基于ITS和nLSU序列的木耳属物种鉴定
2021-12-30牛宇徐全飞聂建军潘保华冯婉君蒙秋霞赵悠悠
牛宇 徐全飞 聂建军 潘保华 冯婉君 蒙秋霞 赵悠悠
(1 山西农业大学农业经济管理学院,山西 太原 030006;2 山西农业大学资源环境学院,山西 太原 030031;3 山西省侯马市出入境检验检疫局,山西 临汾 043000)
木耳属Auricularia的物种是食用历史悠久的食药用菌,早在1 400多年前我国人民就开始人工栽培食用木耳[1]。据《神农百草经》记载,木耳“益气不饥,轻身强志”。木耳属主要栽培品种为黑木耳和毛木耳。根据中国食用菌协会统计,2018年我国黑木耳产量达674万t,占全国食用菌总产量的17.54%,主要产区为东北三省、河北省、山西省、陕西省、云贵川三省[2]。我国木耳属种质资源丰富,《中国食用菌名录》记载有13种,但我国具体有多少种木耳仍存在争议。山西省为木耳主要产区,因此对其木耳属栽培菌株和野生菌株进行物种鉴定,有助于研究木耳种质资源的多样性,也为木耳属种质资源的收集、评价、利用提供分子数据支撑。此外,野外环境中如黑耳属等部分银耳科物种的担子果外观形态与木耳属物种十分相似,但食毒不明,所以有必要借助形态学观察和分子系统学方法加以区别。
近年来,随着分子标记技术的发展和应用,木耳属物种的遗传多样性研究取得了长足的进步。吴芳等[3-4]结合ITS+nLSU+rpb2序列鉴定和形态学观察,研究了黑木耳复合群和毡毛木耳复合群的系统发育和多样性。张丹等[5]对5个人工栽培毛木耳菌株和2个野生分离的毛木耳菌株进行了ITS序列分析。陶鹏飞等[6]利用ITS和SRAP标记对黑龙江省境内采集的14株野生黑木耳菌株和1个南方栽培菌株进行遗传多样性分析。贾定洪等[7-8]对19个毛木耳栽培菌株和3个野生分离菌株进行了ISSR分析和基于ITS序列的系统发育研究。刘华晶等[9]借助ISSR和ITS分子标记对黑龙江省内部分野生黑木耳菌株进行了遗传多样性分析。刘虹等[10]对来自山西省木耳属9份人工栽培菌株和7份野生菌株进行基于ITS片段鉴定,鉴定出3个木耳属物种,即黑木耳A.heimuer、短毛木耳A.villosula和毛木耳A.cornea,但没有匹配到文献记录山西省有分布的木耳A.auricula、多毛木耳A.polytricha和皱木耳A.delicata。