朱红毛斑蛾繁殖行为的研究
2021-12-27吴海盼刘俊延王小云郑霞林
吴海盼,刘俊延,王小云,陆 温,郑霞林
(广西农业环境与农产品安全重点实验室,广西大学农学院,广西南宁 530004)
斑蛾总科(Zygaenoidea )包括斑蛾科(Zygaenida)、毛斑蛾科(Phaudidae)和绒蛾科(Megalopygidae)等13 科2600 余种,其中斑蛾科和毛斑蛾科中的许多种类是农业、园艺和林业上的重要害虫[1-3]。目前,针对斑蛾总科害虫中有关繁殖方面的研究主要集中性冲突/选择[4]、延迟交配[5]、性信息素[6-8]和滞育[9]等方面。尚未见有关毛斑蛾科昆虫繁殖行为方面的研究报道。
朱红毛斑蛾(Phauda flammans)(鳞翅目Lepidoptera:毛斑蛾科Phaudidae)是榕树上的一种常暴发的食叶害虫,其危害不仅影响榕树的生长发育,还影响城市景观和生态效应[10]。该虫广泛分布于东南亚和南亚诸国[11-17],且已明确了其生物生态学特性,包括形态特征[10,18-19]、寄主选择[20]、年生活史[10]、发育起点温度和有效积温[21]、抗寒力[22]、天敌种类[23],越冬虫态的调查方法[24]和防治技术[8,25-27]等。目前,关于朱红毛斑蛾的繁殖行为尚无相关研究报道。鉴于此,本研究研究了朱红毛斑蛾的羽化、求偶、交配和产卵行为及节律,以期明确朱红毛斑蛾的繁殖行为,为该虫的种群管理提供基础资料。
1 材料与方法
1.1 供试虫源
在广西大学校园内(108°29′ E,22°85′ N)的小叶榕上采集6 龄朱红毛斑蛾幼虫,带回室内置于圆形塑料盒(底部直径20.5 cm,顶部直径25.0 cm,高15.0 cm;10 头·盒−1)内,在温度26 ± 2℃、相对湿度70% ± 10%、光周期为16 L:8 D 的环境条件下饲养,每日更换新鲜小叶榕叶片直至化蛹,收集茧(蛹)待用。
1.2 研究方法
1.2.1 朱红毛斑蛾羽化行为的研究 将茧(蛹)置于圆形塑料盒内(底部直径18.5 cm,顶部直径23.0 cm;高10 cm;100 头·盒−1),在上述饲养条件下,人工连续观察和记录塑料盒中朱红毛斑蛾蛹的羽化情况,每隔1 小时统计羽化的雌虫(n=493)和雄虫(n= 536)数量并将其取出,直至无成虫羽化为止,分析成虫的羽化行为、日节律及昼夜节律。
1.2.2 朱红毛斑蛾求偶行为的研究 将雌雄蛹分开保存在不同的塑料盒内,置于上述饲养条件下。待成虫羽化后,随机选取1 日龄健康雌雄虫放入装有盆栽小叶榕的网笼(长×宽×高= 100 cm×100 cm×180 cm)中(100 对·笼−1),人工观察和统计每小时雄虫求偶(雄虫求偶时,腹部翘起几乎与地面垂直,并快速振翅)的数量(n= 1299),并将统计过的雄虫移出笼外,避免重复计数。
1.2.3 朱红毛斑蛾交配行为的研究 随机选取300 对1 日龄健康雌雄虫放入网笼内,在上述饲养条件下人工观察,发现交配时将交配的雌雄虫轻轻取出并置于塑料盒中(1 对·盒−1),统计每小时内的交配数,并分析其交配行为和昼夜节律。对置于塑料盒内交配中的朱红毛斑蛾雌雄虫采用数码摄像机(DS-2CD3T45D-13,海康威视数字技术股份有限公司,杭州,浙江)连续拍摄,直至其结束交配,统计其交配时长。
