抽穗扬花期低温胁迫对双季晚稻生理特性的影响
2021-12-31李辰彦李祖军田雪飞方加海石庆华曾勇军李辉婕吴自明
李辰彦 李祖军,3 田雪飞 方加海 石庆华 曾勇军 李辉婕,2 吴自明,*
(1 江西农业大学作物生理生态与遗传育种教育部重点实验室/江西省重点实验室,江西 南昌 330045;2 江西农业大学人文与公共管理学院,江西 南昌 330045;3 贵州省农业科学院水稻研究所,贵州 贵阳 550006)
双季晚稻在生育后期常常会遭受“寒露风”低温天气的侵袭,孕穗期低温容易导致枝梗及颖花分化不良、每穗粒数减少,且阻碍花粉粒发育、降低花粉萌发能力;开花期低温会阻碍花药正常开裂,导致授粉受精不良,灌浆不利,空秕率增加,直接影响产量[1-2]。当水稻受到低温胁迫时,在生理上会导致叶片叶绿素含量逐渐下降,光合功能随之衰退,叶片净光合速率(net photosynthetic rate,Pn)、气孔导度(stomatal conductance,Gs)、胞间CO2浓度(intercellular CO2concentration,Ci)、蒸腾速率(transpiration rate,Tr)明显降低[3-5]。另外,研究表明,低温胁迫时水稻产生的活性氧增多,抗氧化酶活性增强,可以减少活性氧积累对植株的伤害[6-7];而且水稻体内脂质过氧化反应和氧化反应产生的丙二醛(malondialdehyde,MDA)会引起蛋白质、核酸等生命大分子的交联聚合,使植株受到损伤[8]。同时,低温胁迫下水稻植株体内的可溶性蛋白含量也有所提高,其可作为渗透调节物质维持细胞内环境的稳定[9]。
此外,低温胁迫还会影响稻米品质的形成。有研究表明,水稻在花后低温处理3 d,籽粒淀粉的理化性质发生改变,直链淀粉含量增加,淀粉膨胀力、水溶性降低,糊化温度升高[10]。水稻灌浆前期平均气温低于21℃时,会导致多数水稻品种的出糙率、精米率和整精米率下降[11]。但也有研究发现,拔节期低温胁迫虽然会对营养品质产生负面影响,但拔节期适当的低温胁迫处理可以提高糙米率,改善外观品质[12]。从RVA谱特征值分析,低温胁迫会造成稻米的峰值黏度、热浆黏度与崩解值下降,消减值与峰值时间上升[13]。为了进一步阐明低温胁迫对抽穗扬花期双季晚稻产量与品质影响的生理机制,本研究通过对4个不同晚稻品种在抽穗扬花期进行低温胁迫,分析其对晚稻光合特性、抗氧化酶活性、穗部性状及稻米品质等的影响,以期为双季晚稻抽穗扬花期耐低温品种选育与栽培提供理论依据。
1 材料与方法
1.1 试验材料
试验在2018年双季晚稻品种筛选基础上,参照韩龙植等[14]的研究以自然结实率作为孕穗期耐冷性评价指标,筛选出2个耐冷品种岳优27(杂交籼稻)、昌粳225(常规粳稻),2个冷敏感品种泰丰优208(杂交籼稻)、黄华占(常规籼稻)。试验材料由江西农业大学作物生理生态与遗传育种教育部重点实验室提供。
1.2 试验设计
试验于2019年6月中下旬(20号左右)育秧,7月10日移栽,每桶3株,置于田间保持良好生长,在抽穗扬花前水肥统一管理,于抽穗扬花期挂标识牌定穗,取其中长势相近的植株于人工气候室进行低温处理,每个品种20桶(模拟“寒露风”,温度设置:白天:6:00―9:00设置17℃、9:00―12:00设置23℃、12:00―15:00设置25℃、15:00―18:00设置23℃;晚上:18:00―次日0:00设置21℃、次日0:00―6:00设置15℃,平均温度18℃),分别处理3、6、9、12 d,然后立即取样进行生理生化分析。每个品种挑选长势相近的5桶置于自然气候中作为对照,最高气温35℃,最低气温21℃,平均气温26℃)。将低温处理到3、6、9、12 d的水稻各品种取出5桶,立即测定光合参数并取样,置于室外常温条件下进行正常生长,成熟后取样考察穗部性状并进行稻米品质测定。
