中山杉406ThSHR3 基因的克隆、表达及蛋白互作研究
2021-12-27王芝权殷云龙华建峰
宣 磊,王芝权,殷云龙,华建峰
(江苏省中国科学院植物研究所,江苏省落羽杉属树木种质创新与繁育工程研究中心,江苏南京 210014)
中山杉(Taxodiumhybrid‘Zhongshanshan’)是江苏省中国科学院植物研究所从落羽杉属树木杂交组合中选育出的,是具有一定超亲性状的优良无性系总称。中山杉具有生长迅速、抗逆性强、景观性好、成材率高等特点,已在我国沿海防护林建设、公路及城乡绿化、农田林网和滩涂造林等方面得到广泛应用[1]。目前,中山杉无性系主要通过嫩枝扦插的方法进行繁殖,近些年林业工作者一直致力于通过中山杉扦插技术改良来提高中山杉的生根率,研究表明:泥炭土、珍珠岩及沙壤土组成的混合基质有利于大多数中山杉无性系生根[2]。植物生长调节物质的浓度对中山杉扦插生根率影响显著,2000mg·L−1IAA和2000mg·L−1NAA 混合液处理的中山杉无性系插条,生根率和生根数量都显著提升。然而,在中山杉长期选育过程中发现,不同无性系间生根能力差异显著,即使优化扦插技术,中山杉302 的平均生根率(57.50%)仍显著低于中山杉118(83.93%)和406(87.14%)[3-4],且随着无性系生理年龄的增长,早期选育出的无性系生根力出现衰退现象[1],中山杉的生根机理亟需深入研究。
SHR(SHORT-ROOT)是植物GRAS 家族中与根系发生及形态建成密切相关的一个分支,对根尖干细胞微环境的特化和维持起着关键作用,它调控根皮层、内皮层初始细胞及初始细胞子细胞的垂周和平周不均等分裂,影响植物根的辐射式生长模式[5-7]。拟南芥shr突变体植株根尖分生组织的基本组织子细胞不能发生不对称平周分裂,仅产生一层类似皮层的细胞[8],shr突变体植株表现出主根生长减弱,侧根数量减少,植株矮小,子叶颜色深暗等一系列表型。过量表达SHR基因,根尖形态变化显著,基本组织发生大量的平周分裂,产生多个细胞层[9]。同时,SHR基因是一个功能进化比较保守的基因,目前已在玉米(Zea maysLinn.)[10]、水稻(Oryza sativaLinn.)[11]、毛果杨(Populus trichocarpaTorr.&Gray.)[12]、辐射松(Pinus radiataD.Don)[13]等植物中发现的SHR 家族成员均和根的生长发育息息相关。研究表明:SHR 转录功能的行使,通常需要依靠其下游另一个GRAS家族成员SCR(SCARECROW)[14]。SCR 与SHR 形成SCR/SHR 复合体共同激活下游基因表达,SCR/SHR 复合体直接作用于CYCD6.1(Dtype cyclin 6.1)蛋白,CYCD6.1 是细胞发生平周分裂的标志性蛋白,保证根尖细胞会在早期和晚期分别自发地进行2 次平周分裂,产生完整的基本组织[15-17]。国内外学者对于不同植物SHR基因的挖掘以及SHR对根部细胞分裂调控机制的解析一直处于探索之中,本研究在前期已获得2个中山杉SHR基因(ThSHR1,ThSHR2)的基础上[18],新筛选到1个在中山杉406 不定根生长期高表达的基因ThSHR3,而ThSHR3的生物学功能尚不清楚,拟通过生物信息学手段对其进行系统进化分析,探索其在中山杉不定根发育过程中表达模式,对ThSHR3 蛋白进行亚细胞定位以及互作研究,推测其可能存在的信号通路。以期为中山杉以及落羽杉属植物不定根发育的分子机理研究提供新的理论依据。
