利用TRV过表达和沉默系统验证植物油脂合成相关基因的功能
2021-12-27谢桂兰
江 悦, 谢桂兰, 陈 放, 徐 莺
(四川大学生命科学学院生物资源与生态环境教育部重点实验室, 成都 610064)
1 引 言
烟草脆裂病毒(Tobacco Rattle Virus, TRV)是一种双正链RNA病毒,其基因组由RNA1和RNA2两部分组成.RNA1的大小和基因序列比较保守,主要包括依赖于RNA的RNA聚合酶(RNA dependent RNA polymerase, RdRp)和移动蛋白(Movement Protein, MP),其可以使病毒粒子在宿主植物内复制和移动. RNA2一般包含病毒衣壳蛋白(Coat Protein, CP)和一些非结构性基因,RNA2基因组可以被修饰以携带外源基因,因此目的基因通常被克隆到RNA2上[1]. 烟草脆裂病毒不仅能够侵染烟草,还能够侵染番茄果实、辣椒叶、矮牵牛花、耧斗菜、唐松草花、玫瑰花、草莓果实、加州罂粟、麻疯树、紫茉莉[2]和菠菜[3].
在植物油脂基因工程中,常用传统的转基因技术,即将特定外源基因稳定的在受体生物中表达[4],但实验过程繁琐,周期较长,而利用烟草农杆菌介导的瞬时系统仅在被侵染叶片表达目的基因[5]. TRV作为一种植物病毒载体,具有复制快,感染强,对植株伤害小等优点,不仅能在侵染叶片表达目的基因,在未侵染叶片也能检测出目的基因的表达[6].
研究发现WRI1是提高总油脂含量的关键基因[7],而FAD2基因是单不饱和脂肪酸生成多不饱和脂肪酸的关键[8].而多不饱和酸不利于储存,沉默FAD2基因可以减少多不饱和脂肪酸的含量[9]. 本研究通过构建TRV-WRI1过表达载体和TRV-FDA2沉默载体,对侵染7天和14天的本氏烟草株系WRI1下游基因进行定量分析,并测定植株叶片的脂肪酸含量和种类,探讨利用TRV过表达系统进行油脂相关基因功能研究的可行性.
2 材料与方法
2.1 材 料
2.1.1 植物材料 本氏烟草(Nicotianabenthamiana)种子为本实验室保存.
2.1.2 菌株和质粒 大肠杆菌(E.coli)和农杆菌(Agrobacteriumtumefaciens)GV3101菌株于-80 ℃冰箱保存. 烟草脆裂病毒过表达及沉默载体菌株于-80 ℃冰箱保存.
2.1.3 PCR引物 见表1.

表1 本文中所用PCR引物
2.2 方 法
2.2.1WRI1和FAD2基因序列克隆和载体构建 以拟南芥RNA经反转录的cDNA为模板,利用高保真酶Primer STAR Max(TaKaRa, Japan)进行WRI1和FAD2基因的扩增,引物序列如表1所示. 利用TaKaRa公司的限制性核酸内切酶对TRV载体的TRV2质粒和PCR产物进行双酶切. 酶切反应结束后用核酸纯化回收试剂盒回收酶切产物,然后进行连接. 测序后转入农杆菌GV3101.
2.2.2 农杆菌重悬液制备 将转入TRV-WRI1和TRV-FAD2的农杆菌接种到含有Kana、Rif抗性的培养基中,28 ℃,200 r/min活化24 h;取1 mL活化菌液加入 YEB培养基,28 ℃,200 r/min,12 h,直至OD600≈0.8~1.0;5 000 r/min,常温离心富集农杆菌菌体;用重悬液(1 mol/L MgCl2,1 mol/L MES,200 mmol/L AS)重悬菌体至OD6000.8~1.0,将TRV2菌液与TRV1菌液按照1∶1体积混匀,室温避光静置2~4 h.
2.2.3 侵染本氏烟草 选择4~6叶期的本氏烟草,将农杆菌重悬液注射扩散到膨大叶的远轴侧. 每株植物注射2~3片叶,将侵染后暗室培养24 h后,置于25 ℃,16 h/8 h光周期温室培养.
2.2.4 qRT-PCR分析 利用天根生物RNAPrep Pure Plant Kit试剂盒提取本氏烟草总RNA,PrimeScriptTMRT reagent Kit (TaKaRa, Japan)试剂盒合成单链cDNA. 使用SYBR®Premix Ex TaqTMⅡ试剂对WRI1下游基因进行qRT-PCR扩增,所用引物见表1.
