觉经验即使是短暂的作用都极易干扰和破坏神经系统的正常发育,导致视力、立体视功能、眼球运动功能等发育的异常[12]。在视觉神经系统中,视觉刺激引起视皮层神经元群体产生的信号必须进行整合以形成视觉感知[13]。正常的视觉信息的整合既包括空间信息的整合,也包括时间信息的整合。既往研究发现,大鼠外侧膝状体神经元的时间频率调谐特性随着发育逐渐成熟,其最优时间频率至成年时达到最高值[14]。有研究推测,弱视患者之所以对高时间频率视觉刺激的敏感性下降,可能原因是视觉系统对视觉信息的传递和整合存在缺陷[15]。功能性磁共振研究也发现,弱视患者对视觉信息的处理能力受损[16],并且可能存在相关视皮层区域的神经解剖学损害[17]。Yang等[6]通过心理物理学研究认为,弱视患者存在时间频率甄别缺陷。然而,目前尚鲜见有关弱视的时间信息整合能力缺陷的神经回路和细胞分子机制及干预策略的深入研究,迫切需要在整体动物水平建立可客观评估的模型。PVEP是一种客观评估整体视功能的技术手段。小鼠是广泛应用于眼科与视觉科学研究的模式动物,其饲养简单、成本较低,且有大量以C57品系为背景的转基因小鼠可供深入开展相关机制的研究。然而,由于小鼠的PVEP记录难度较高,以小鼠为模型动物开展的有关弱视时间信息整合能力缺陷的研究鲜见报道。本研究采用基于脑表面埋置电极记录[9]的方法探讨了MD小鼠PVEP的时间频率调制的变化,揭示了MD组小鼠的时间频率调制曲线表现为低通特性,其在高时间频率刺激条件下的反应显著低于中低时间频率刺激条件。与对照组相比,高时间频率视觉刺激条件下,MD组反应显著降低,而在中低时间频率刺激条件下,MD组视觉反应的损害未受到明显影响。笔者推测MD小鼠视皮层神经元对于高时间频率的视觉信息的加工整合和分析能力降低,造成视觉神经元的反应受到抑制。
另外,在本研究中,对照组小鼠对6种时间频率的PVEP P100波反应幅值无显著差异,其时间频率调制曲线基本表现为全通特性,但是在1.25 Hz和0.50 Hz处存在幅度较小的2个峰。既往研究发现,采用扫描VEP可记录到双峰现象[18-19]。这种双峰是视觉信息在2个平行通道,即瞬态通道和持续通道交互作用的结果,2个通道具有不同的空间和时间处理能力。已知在视觉系统中存在2条主要的携带视觉信息的传导通路,即大细胞通路和小细胞通路。大细胞通路主要携带高时间频率和低空间频率的视觉信息,小细胞通路主要携带高空间频率和低时间频率的视觉信息[20]。2种细胞通路将视觉信息从视网膜传输到视皮层。本研究发现,MD组时间频率调制曲线既未出现双峰,也未在低空间频率段出现单峰,且在高时间频率段PVEP反应较对照组明显降低,提示在MD组小鼠两条通路的时间信息加工能力均存在损害,但以大细胞通路的时间信息加工功能受损更为明显。