1.3 测定项目与方法
1.3.1 光合参数测定 低温分别处理3、6、9、12 d后,立即测定水稻主茎剑叶光合参数。选择晴朗无云的天气,在上午9:00―12:00选择发育一致的剑叶,采用CI-340便携式光合测定仪(美国CID公司)测定其Pn、Gs、Ci、Tr等指标,每株测定5片叶,重复3次。
1.3.2 花粉活力测定 参照张治安等[15]的方法,稍作改进。在处理0、3、5 d后,每天上午9:00―11:00随机取3穗,取成熟且尚未开花的上、中、下3个部位颖花各10个,混合均匀后随机挑选其中3个制作花粉样品,载玻片上加1~2滴2,3,5-氯化三苯基四氮唑(triphenyltetrazolium chloride,TTC)溶液,用摄子捣碎花药,去掉花药壁组织,盖上盖玻片,置于35℃通风条件下反应3~5 min,显色后观察,每个载玻片观察3~5个视野,记录染色显示深红色、浅红色、无色的花粉粒。花粉活力=(深红色花粉粒个数+浅红色花粉粒个数)/(深红色花粉粒个数+浅红色花粉粒个数+无色花粉粒个数)×100%。
1.3.3 抗氧化酶活性测定 分别取低温胁迫3、6、9、12 d的水稻剑叶测定抗氧化酶活性。超氧化物歧化酶(superoxide dismutase,SOD)活性测定参照李合生[16]的氮蓝四唑(nitrotetrazoliumbluechloride,NBT)光还原法;过氧化物酶(peroxidase,POD)活性测定参照邹琦[17]的愈创木酚法;过氧化氢酶(catalase,CAT)活性测定采用紫外吸收法[18]。
1.3.4 脂类化合物与可溶性蛋白含量测定 分别取低温胁迫3、6、9、12 d的水稻叶片,测定丙二醛(MDA)及可溶性蛋白(soluble protein,SP)含量。MDA含量测定参照郝建军[19]的硫代巴比妥酸法;SP含量测定参照李合生[16]的考马斯亮蓝法。
1.3.5 穗部性状 收获后每个处理随机选5株考察其每穗总粒数、结实率、千粒重。
1.3.6 稻米品质 水稻籽粒收获3个月后对稻米品质进行测定,其中出糙率、精米率、整精米率、垩白粒率、垩白度、直链淀粉含量的测定方法参照中华人民共和国国家标准《GB/T 17891-2017优质稻谷》[20];蛋白质含量采用凯氏定氮法[16],先测定米粉中的含氮量,然后乘以换算系数5.95。
1.3.7 稻米淀粉粘滞性 采用Super 3型快速黏度分析仪(rapid viscosity-analyzer,RVA,澳大利亚 Newport Scientific公司)测定RVA,并用TCW(thermal cycle for windows)配套软件进行数据分析[21]。
1.4 数据分析
采用SPSS 20.0软件进行数据LSD显著性分析,采用Microsoft Excel 2020制图。
2 结果与分析
2.1 抽穗扬花期低温胁迫对不同晚稻品种剑叶光合参数的影响
抽穗扬花期低温胁迫下水稻剑叶光合作用变化不同。由图1可知,在常温对照下随着时间的延长,各晚稻品种光合参数并无明显变化。由图2可知,各晚稻品种随着低温处理时间的延长Pn、Gs均呈下降趋势,总体表现为岳优27>昌粳225>泰丰优208>黄华占,其中耐冷品种(岳优27和昌粳225)的Pn在不同处理下降低幅度较冷敏感品种(泰丰优208和黄华占)更小;Ci则随着低温处理时间的延长呈先降低后稳定的趋势,各品种总体表现为岳优27>昌粳225>泰丰优208>黄华占;各晚稻品种的Tr随着低温处理时间的延长表现出持续降低的趋势,但总体上耐冷品种Tr在各时期显著高于冷敏感品种。