1.2.4 朱红毛斑蛾产卵行为的研究 随机选取交配结束后的朱红毛斑蛾雌虫置于塑料盒中(1 头·盒−1),并给以新鲜小叶榕枝条,在上述饲养条件下,使用数码摄像机连续拍摄直至雌虫死亡,记录雌虫每天每小时内的产卵情况,分析其产卵行为和昼夜节律,并统计每头雌虫(n= 202)的总产卵量。每天收集榕树叶上的卵块置于垫有湿润滤纸的培养皿内(直径= 12 cm),统计其在上述饲养条件下的孵化率。
1.3 数据分析
使用SPSS 16.0(SPSS Inc.,Chicago,IL,USA)软件对24 h 内不同时间段雌虫的产卵率进行数据统计分析,采用ANOVA 单因素方差分析,并用Tukey’s HSD 进行多重比较(P<0.05)。统计分析前,百分率数据进行平方根转换。
2 结果与分析
2.1 朱红毛斑蛾的羽化行为
2.1.1 朱红毛斑蛾的羽化行为特征 朱红毛斑蛾羽化前,蛹体渐变为深红色。近羽化时蛹体轻微扭动,随后频率逐渐加快,约3~5 min 后蛹的头部先钻出茧室一侧留下的稀薄羽化孔,然后虫体继续抖动,头胸部先钻出蛹壳,通过胸足的不断爬行利用茧室羽化孔的摩擦逐步褪去腹部蛹壳,蛹壳留于蛹室内。成虫爬出茧室后,在蛹室附近静伏,并用前足梳理触角,数分钟后成虫会通过其排泄孔射出棕黄色排泄物。待完全展翅后,双翅竖立数分钟,随后飞行离开(图1)。

图1 朱红毛斑蛾的羽化行为Fig.1 Emergence of Phauda flammans adult
2.1.2 朱红毛斑蛾的羽化节律 朱红毛斑蛾雌雄成虫仅在光期06:00—16:00 羽化,在11:00—13:00达到羽化高峰期,且雌虫羽化时间比雄虫提早约1 h,雌雄成虫的昼夜羽化节律均呈“单峰”型。采集的同一批样本中,朱红毛斑蛾化蛹持续15 d,蛹期11~17 d,雌雄成虫的羽化持续19 d,雌虫早于雄虫约2 d 羽化,提早1 d 结束。雌雄虫的日羽化高峰期均出现在开始羽化后的8~11 d,在羽化高峰期的羽化率分别占雌雄成虫总羽化率的59.35%和57.01%,雌雄虫的日羽化节律均呈“单峰”型(图2)。

图2 朱红毛斑蛾成虫的羽化节律Fig.2 Emergence of Phauda flammans adults
2.2 朱红毛斑蛾的求偶行为
2.2.1 朱红毛斑蛾的求偶行为特征 朱红毛斑蛾雌雄虫仅在光期求偶。雌虫羽化后,大多数个体顺着树干爬至寄主植物枝干或叶片上,也有少量雌虫通过飞行抵达寄主植物。雌虫在寄主植物上寻找到合适的位置后,开始静止等待雄虫,在此过程中,常见雌虫无规律快速震颤腹部,偶见腹部末端翘起现象。与此同时,雄虫飞行搜索雌虫。当飞抵雌虫附近时,雄虫继续爬行,并伴随触角快速摆动侦测雌虫位置。待雄虫准确定位雌虫后立即表现出明显的求偶行为,包括雄虫触角不停敲击雌虫触角和身体,雄虫在雌虫周围快速振翅,腹部高高翘起几乎垂直于地面,然后暴露阳茎试图与雌虫交配。雌虫若接受雄虫的求偶,则立即接受交配;雌虫若不接受该雄虫的求偶,则快速爬行以挣脱雄虫,而雄虫则穷追不舍,并重复上述求偶行为,试图再次与雌虫进行交配,若雌虫接受该雄虫的再次求偶则立即交配,否则雌虫飞走,雄虫需重新寻找和定位新的雌虫(图3)。