1 材料和方法
1.1 材料和试剂
“中山杉406”的嫩枝扦插试验于2019年7月中旬开展于江苏省中国科学院植物研究所中山杉苗圃内。插穗长度约10~15 cm,扦插基质为珍珠岩和泥炭土,体积比例为1:1,光周期为14 h 的光照周期和10 h 的暗周期。根据中山杉406 插穗基部明显的形态学改变,在4个时间点取样:0 d 皮层休眠期(S0),21 d 愈伤组织形成期(S1),35 d初生根形成期(S2)和56 d 根系生长期(S3)[18]。其中,皮层休眠期和愈伤组织形成期的取样部位为插穗基部皮层组织(0.3 cm 左右),初生根形成期和根系生长期的取样部位为根系。
RNeasy Plant Mini Kit 及Plant Genomic DNA Kit 购自QIAGEN 公司;PrineScript @ RTase 逆转录试剂盒、3′-Full RACE 和5′-FullRACE 试剂盒、LAtaq 酶、rtaq 酶均购自TaKaRa 公司;载体构建使用的GATEWAY 技术及相关试剂均来自Invitrogen公司。
1.2 基因克隆
使用RNeasy Plant Mini Kit 试剂盒提取中山杉406 根总RNA,确保RNA 浓度为1 ng·μL−1,并通过2%琼脂糖凝胶电泳检测RNA 的完整性。使用PrineScript @ RTase 逆转录试剂盒合成中山杉406 根cDNA,根据前期不定根转录组数据,筛选到605 bp 的目的基因ThSHR3的片段(CL8931.Contig2),通过PCR 扩增验证目的基因序列,PCR 扩增体系如下:cDNA 模板1.0 μL,TaKaRa LA Taq (5 U·μL−1)0.5 μL,10×LA PCR Buffer (Mg2+Free) 5.0 μL,MgCl2(25 mmol·L−1)5.0 μL,dNTP Mixture(各2.5 mmol·L−1)8.0 μL,正向引物(10 μmol·L−1) 2.0 μL,反向引物(10 μmol·L−1) 2.0 μL,加超纯水补至50.0 μL。PCR扩增程序为:94℃ 3 min;94℃ 30 s,56℃ 30 s,72℃ 2 min,35个循环;72℃ 10 min。进一步参照TaKaRa 公司3′和5′-Full RACE 试剂盒说明进行巢氏PCR 扩增,完成ThSHR3基因全长克隆,其中,涉及到的引物参考(表1)。将扩增片段序列进行序列比对、拼接,最终获得了ThSHR3基因的全长cDNA 序列。利用BioXM 软件预测ThSHR3基因的开放阅读框(ORF),进一步通过PCR 验证。

表1 引物序列Table 1 Primer sequences
1.3 生物信息学分析
利用美国国家生物信息中心(NCBI)在线BLAST 软件比对分析ThSHR3的DNA 和蛋白质序列。使用在线程序Expasy Protparam 计算蛋白质的理论等电点(pI)、分子量(MW)和氨基酸组成。通过PROSITE 以及GORIV 二级结构检测程序分别预测蛋白质的结构域及二级结构。使用ClustalX2软件将ThSHR3 的蛋白序列与其他植物已公布的SHR 蛋白序列进行序列多重比对。利用NLStradamus程序预测ThSHR3 蛋白是否具有核定位信号。通过MEGA 7.0 软件,选用最大似然法构建系统发育树,自举检测1 000 次。
1.4 实时定量及半定量PCR