2.2.5 油脂成分测定 称取0.1 g侵染后的本氏烟草叶片或侵染后的系统叶叶片放入玻璃管中,加入1 mL新鲜配制的5%硫酸甲醇溶液(v/v). 加入300 μL甲苯作为促溶剂,再加入10 μL 2%的BHT溶液和10 μg十七烷酸甘油三酯溶液作为内参;各试剂加入结束后迅速震荡混匀,90~95 ℃水浴反应1.5~2 h;取出玻璃管冷却至室温,加入1.5 mL新鲜配制的0.9% NaCl溶液,混匀. 再向玻璃管中加入1 mL正己烷萃取脂肪酸甲酯,震荡混匀,静置5 min,取上清液于2 mL棕色玻璃瓶中;用气相色谱与质谱联用仪GC-MS 2010对提取的脂肪酸甲酯含量进行测量,分离柱为HP-88. GC条件为:火焰离子化检测器和加样器温度为220 ℃. 炉温升温程序为:160 ℃,保持1 min;然后每分钟升高10 ℃,升到200 ℃,保持1 min;然后每分钟升温5 ℃,升到210 ℃,保持1 min. 脂肪酸甲酯含量通过与内参的峰面积对比进行计算.
3 结果与分析
3.1 TRV-WRI1过表达载体及TRV-FAD2沉默载体构建
用表1中设计的引物进行扩增,凝胶电泳检测出WRI1大小约1 300 bp,FAD2大小约为500 bp(图1),片段大小与预期结果相符.
图1 WRI1和FAD2基因PCR产物电泳图
3.2 TRV-WRI1过表达植株中下游基因表达量分析
用TRV-WRI1过表达载体的农杆菌侵染本氏烟草植株后,分别在侵染后第6 d和第13 d提取系统叶叶片的总RNA,以烟草β-actin为内参,利用qRT-PCR技术检测WRI1转录因子相关基因的表达量. 本研究对ACP1,KAS1,BCCP2,Pkb1和SUS2基因进行定量分析,实验结果表明,无论是在侵染叶还是系统叶中,过表达株系的这5个相关基因的表达量均显著高于对照组;第6天的侵染叶中ACP1基因增加量最为明显,与对照组相比增加来约3倍;侵染第13 d的系统叶中,ACP1基因和SUS2基因表达量增加最明显,增加了约1倍和1.5倍. 总体而言,侵染叶中WRI1相关基因的相对增加幅度略高于系统叶中相关基因的相对增加幅度(图2). TRV-WRI1过表达后,在转录水平初步启动了植物中与脂肪酸合成相关基因的表达.

图2 TRV-WRI1过表达植株叶片中WRI1下游基因表达分析
3.3 TRV-WRI1过表达植株中油脂含量和成分的测定
为进一步探究WRI1基因对植物油脂含量积累的影响,本研究将TRV-WRI1和TRV空载(CK)侵染本氏烟草,分别在第7 d和第14 d取侵染叶叶片、系统叶叶片,测定油脂的成分和含量. 结果显示,在侵染后第7 d,实验组中的软脂酸含量和大部分硬脂酸含量显著增加. 以每克植物叶片中的油脂含量来分析,实验组中的总油脂含量比对照组约增加16%. 侵染14 d后与对照组相比,实验组中的部分硬脂肪酸含量显著增加,总油脂含量比对照组约增加28%(图3).
3.4 TRV-FDA2沉默植株中油脂含量的成分和测定
为进一步探究FAD2基因对植物油脂含量积累的影响,本研究将TRV-FDA2和TRV空载(CK)侵染本氏烟草,分别在第7 d和第14 d取侵染叶叶片、系统叶叶片,测定油脂的成分和含量. 结果显示,在侵染后第7 d和第14 d,实验组中的多不饱和脂肪酸含量显著减少. 以每克植物叶片中的油脂含量来分析,实验组中的多不饱和脂肪酸含量比对照组减少约25%. 在侵染后第14 d,与对照组相比,实验组中的多不饱和脂肪酸含量显著减少,相比对照组减少约24%(图4).