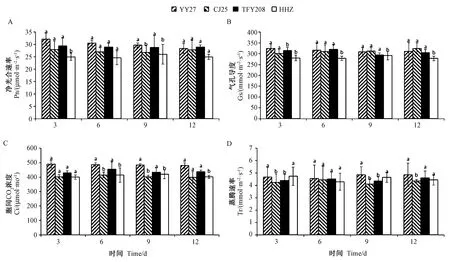
注:YY27:岳优27;CJ25:昌粳225;TFY208:泰丰优208;HHZ:黄华占。不同字母表示不同品种在同一处理时间差异显著(P<0.05)。下同。Note: YY27: Yueyou 27. CJ25: Changjing225. TFY208: Taifengyou208. HHZ: Huanghuazhan. Different letters indicate significant difference among different varieties of the same treatment time at 0.05 level. The same as following.图1 抽穗扬花期常温对照水稻剑叶光合参数Fig.1 Effects of normal temperature on photosynthetic parameters of rice flag leaf in rice at heading and flowering stage

图2 抽穗扬花期低温胁迫水稻剑叶光合参数Fig.2 Effects of low temperature stress on photosynthetic parameters of rice flag leaf in rice at heading and flowering stage
2.2 抽穗扬花期低温胁迫对不同晚稻品种花粉活力的影响
抽穗扬花期低温胁迫降低晚稻的花粉活力。由表1可知,随着低温处理时间延长各参试晚稻花粉深红色个数减少,浅红色和无色个数增加,且不同品种增加的个数不同,昌粳225增加个数最少。从各参试晚稻品种花粉活力来看,除处理3 d后昌粳225较处理0 d时显著降低,其他各品种间花粉活力差距不大;而处理5 d后,各品种均较处理0 d时显著降低,其中岳优27与昌粳225的花粉活力高于泰丰优208与黄华占。从各参试晚稻品种花粉活力降幅来看,低温胁迫5 d后,冷敏感品种泰丰优208和黄华占降幅较大,分别为15.57%和11.21%,而耐冷品种岳优27和昌粳225降幅分别为9.63%和4.38%。
2.3 抽穗扬花期低温胁迫对不同晚稻品种剑叶抗氧化酶活性及可溶性蛋白的影响
抽穗扬花期低温胁迫导致各晚稻剑叶抗氧化酶(除CAT外)活性及MDA、SP含量上升,而CAT活性呈先降低后升高的趋势。由图3-A可知,各晚稻品种随着低温处理时间的延长SOD活性变化规律为单峰曲线,呈现先升高后降低的趋势,具体表现为各晚稻品种SOD活性低温处理6 d后急剧上升达到最大值,而处理9和12 d后SOD含量逐渐下降。各晚稻品种在低温处理3、6、9 d后岳优27、昌粳225和黄华占的SOD活性显著高于泰丰优208。
由图3-B可知,POD活性随着低温处理时间的延长逐渐升高,并在低温处理12 d后达到最高水平,其中岳优27和昌粳225在除第9天外的各时期均显著高于泰丰优208和黄华占;且耐冷品种POD的增幅显著大于冷敏感品种。
由图3-C可知,CAT活性总体呈先降低后增加的趋势,且在低温处理3 d后各晚稻品种CAT活性最高。岳优27和昌粳225显著高于泰丰优208和黄华占。不同低温处理天数下CAT活性的降低幅度不同,冷敏感品种泰丰优208与黄华占降幅大于耐冷品种岳优27与昌粳225。
由图3-D可知,随着低温处理时间延长,各晚稻品种的MDA含量呈持续增加的趋势,均在低温处理9 d后达到最大值。总体上,泰丰优208和黄华占的MDA含量显著高于岳优27和昌粳225。各参试品种的MDA含量增加幅度不同,具体表现为黄华占>泰丰优208>昌粳225>岳优27。