由于木耳属种类较多,且其子实体的形态、颜色、大小及子实层的形态常随生长环境及着生部位的不同而变化,因此同物异名或同名异物的情况时有发生[11]。当前人们应用分子生物学和遗传学新技术,通过对DNA等遗传物质的比较,可从分子层面鉴定木耳属物种并探索其进化关系。笔者基于ITS和nSLU序列,结合形态学观察,对7份山西省木耳属栽培菌株和4份野生菌株进行鉴定,以期更好地对山西省木耳属和部分形态近似的野生银耳科物种进行鉴别和分类,为进一步开发利用该物种资源提供科学依据。
1 材料与方法
1.1 供试菌株与培养基
供试菌株共11个,其中7个菌株为山西省常见木耳栽培品种,4个为野生菌株,于2018—2019年采集(表1)。供试菌株均用组织分离法获得纯培养物,担子果标本保存于山西农业大学农业资源与经济研究所食用菌实验室。
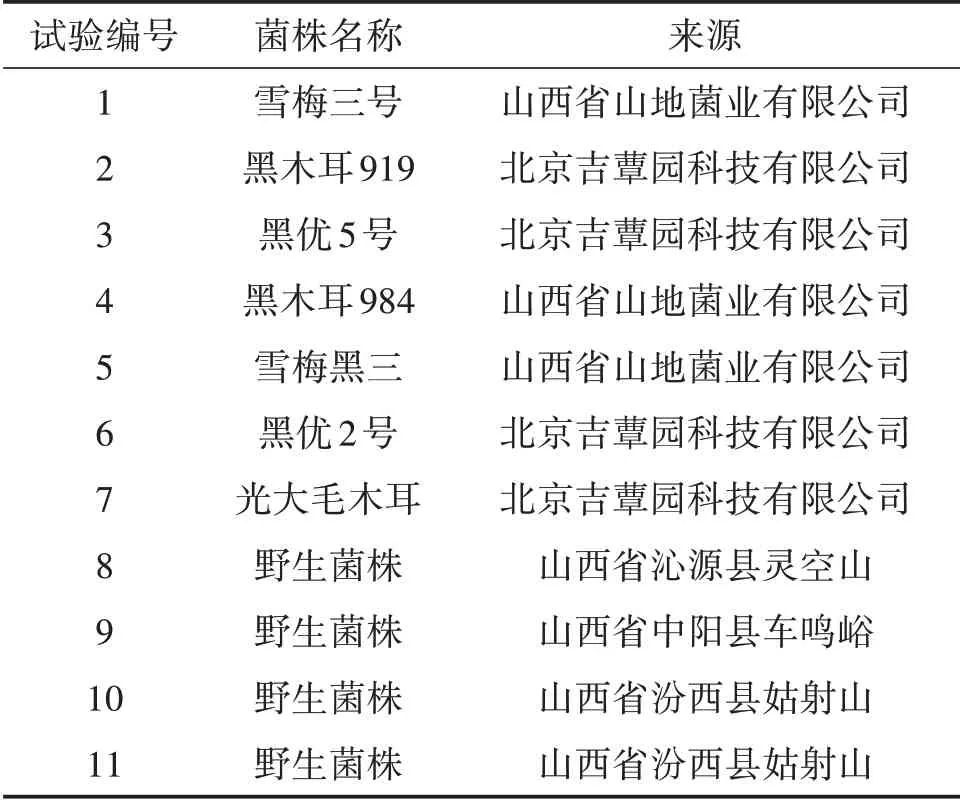
表1 供试菌株及来源
PDA固体培养基:马铃薯20%,葡萄糖2%,蛋白胨0.3%,硫酸镁0.2%,磷酸氢二钾0.2%,琼脂2%,pH自然。
1.2 试剂与仪器
DP302基因组DNA提取试剂盒,Taq DNA聚合酶,10×Buffer和6×Loading Buffer(北京天根生化科技有限公司);PCR引物(上海派森诺生物技术有限公司);脱氧核糖核苷三磷酸(dNTP)[宝日医生物技术(北京)有限公司];AxyPrep DNA凝胶回收试剂盒(杭州Axygen生物技术有限公司);Marker Plus(DL2000 Plus)(北京康润诚业生物科技有限公司);核酸染料和RNaseA(北京索莱宝科技有限公司)。
Neofuge 13R台式高速冷冻离心机(上海力申科学仪器有限公司);Applied Biosystems ABI-2720PCR仪,ABI 3730XL测序仪(上海爱普拜斯应用生物系统贸易有限公司);Major Science Mini Pro 300 V Power Supply电泳仪(上海珩璟科技有限公司);Labnet Sub System 70电泳槽(北京百奥创新科技有限公司)。
1.3 菌丝体的培养与收集
将各菌株纯培养物接入PDA培养基,培养6 d后用灭菌的手术刀片在不同菌株的菌落表面刮取少许菌丝,置于2.0 mL的离心管中,编号备用。
1.4 DNA的提取
提取样品DNA,并电泳,得到DNA电泳条带。
1.5 PCR扩增及测序