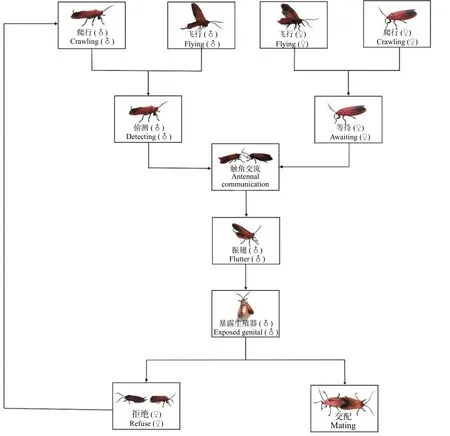
图3 朱红毛斑蛾的觅偶和求偶行为Fig.3 Mate searching,courtship and calling behaviors of Phauda flammans
2.2.2 朱红毛斑蛾求偶节律 朱红毛斑蛾从09:00—18:00 均有个体求偶,求偶高峰期为11:00—16:00,占求偶个体总数的69.44%(图4)。
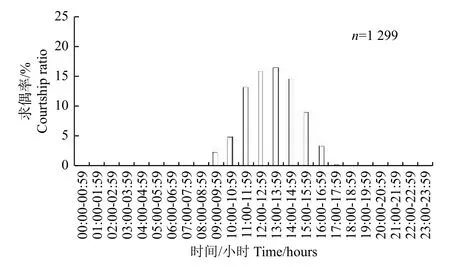
图4 朱红毛斑蛾求偶的昼夜节律Fig.4 Circadian rhythm of courtship of Phauda flammans
2.3 朱红毛斑蛾的交配行为
2.3.1 朱红毛斑蛾的交配节律及行为求偶率 朱红毛斑蛾的交配行为在10:00—18:00 均可发生,14:00—16:00 为交配高峰期,暗期未见交配行为。朱红毛斑蛾交配姿势多为“一”或“V”字型(图5),交配地点多在寄主植物叶背和枝干,交配时长为16.16 ± 0.26 h(n= 190),交配率63.33%,雌雄成虫均一生只交配1 次,为单配制蛾类。

图5 朱红毛斑蛾交配行为Fig.5 Mating postures of Phauda flammans
2.4 朱红毛斑蛾的产卵行为
2.4.1 朱红毛斑蛾的产卵行为特征 朱红毛斑蛾雌虫在交配结束约1 h 或次日光期即可产卵。产卵前,雌虫常在寄主植物上爬行,并用触角不停地触碰寄主植物以搜寻合适的产卵地。观察发现,卵多产于寄主植物的叶柄或叶尖部位。产卵时,腹部末端对准产卵地,不断抖动腹部,卵从腹部产卵孔排出,卵粒粘结呈线状或鳞片状,无重叠(图6)。每头雌虫一生产1~2个卵块,单雌平均产卵量为107.3 ± 3.0 粒,平均孵化率为88.4 %±1.6%。

图6 朱红毛斑蛾的产卵行为Fig.6 Oviposition behaviors of Phauda flammans
2.4.2 朱红毛斑蛾的产卵节律 在5:00—23:00 均可观察到朱红毛斑蛾雌虫产卵,但主要集中在10:00—14:00(图7),且其产卵率显著高于其他时间段(F= 32.73,df= 24,725,P<0.001)。
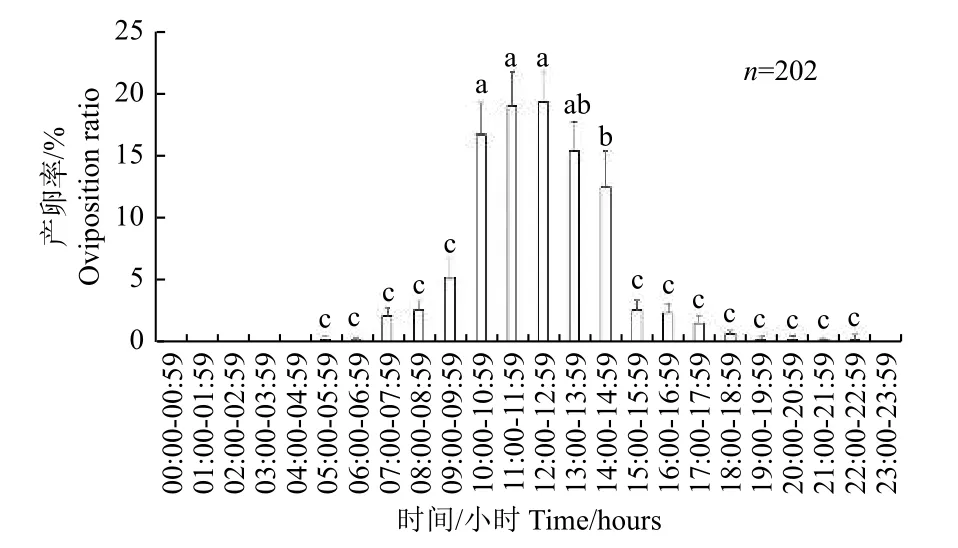
图7 朱红毛斑蛾产卵的昼夜节律Fig.7 Circadian rhythm of oviposition of Phauda flammans
3 讨论