由图3-E可知,各晚稻品种随着低温处理时间的延长,SP含量变化呈单峰曲线,泰丰优208和黄华占在低温处理6 d后达到最大值,而岳优27和昌粳225则于低温处理9 d后达到最大值。不同晚稻品种进行不同天数的低温胁迫,其SP含量变化不同,具体表现为黄华占在低温处理6 d后SP含量显著高于其他品种,岳优27和昌粳225在低温处理9 d后显著高于泰丰优208和黄华占,而低温处理12 d后昌粳225显著高于其他3个品种,表明在低温胁迫下耐冷品种的SP含量开始降低的时间较冷敏感品种延迟,随着时间的延长,耐冷品种的SP含量高于冷敏感品种。

图3 抽穗扬花期低温胁迫对水稻剑叶抗氧化酶活性与可溶性蛋白含量的影响Fig.3 Effects of low temperature stress on antioxidant enzyme activity and soluble protein content of flag leaves of rice at heading and flowering stage
2.4 抽穗扬花期低温胁迫对不同晚稻品种穗部性状的影响
耐冷晚稻品种结实率与千粒重受抽穗扬花期低温胁迫影响较小。由表2可知,抽穗扬花期低温处理对不同晚稻品种每穗粒数无明显影响。随着处理时间延长各晚稻品种的结实率和千粒重均有不同程度的降低,总体上,岳优27和昌粳225在不同时间低温胁迫下均高于泰丰优208和黄华占。从结实率上看,各晚稻品种结实率随着低温处理天数的延长而降低,并表现为冷敏感品种泰丰优208和黄华占降幅大于耐冷品种岳优27和昌粳225。从千粒重上看,不同品种千粒重降低程度不同,随着低温处理时间延长至12 d后,岳优27降低5.54%,昌粳225降低3.83%,泰丰优208降低6.73%,黄华占降低8.33%。

表2 抽穗扬花期低温对不同晚稻品种穗部性状的影响Table 2 Effect of low temperature on panicles of different late rice varieties at heading and flowering stage
2.5 抽穗扬花期低温胁迫对不同晚稻品种稻米品质的影响
从表3可知,抽穗扬花期低温胁迫降低了晚稻的加工品质和外观品质。各晚稻品种随着低温时间的延长,岳优27和昌粳225的出糙率、精米率和整精米率总体高于泰丰优208和黄华占。不同品种间的加工品质降幅有所不同,其中整精米率岳优27降幅低于其他品种;出糙率和精米率均以粳稻品种昌粳225降幅较大,而籼稻品种黄华占出糙率和精米率降幅较小。各品种垩白粒率与垩白度随着低温时间的延长呈逐渐增加趋势,其中粳稻品种昌粳225的垩白率与垩白度在各时期均高于其他品种。从增幅来看,岳优27和昌粳225的垩白粒率变化幅度小于泰丰优208和黄华占。垩白度变化趋势与垩白粒率相似,但岳优27和昌粳225垩白度的增幅大于泰丰优208和黄华占。

表3 抽穗扬花期低温对不同晚稻品种稻米品质的影响Table 3 Effect of low temperature on rice quality of different late rice varieties at heading and flowering stage