选用5条特异引物对11个菌株的DNA进行特异性扩增(表2)。50 μL反应体系成分:20 ng/μL基因组DNA、10×Buffer(含2.5 mmolL Mg2+)、5 U/μL Taq DNA聚 合 酶、10 mmolL dNTP、10 μM引 物、ddH2O。ITS序列PCR扩增反应条件:95℃预变性5 min,95℃变 性30 s,58℃退 火45 s,72℃延 伸1 min30 s,共34个循环,最后72℃延伸10 min,4℃保温。nLSU序列PCR扩增反应条件:95℃预变性1 min,95℃变 性30 s,50℃退 火1 min,72℃延 伸90 s,共35个循环,最后72℃延伸10 min,4℃保温。反应完成后,取3 μL PCR产物1%琼脂糖凝胶电泳检测,确认PCR扩增片段。回收、纯化后PCR产物送上海派森诺生物技术有限公司,在ABI 3730XL测序仪上进行DNA测序。

表2 试验所用的基因片段及其对应引物和引物序列

图1 供试菌株子实体
1.6 ITS和nLSU序列分析
用NCBI Blast程序将拼接后的序列文件与NCBI核酸数据库(https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastn&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome)中的数据比对,得到与待测物种序列相似性最大的物种信息。
1.7 构建系统发育树
将供试标本及来自GenBank登录木耳属、黑耳属、银耳属的54个菌株ITS序列、ITS+nLSU序列用Clustal X软件进行多序列比对分析,并用MEGA5.0软件中的邻接分析(Neighbor-Joining,NJ)法构建系统发育树,Bootstrap检验1 000次。
2 结果与分析
采用特异性ITS引物对供试菌株DNA进行PCR扩增,获得约650 bp的单一条带(图2、图3);采用特异性nLSU引物对供试菌株DNA进行PCR扩增,获得约1 000 bp的单一条带(图4),条带清晰明亮,无杂带、拖尾现象,扩增质量较好。
图2 供试菌株的ITS1-PCR凝胶电泳图
图3 供试菌株的ITS 5/4-PCR凝胶电泳图
图4 供试菌株的nLSU-PCR凝胶电泳图
基于ITS+nLSU序列构建了木耳属系统发育树(分支上的数字为用Bootstrap分析获得的支持值)。如图5所示,11份菌株聚在5个不同的末端分支。11份菌株的形态特征见表3。