以中山杉406 不定根发育的4个时间点S0~S3为材料,根据已验证的cDNA 序列设计定量引物(表1),以中山杉的APRT基因为内参基因[19],采用半定量PCR 和荧光定量PCR 分别对ThSHR3进行表达分析检测。半定量PCR 反应体系及程序参照rtaq 酶说明书(https://www.takarabiomed.com.cn/)。选择Analitik Jena qTOWER2.2 PCR 系统(Biometra,德国)进行荧光定量RT-PCR(qPCR)。反应程序设定为:50℃ 2 min;95℃ 10 min;40个循环:95℃ 15 s,60℃ 1 min;通过从60℃到95℃加热扩增产物,获得熔解曲线。反应体系共20 μL,其中包括2μL稀释后的cDNA,10 μL FastStart Universal SYBR Green Master(Rox,德国),10 pmol正向引物,10 pmol 反向引物以及灭菌去离子水,每个样品设3 次技术重复,采用2−ΔΔCT法进行相对定量的分析[20]。
1.5 载体构建和原生质体瞬时表达
表达的载体构建利用Gateway Technology(Invitrogen,美国)技术,先将不包含终止密码子的ThSHR3ORF 序列连接到入门载体pCR8/GW/TOPO 上,再使用LR 酶将入门载体连接到包含绿色荧光蛋白(GFP) 标签的目的载体p2GWF7 上。同理,使用相同的方法构建ThSHR3 及ThSCR 的蛋白互作载体,目的载体为含有黄色荧光蛋白(YFP)标签的pUC-SPYNE、pUC-SPYCE。最终重组表达载体有:35S::ThSHR3-GFP、ThSHR3-YFPN、ThSHR3-YFPC、ThSCR-YFPN、ThSCR-YFPC。利用PEG 介导转化法将测序正确的融合表达载体导入杨树叶肉原生质体中[21]。利用BX51 荧光显微镜(Olympus,日本)观测样品中荧光表达情况。
2 结果与分析
2.1 ThSHR3 基因克隆及生物信息学分析
根据中山杉不定根发育的转录组及蛋白组数据,利用RACE 巢式PCR 扩增,拼接得到ThSHR3基因的全长序列。结果表明:ThSHR3基因的cDNA 全长为2 019 bp,包含1个1 446 bp 的开放阅读框(ORF),5′端非编码翻译区(UTR)长度为354 bp,3′端非编码翻译区(UTR)长度为219 bp。ThSHR3基因编码482个氨基酸残基。ExpasyProtparma 预测ThSHR3 分子量和等电点分别为66.082 和5.17。GOR IV 的分析结果显示:ThSHR3 含有30.03%α 螺旋(Hh),15.77% 延伸链(Ee),54.19% 的无规则卷曲(Cc),不含有beta 转角(Tt)。NLStradamus 程序预测ThSHR3 蛋白主要定位在细胞核上。Prosite 分析表明:ThSHR3蛋白具有保守的GRAS 结构域。
2.2 ThSHR3 基因同源性及系统进化分析
将ThSHR3 的蛋白质序列与TAIR 中的拟南芥AtSHR(At4g37650.1) 蛋白序列和杨树PeSHR1、PeSHR2、PeSHR3 蛋白序列[22],中山杉ThSHR1(MF045148)、ThSHR2(MF045149)蛋白序列进行多重序列比对。结果表明:ThSHR 蛋白的N 端不保守,而C 端相对比较保守,和其他物种的SHR 蛋白一样,ThSHR 蛋白包括GRAS 家族成员特有的LHRI、VHIID、LHRII、PFYRE 和SAW基础序列(图1)。