抽穗扬花期低温胁迫会使参试晚稻直链淀粉和粗蛋白含量增加,耐冷品种较冷敏感品种增加缓慢,呈现出更好的口感与营养。各晚稻品种随着低温处理时间的延长,胶稠度呈不同的变化趋势,其中岳优27呈逐渐增加的趋势,而泰丰优208呈先增加后降低的趋势,在处理9 d后到达最大值;昌粳225与黄华占则是随着低温处理时间的延长呈下降趋势。各晚稻品种随着低温处理时间的延长,直链淀粉含量总体呈先升高后降低的趋势,其中岳优27和昌粳225在处理6 d后达到最大值,而泰丰优208则在处理9 d后达到最大值,黄华占在处理12 d后达到最大值,且岳优27和昌粳225的直链淀粉含量在各时期总体低于泰丰优208和黄华占。各晚稻品种随着低温处理时间的延长,粗蛋白含量呈现不同的变化趋势。其中岳优27、泰丰优208与黄华占呈先升高后降低的趋势,不同的是岳优27在处理9 d后达到最大值,泰丰优208在处理6 d达到最大值;而黄华占在处理3 d后达到最大值;昌粳225则随着低温处理时间的延长,呈持续增加的趋势。总体上,岳优27的粗蛋白含量在各时期均高于其他品种,而黄华占则低于其他品种。
2.6 抽穗扬花期低温胁迫对稻米淀粉RVA谱特征值的影响
抽穗扬花期低温胁迫会影响稻米淀粉RVA谱的变化。由表4可知,各晚稻品种随着低温处理时间的延长,峰值黏度呈整体降低的趋势,其中岳优27和昌粳225降低幅度较大,在低温处理12 d后分别降低5.94%和12.28%。各晚稻品种的热浆黏度与对照相比总体上呈降低趋势,其中岳优27、昌粳225分别在低温处理9和12 d后达到最低值,而泰丰优208和黄华占在低温处理3 d后达到最低值。各晚稻品种的崩解值随着低温处理时间的延长呈降低趋势,其中泰丰优208在处理9 d后出现显著的降低;耐冷品种岳优27和昌粳225降幅较小,在低温处理12 d后分别较处理0 d时降低11.26%和23.37%。各低温处理中,岳优27和昌粳225的冷胶黏度和回复值在处理3 d后最高,随着低温处理时间的延长之后呈降低趋势;泰丰优208则随着低温处理时间的延长逐渐升高,在处理12 d后到达最大值;黄华占呈先增高后降低的趋势,冷胶黏度和回复值分别处理6和9 d后达到最大值。消减值中粳稻品种昌粳225为正值,其他品种均为负值,且泰丰优208和黄华占随着低温处理时间的延长呈增加趋势。低温胁迫对峰值时间和糊化温度影响较小,经低温处理后,耐冷品种岳优27和昌粳225的峰值时间和糊化温度与对照差异不显著;冷敏感品种泰丰优208和黄华占的峰值时间与对照差异不显著,但泰丰优208在低温处理6 d后和黄华占在低温处理9 d后的糊化温度较对照显著降低。
3 讨论
水稻在栽培过程中受温度影响较大,低温常导致水稻减产[22-23]。抽穗扬花期正值水稻生殖生长阶段,对低温最为敏感,该时期受低温影响会导致水稻花药开裂系数、花粉萌发率降低,不育花粉增多[24]。本研究发现,不同持续时间低温胁迫下各参试晚稻品种的花粉活力均降低,其中耐冷品种的花粉活力较冷敏感品种高,且降幅较低,表明耐冷品种在低温胁迫下具有较好的花粉活性,这与邓化冰等[25]的研究结果相一致。
光合作用可为植物提供生长发育所需物质,孕穗期低温胁迫使小麦光合速率降低,且低温处理时间越长,对光合速率影响越大[26]。水稻在孕穗开花期遭受低温胁迫亦会使Gs降低,影响光合速率[27]。本研究发现,随着低温处理时间的延长,各参试晚稻品种的Pn、Gs均降低。其中岳优27和昌粳225的Pn均高于泰丰优208和黄华占,降低幅度较小。Ci和Tr变化趋势与之相似,随着低温处理天数的延长呈持续增加的趋势,岳优27和昌粳225的变化幅度较小。说明随着低温处理时间的延长,岳优27和昌粳225的光合作用受低温胁迫影响较小。