一般情况下,昆虫经过长期的进化,其繁殖行为及节律相对稳定,主要是受其生物钟的调节[28]。本研究结果表明,朱红毛斑蛾的羽化、求偶和交配行为均发生在光期,为典型的日行性蛾类,与已报道的斑蛾科其他昆虫的繁殖行为和节律相似,如云南锦斑蛾(Achelurayunnanensis)[29]、朱颈褐锦斑蛾(Soritialeptalina)[30]、大叶黄杨长毛斑蛾(Pryeriasinica)[31],说明毛斑蛾科昆虫经过长期进化也表现出与斑蛾科类似的在光期繁殖的生物学特性。前人研究结果表明,昆虫繁殖行为及节律的变化可以减少物种对同一资源的直接竞争,并通过种群行为的同步,确保相似种间的遗传隔离[32]。事实上,蛾类的繁殖行为及节律除了由内在的生理因素决定之外,还受环境温度、光周期、湿度等生态因子的影响。然而,控制朱红毛斑蛾繁殖行为及节律的内外因子目前均尚不明确,需进一步研究。
朱红毛斑蛾羽化后,雌虫多停留在隐蔽处等待雄虫觅偶,觅偶成功后均是雄虫主动发起求偶,这与多数蛾类昆虫的求偶行为相似,如柚木野螟(Eutectona machaeralis)、栎黄枯叶蛾(Trabala vishnou gigantina)、草原毛虫(Gynaephora alpherakii)等[33-35]。前期研究证实,朱红毛斑蛾雌虫释放的性信息素对雄虫具有显著的引诱作用[8],推测朱红毛斑蛾雄虫在觅偶过程中依靠雌虫释放性信息素进行异性定位,但雄虫对性信息素的感受机制需进一步研究。
在蛾类昆虫中,多数种类均有多次交配的现象,因为多次交配对其有物质利益和基因利益等[36]。然而,朱红毛斑蛾雌雄成虫一生仅交配1次,为单配制蛾类,说明朱红毛斑蛾进行一次交配就能提供足够的精子供雌虫使用。在部分蛾类昆虫中,也观察到这种现象,如马尾松毛虫(Dendrolimus punctatus)、小菜蛾(Plutella xylostella)、粉斑螟(Cadra cautella)和棉褐环野螟(Haritalodes derogata)[37-40]。朱红毛斑蛾虽然一生仅交配1 次,但雌蛾的累计产卵量和孵化率均较高,说明该虫繁殖系数高,群体积累快,可能是其常暴发的潜在原因之一。
“选择性和适合度”假说(preference-performance hypothesis,PPH)认为由于植食性昆虫幼虫期不能远距离移动,所以雌虫常将卵产于适合其子代发育的寄主上[41]。朱红毛斑蛾是寡食性害虫,雌虫将卵产于小叶榕和垂叶榕叶柄或叶尖,推测是朱红毛斑蛾雌虫为便于其子代孵化后能有适口和充足的食物有关[42]。研究表明,卵的成功发育需要一个合适的产卵场所,其非生物条件适合,捕食率、寄生率和疾病率较低,及给子代留有充足的食物[43]。朱红毛斑蛾雌虫将卵产于寄主植物叶柄或叶尖,除了为子代提供充足的食物之外,是否还与抵御天敌的捕食或寄生有关尚不明确。此外,植食性昆虫产卵场所的选择是昆虫与寄主植物长期协同进化的结果[44],但朱红毛斑蛾产卵场所选择的进化驱动力仍有待进一步研究。
本研究结果还提示,可基于朱红毛斑蛾的繁殖行为及节律,在野外利用性信息素干扰其交配,使其错过最适交配时间,降低交配率,从而达到控制其种群数量的目的[5];在其繁殖高峰期,应用性信息素大量诱捕和监测该虫野外的种群动态及密度,为其防控、预测预报和制定综合防治策略等提供参考依据。
4 结论
朱红毛斑蛾仅在光期求偶、交配和产卵,并形成了相对固定的行为和节律,是一种日行性蛾类;一生仅交配1 次,为单配制蛾类。