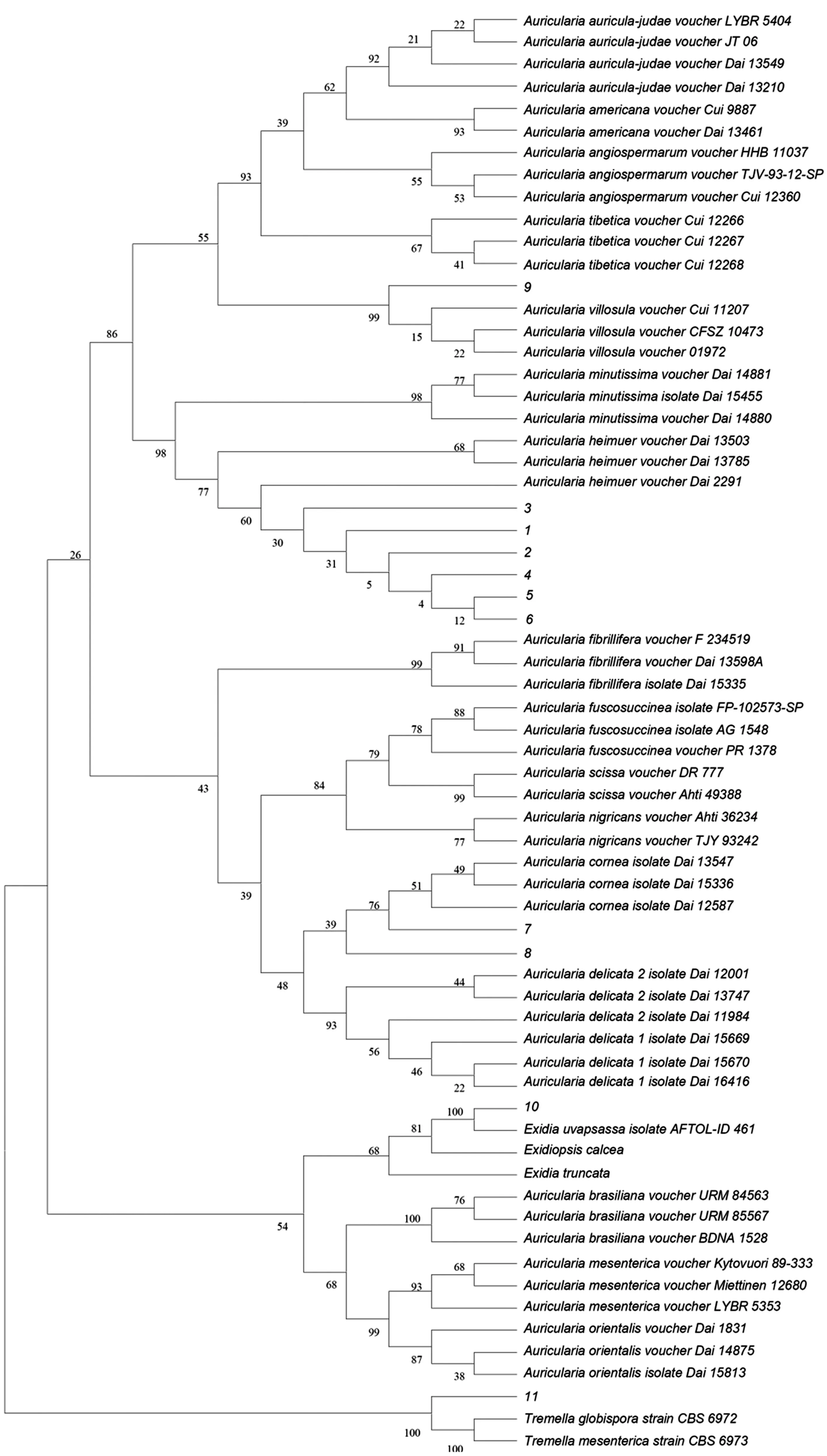
图5 基于ITS和nLSU序列的菌株和相关种属的木耳属系统发育树
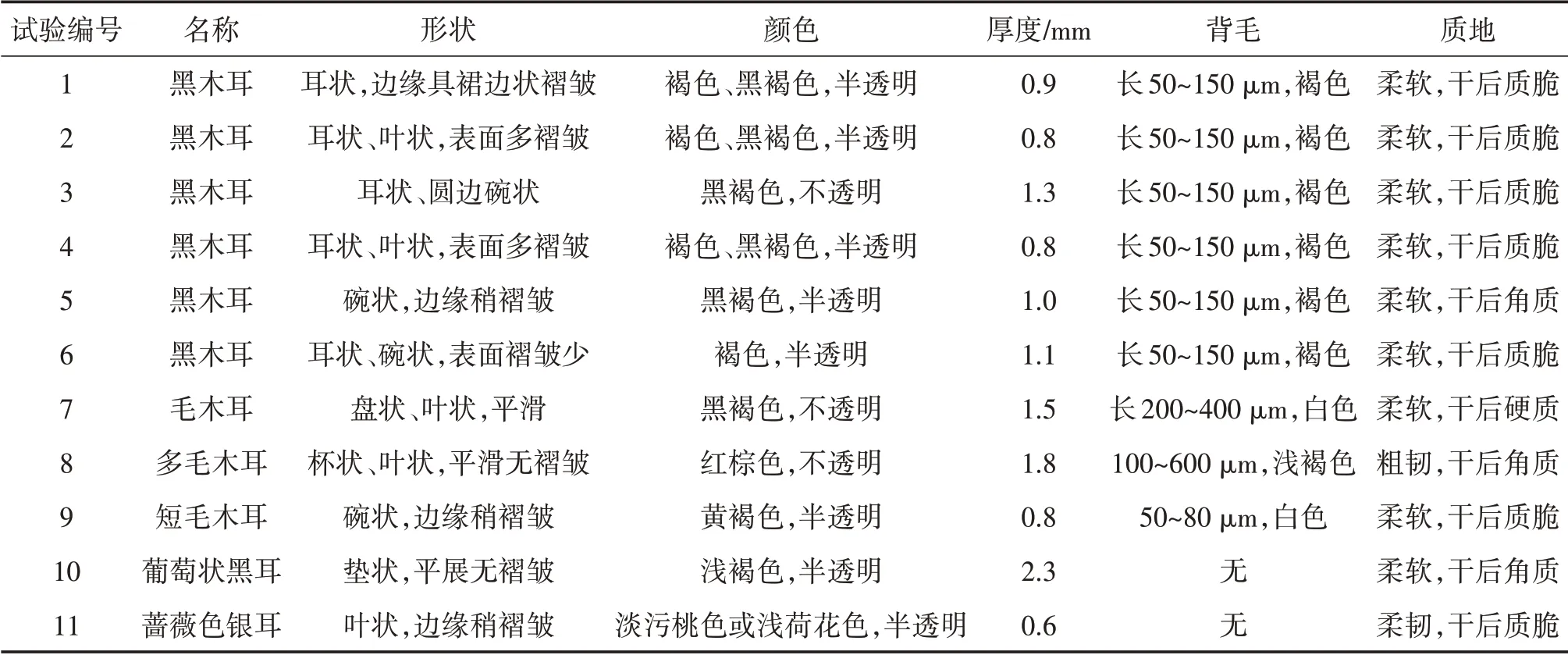
表3 供试11个菌株子实体形态特征
样本1、2、3、4、5、6与黑木耳A.heimuer的可信样本序列聚为一支,自举值为98%,结合担子果耳状、子实体层被毛少等特征,以及BLAST结果(ITS1、ITS5/4、nLSU序列相似性均在99.28%~100.00%),可以确定这些样本为黑木耳A.heimuer。
样本7聚在毛木耳A.cornea分支中,自举值为76%,结合其担子果褐色或红褐色、厚而坚韧、不孕面被明显长柔毛的形态特征,以及BLAST结果(ITS1、ITS5/4、nLSU序列相似性分别为99.82%、99.49%和99.93%),可以确定该样本为毛木耳A.cornea。
样本8也聚在毛木耳A.cornea分支中,自举值为39%,结合其担子果红褐色并带紫色、杯状或耳状、新鲜时胶质不透明、质地粗韧、平滑无皱褶、不孕面具致密长刚毛的形态特征,以及BLAST结果(nLSU序列相似性为99.63%),推测该样本为多毛木耳A.polytricha。
样本9聚在短毛木耳A.villosula分支中,自举值为99%,结合其担子果黄褐色、新鲜时胶质、杯状或盘状、不孕面具明显短柔毛的形态特征,以及BLAST结果(ITS1、ITS5/4、nLSU序列相似性分别为99.82%、98.81%和99.78%),可以确定该样本为短毛木耳A.villosula。
样本10聚在黑耳属分支中,自举值为100%,结合其担子果垫状、平展贴生、胶质、浅褐色的形态特征,以及BLAST结果(ITS1、ITS5/4、nLSU序列相似性分别为97.96%、98.60%和99.78%),可以确定该样本为银耳科葡萄状黑耳Exidia uvapassa。
样本11聚在银耳属分支中,自举值为100%,结合其担子果叶状、薄、丛生、胶质、淡污桃色或浅荷花色、干后锈褐色的形态特征,以及BLAST结果(ITS1、ITS5/4序列相似性分别为99.82%和99.16%),可以确定该样本为银耳科蔷薇色银耳Tremella roseotincta。
3 小结与讨论