植物体可通过抗氧化酶系统维持体内的自由基平衡,保护植物体免受自由基的伤害[28]。有研究表明,当水稻植株遭受低温冷害后,SOD活性快速上升,POD活性有一个缓慢上升的过程,CAT活性则随着低温胁迫程度的加剧而逐渐增强,SOD的变化先于POD和CAT[29]。本研究结果发现,SOD活性变化规律为单峰曲线,随着胁迫时间延长呈先升高后降低的趋势,POD活性逐渐升高,CAT活性呈先降低后升高的趋势。但总体来看,随着低温处理时间的延长耐冷品种岳优27和昌粳225的SOD、POD、CAT活性较大,SOD、POD在胁迫处理时间较短时增长幅度较高,而CAT活性稳定在一定水平上,变动幅度较小。MDA作为膜脂过氧化产物,其含量可表示植物遭受低温的伤害程度,而SP作为渗透调节物质,对低温胁迫下的细胞内环境相对稳定起到保护作用[8,30]。本研究发现,随着低温处理时间的延长,冷敏感品种在遭受低温胁迫时MDA含量较高,且随着处理时间的延长增加幅度较大;耐冷品种SP含量较先达到最大值,且随着低温处理时间的延迟,耐冷品种SP含量总体高于冷敏感品种。本研究发现,抽穗扬花期低温处理对不同晚稻品种的每穗粒数无明显影响,但随着低温处理时间的延长,各参试晚稻品种结实率和千粒重呈现出不同程度的降低。总体来看耐冷品种岳优27和昌粳225的结实率和千粒重降幅较小,这符合夏楠等[31]对寒地粳稻灌浆期低温胁迫后结实率与千粒重降低的研究。有研究表明,孕穗期低温冷害降低了水稻稻米的加工品质,致使糙米率、精米率和整精米率均有不同程度的下降,其中整精米率的降幅最大[32]。另外,有研究发现适当的低温有助于常规籼稻与常规粳稻蛋白质的积累,水稻受到低温胁迫后又会阻碍杂交籼稻蛋白质的积累,而蛋白质含量又与稻米的垩白粒率呈正相关[33-34]。本研究发现,低温胁迫下各参试品种的加工品质逐渐降低,其中整精米率下降幅度最大;随着低温处理时间的延长,各晚稻品种垩白粒率、垩白度、胶稠度增加,而直链淀粉与粗蛋白含量呈先增加有降低的趋势,相较于冷敏感品种,耐冷品种的精米率、垩白粒率、胶稠度变化幅度较小,而垩白度变化幅度较大。各晚稻品种直链淀粉含量均随低温处理时间的延长先增加后降低,其中岳优27和泰丰优208变化幅度较大,另外,粗蛋白含量各品种均呈先升高后降低的趋势,而常规品种昌粳225与黄华占变化幅度较小。在抽穗扬花期低温胁迫后水稻稻米品质与前人研究结论一致[31-34],随着低温胁迫时间的延长,加工品质、外观品质与食味品质各水稻品种都有明显的下降,但耐冷品种降幅较低;而营养品质方面,则是常规品种昌粳225与黄华占的粗蛋白含量受到低温胁迫的影响较小,说明水稻在抽穗扬花期受到低温胁迫时会影响花粉的活性导致灌浆时的效率降低与水稻籽粒的成熟度不足,从而影响了水稻的稻米品质。
RVA与稻米的蒸煮食味品质密切相关,一般情况下,口感较好的稻米RVA谱往往具有较高的峰值黏度、崩解值以及较低的消减值[35]。有研究认为,在灌浆结实期低温弱光复合胁迫下,峰值黏度、热浆黏度、崩解值均显著降低[36]。而褚春燕等[37]研究认为,孕穗至灌浆期进行低温处理,其稻米峰值黏度、冷胶黏度、热浆黏度降低,但对崩解值、消减值和回复值的影响较小。本研究发现,与常温对照相比,低温胁迫下各品种峰值黏度、热浆黏度和崩解值呈降低趋势,而冷胶黏度、回复值和消减值呈升高趋势。与冷敏感品种泰丰优208和黄华占相比,耐冷品种岳优27和昌粳225的峰值黏度、冷胶黏度和回复值变化幅度较大,热浆黏度、崩解值和消减值变化幅度较小。耐冷品种可能是由于有更好的花粉活性及灌浆效率,使其籽粒更加饱满优质,从而有着更出色的食味蒸煮品质。
4 结论
抽穗扬花期低温胁迫下,与冷敏感品种泰丰优208和黄华占相比,耐冷品种岳优27和昌粳225的花粉活力较高,光合作用受影响较小,抗氧化酶活性和SP含量较高,MDA含量较低;另外,耐冷品种结实率和千粒重降低幅度较低,稻米加工品质与外观品质降低幅度较低,蒸煮食味品质与营养品质优于冷敏感品种。总的来说,抽穗扬花期低温胁迫影响水稻的生长发育,而耐冷品种岳优27和昌粳225抗氧化物酶活、光合效率以及渗透性物质在体内升高,保证了其灌浆效率,从而导致耐冷品种产量与品质由于冷敏感品种。