图3 TRV-WRI1过表达植株叶片中脂肪酸分析
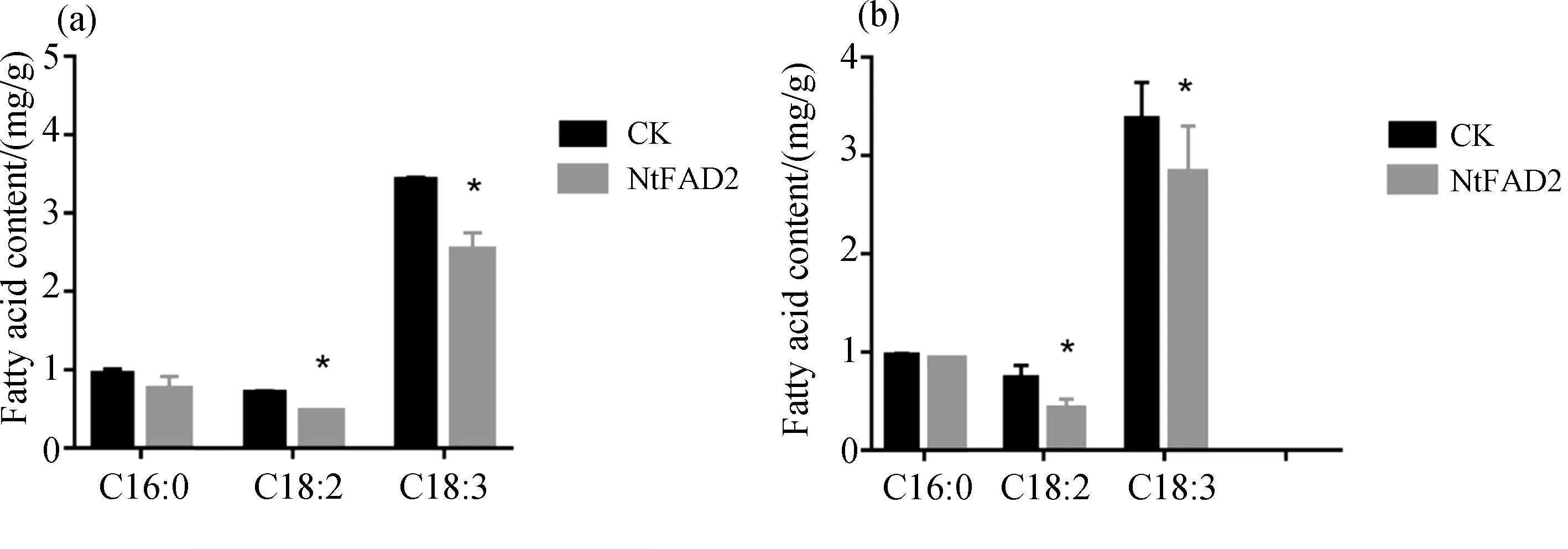
图4 TRV-FDA2沉默植株叶片中脂肪酸分析
种子油脂合成[11-12]. 利用TRV过表达系统,在本氏烟中过表达WRI1基因,随着侵染后时间的延长,外源基因短期内在本氏烟中大量表达,在侵染后的第6 d和第13 d对WRI1的下游靶基因进行定量分析,实验结果表明,WRI1的过表达启动了其下游靶基因的表达,过表达植株中下游靶基因的含量显著高于对照组. 从qRT-PCR结果可知,系统叶中WRI1下游基因的总体增加水平没有侵染叶中的高,特别是与对照组相比,系统叶中的BCCP1、KAS1和Pl-Pkβ1基因的上升相对值没有侵染叶中的高.
对其脂肪酸含量进行测定,总的侵染叶片脂肪酸含量第7 d的增加16%,第14 d系统叶增加28%. 与对照组相比,侵染叶中的软脂酸、硬脂酸和大部分不饱和硬脂酸均呈显著性增加,而系统叶中的软脂酸和部分不饱和硬脂酸的含量仅呈上升趋势,并没有出现显著性的差异. Yang等[13]在二穗短柄草中过表达BdWRI1诱导了糖酵解和脂肪酸生物合成相关基因的转录,使8 w龄叶片中的油脂含量增加了32.5倍,促进了叶片中脂肪酸的合成和标记的积累,然而,BdWRI1的过表达也导致了游离脂肪酸含量的增加,具有细胞毒性作用,导致细胞死亡.同时,利用TRV病毒载体沉默FDA2基因,脂肪酸测量结果显示,在第7 d和第14 d,与对照组相比,多不饱和脂肪酸分别下降了25%和24%,且饱和脂肪酸含量差异不显著,本实验成功利用病毒载体沉默FDA2基因.
病毒载体瞬时侵染植物表达目的基因相比于稳定表达系统在时间上大大缩短,本实验中,仅第6 d就检测到了目的基因的表达,本氏烟中脂肪酸含量得到增加. 但是病毒载体的承载容量有限,不能包装大片段的外源DNA基因,复制稳定性差,外源基因无法在植物细胞中稳定遗传[14-15];同时TRV病毒载体侵染的物种也有局限性,但是可以通过开发更多的病毒载体来解决这一问题. 后续研究可以同时在烟草叶片上利用TRV病毒载体,过表达WRI1和沉默FAD2基因,达到提高油脂总量和减少不饱和脂肪酸比例目的.