将ThSHR3 的氨基酸序列与其他物种中已公布的45个较典型的GRAS 蛋白的氨基酸序列进行系统进化树分析。结果表明:GRAS 蛋白家族可划分为具有不同的特征的SHR、DELLA、PATl、SCL9、SCR、LAS/SCL18、SCL4/7、HAM 的8个分支[9](图2)。ThSHR3 被划分到SHR 分支,和其他物种的SHR 蛋白聚为一类,ThSHR3 和中山杉ThSHR1 亲缘关系最近,同源性均为81%,和松科植物PmSHR(QCU71495.1)及辐射松PrSHR(ABW20412.1)的亲缘关系也较近(图2)。
2.3 ThSHR3 基因表达分析
本研究利用半定量PCR 和荧光定量PCR 方法,检测ThSHR3基因在中山杉406 不定根皮层休眠期、愈伤组织形成期、初生根形成期和根系生长期4个时期的动态表达情况,内参基因均为APRT基因。结果表明:ThSHR3基因在中山杉不定根4个不同发育阶段均有表达。ThSHR3表达呈现持续上升的表达模式,在根系生长期表达量最高,在皮层休眠期表达量最低,最高表达量是最低表达量的28.4 倍,且半定量PCR 和荧光定量PCR结果基本一致(图3)。
2.4 ThSHR3 蛋白的亚细胞定位
本研究以绿色荧光蛋白作为报告基因,在前期已经成熟的杨树叶肉原生质体瞬时表达体系的基础上[21],将中山杉重组融合表达载体导入杨树叶肉原生质体中。经过18 h,23℃暗培养,观察可得GFP 标签在488 nm 蓝光激发下产生了509 nm 的绿色荧光,阳性对照35S::GFP 在细胞核、细胞质、细胞膜等区域中都产生了明显的绿色荧光信号,35S::ThSHR3-GFP 融合蛋白仅在细胞核区域产生绿色荧光,表明中山杉ThSHR3基因所编码的蛋白质定位于细胞核,这和前期在线程序预测结果一致,也符合其作为转录因子的特性(图4)。
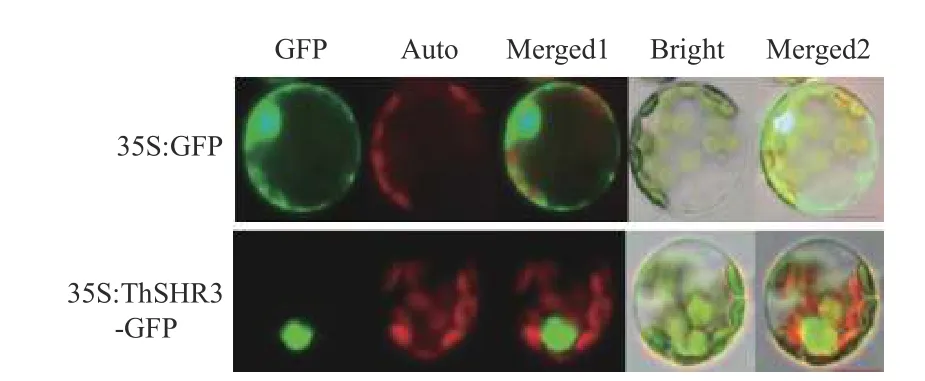
图4 ThSHR3 在杨树叶肉原生质体内的亚细胞定位Fig.4 Subcellular-localization of ThSHR3 in Populus mesophyll protoplasts
2.5 ThSHR3 与ThSCR 的蛋白互作
双分子荧光互补(BiFC)是目前用于检测细胞体内蛋白质互作的一项成熟技术,本研究利用BiFC 技术检测中山杉的ThSHR3 与ThSCR 蛋白的互作情况,前期已经证明ThSHR3 及ThSCR 亚细胞定位结果均定位在细胞核[23]。3 次重复性实验结果显示:ThSHR3-YFPN与ThSCR-YFPC、ThSHR3-YFPC与ThSCR-YFPN这2个组合的YFP 荧光蛋白重新恢复活性,在细胞核处观察到清晰的荧光信号,2 组对照组合均无荧光信号产生(图5),表明ThSHR3 与ThSCR 在细胞核发生了互作。
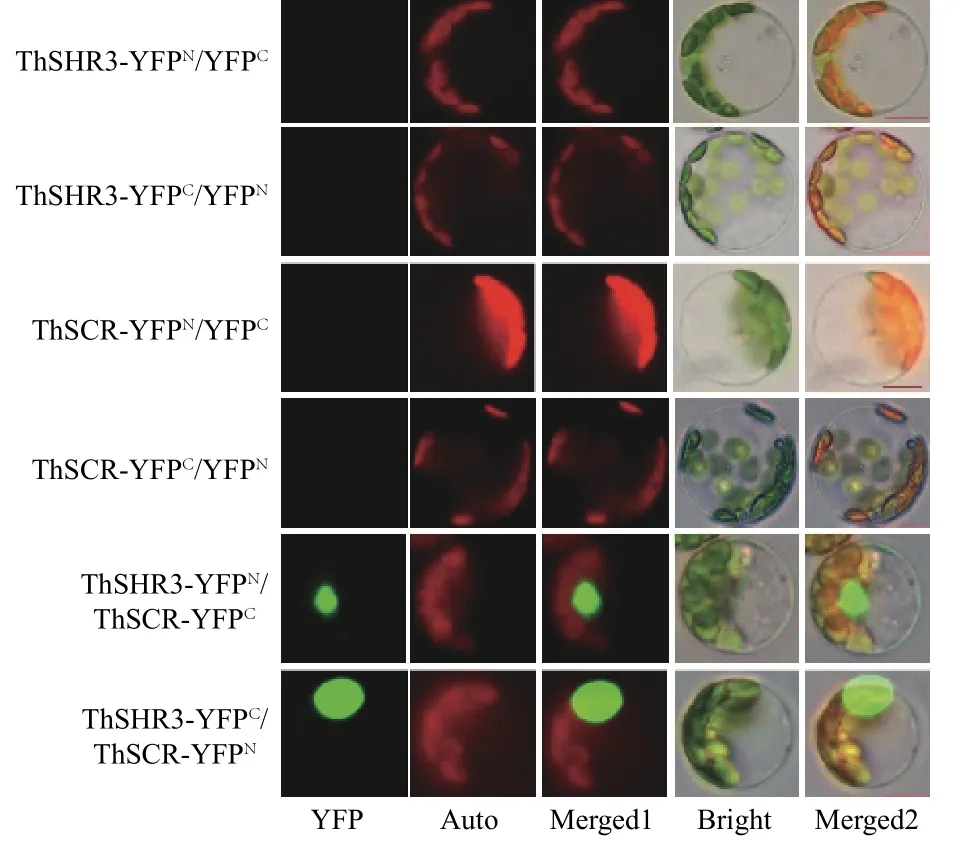
图5 ThSHR3 和ThSCR 蛋白互作Fig.5 ThSHR3 and ThSCR interaction by BiFC
3 讨论
根系是植物长期适应陆地生境而形成的重要器官,根系从土壤中吸收水分和矿质营养,并对植物起机械支撑作用[24]。2000年,Helariutta等首次在拟南芥根中发现了转录因子SHR[8],证明其参与根尖辐射形态建成,在基本组织细胞平周分裂形成过程中发挥着的重要作用,进而其他物种的SHR基因陆续被挖掘研究[25]。本研究成功克隆获得了中山杉406ThSHR3基因全长。经过氨基酸序列比对和系统进化分析发现,ThSHR3和中山杉ThSHR1、ThSHR2高度同源,和其他物种的SHR基因一起划分到SHR 分支,且具有植物GRAS 蛋白家族特有的结构域,因而推测ThSHR3可能参与中山杉406不定根的发育。