基于ITS和nLSU序列分析,供试11份菌株中鉴定出4个木耳属物种,即黑木耳、毛木耳、多毛木耳和短毛木耳。其中7个山西省常见人工栽培木耳菌株中,除菌株7为毛木耳A.cornea外,其余菌株均聚在亚洲本地种黑木耳A.heimuer[4],可见山西省常见的木耳栽培种多数为黑木耳,这与前人[10]的研究结果相似。黑木耳是亚洲本地种,并被广泛栽培[12]。毛木耳也是亚洲物种,并在中国广为栽培[13],但在供试木耳栽培种中仅占很小的比例。
自2013年以来,多毛木耳A.polytricha被认为是黑色毛木耳A.nigricans的同物异名[14],检索真菌索引(Index Fungorum)和MycoBank数据库得出,该种的现用名为黑色毛木耳A.nigricans。供试的多毛木耳A.polytricha样本采自山西省沁源县,为文献记载的分布区域[15]。依据nLSU序列相似性结果(99.63%)推测供试野生菌株8为多毛木耳A.polytricha,然而其在进化系统树中并未与来自美国和哥斯达黎加的同物异名种A.nigricans的样本系列聚为一支,而是被聚在毛木耳一支,但自举值并不高(39%),且该物种与毛木耳可信样本的ITS序列相似度仅为97.03%。根据Renske划定的标准[16],序列相似度≥99%时,可以判定为同一个种,序列相似度在95%~99%时,可以判定为同一个属。可见,菌株8可能是一个与毛木耳A.cornea或黑色毛木耳A.nigricans不同的物种,其分类学地位有待于进一步研究。
短毛木耳A.villosula分布于亚洲温带和亚热带,在中国广泛分布,并被人工栽培[17],俄罗斯也有分布[18],采自山西省中阳县的野生菌株(菌株9)与其相似,但在人工栽培菌株中未发现。刘虹等[10]在山西省境内采集的2份短毛木耳菌株也均为野生。因此,山西省可能尚未人工栽培该物种。
2个银耳科物种葡萄状黑耳E.uvapassa、蔷薇色银耳T.roseotincta为山西省首次记录种。其中,葡萄状黑耳Exidia uvapassa为银耳科黑耳属真菌,已被发现分布于韩国东南部[19]、中部[20]及日本[21];蔷薇色银耳Tremella roseotincta也属于银耳科银耳属,分布于中国、日本[22]、俄罗斯[23],其形态与毡盖银耳Pha⁃eotremella fuscosuccinea(Tremella fuscosuccinea)非常相似,区别在于蔷薇色银耳生长于落叶树上,目前其真菌宿主尚不清楚,生态偏好也需进一步研究[24]。