木本模式植物杨树的SHR分支基因包含3个成员:PeSHR1、PeSHR2、PeSHR3,其表达存在着2种模式,PeSHR1、PeSHR2基因表达量在根的发育过程中呈逐渐上升趋势,根发育到4 周时表达量最高,而PeSHR3基因在根中的表达模式为先上升后下降[22]。中山杉3个SHR基因的表达特性在不定根不同发育阶段是否也存在着差异?研究表明:ThSHR1呈逐渐上升的表达趋势,在根的生长期表达量最高,而ThSHR2表达量先上升再下降,在初生根形成期表达量最高[23],中山杉ThSHR3和ThSHR1的表达趋势基本一致,且聚类分析发现二者的同源性较高,推测二者的功能可能更相近。SHR基因家族成员不同的表达模式,暗示着它们可能在根系发育过程中行使着不同的生物学功能,但也不排除它们之间存在功能冗余性,有待通过遗传转化等方法进行深入探究。
染色体免疫共沉淀及酵母双杂交实验表明,拟南芥中SCR 与SHR 互作区域在LHRI-VHIID-LHRII基序之间[15]。水稻的SHR 与SCR 蛋白也存在着互作,然而水稻的2个SHR基因OsSHR1和OsSHR2,仅发现OsSHR1可与OsSCR发生互作[11]。目前,关于SHR 和SCR 及其互作基因调控根部细胞不均等分裂的研究,主要集中在SHR/SCR/RBR/CYCD6-1-CDK 这信号通路中。SHR/SCR/RBR/CYCD6-1-CDK 是一条双稳信号回路,细胞周期蛋白CYCD6及其依赖性激酶1-CDK 共同磷酸化RBR(RING Between RING)蛋白,进而影响SCR 蛋白活力,同时CYCD6 转录活性又受到SHR-SCR 复合物共同的转录调控,此通路中SHR-SCR 复合物的活性适中,才能保证根部细胞不均等分裂有序完成[15]。胥猛等利用BiFC 技术对杨树SHR/SCR/RBR/CYCD6-1-CDK 信号通路基因的互作进行研究,发现仅PeSHR1 与PeSCR、PeSHR1 与PeCYCD6、PeSCR与PeCYCD6 三个组合发生了互作[22,26]。中山杉SHRSCR 信号通路的研究尚未开展,本研究对中山杉中新挖掘出的ThSHR3 进行了蛋白互作研究,BiFC实验发现中山杉ThSHR3 可以和ThSCR 发生明显互作,推测ThSHR3 可能和ThSCR 存在于同一信号通路中,通过形成ThSHR3/ThSCR 复合体来行使转录功能;而ThSHR 分支上的其它成员之间以及ThSHR1、ThSHR2 与ThSCR 的互作情况仍然未知,具体的作用机制仍需通过分子生物学手段进一步深入解析。
4 结论
本研究从中山杉406 扦插苗不定根中克隆获得了ThSHR3基因,该基因编码的蛋白具有GRAS家族成员特有的保守结构域,且从属于SHR 亚家族。ThSHR3基因在中山杉皮层休眠期、愈伤组织形成期、初生根形成期、根系生长期4个时期中呈逐渐上升的表达趋势。双分子荧光互补实验显示:ThSHR3 蛋白和ThSCR 蛋白存在着明显互作,而SHR-SCR 信号通路在植物根系发育过程中发挥着关键调控作用,表明ThSHR3 为中山杉不定根发育相关的重要转录因子,为进一步探究中山杉以及落羽杉属树木不定根发育的分子机理提供了理论基础。
