链形藻类的中国新记录属
——中眼藻属
2021-12-24董晓琪刘国祥
董晓琪 刘国祥 朱 欢
(1. 西藏大学理学院, 拉萨 850000; 2. 中国科学院藻类生物学重点实验室, 武汉 430072; 3. 中国科学院大学, 北京 100049)
绿色植物界(Viridiplantae)是由绿藻门(Chlorophyta)和链形植物门(Streptophyta)组成的一类光合生物的统称[1-6]。它主要由传统意义上的低等绿藻(Green algae)和高等植物(Higher plants)组成, 后者通常又被称之为有胚植物(Embryophyta)或陆生植物(Land plants)。根据Algaebase(https://www.algaebase.org/)和NCBI taxonomy(https://www.ncbi.nlm.nih.gov/taxonomy/)两个数据库所采用的分类系统, 这一定义中的绿藻(Green algae)分为两大类群, 其一为传统的绿藻门类群(即绿藻门, Chlorophyta), 它主要包括葱绿藻(Prasinophytes)和核心绿藻(core Chlorophytes)两个大类群; 其二为链形藻类(Streptophyta algae), 它主要包括广义的轮藻门藻类(Charophytasensu lato), 即由轮藻纲(Charophyceae)、双星藻纲(Zygnematophyceae)、鞘毛藻纲(Coleochaetophyceae)、克里藻纲(Klebsormidiophyceae)、绿方藻纲(Chlorokybophyceae)和中眼藻纲(Mesostigmatophyceae)六个类群的藻类组成。
被藻类学家广为采纳的绿藻分类系统逐渐脱离了依据形态特征所建立的框架, 并与基于鞭毛根系统(Flagellate root system)的超微结构研究和分子系统发育研究所建立的分类框架趋于一致[5,7,8]。而在这一系列的研究进程中, 葱绿藻的分类与系统发育具有极为重要的意义。葱绿藻的典型特征是原生质体外多由酸性多糖质的鳞片所包被, 且在光合作用产物、游动细胞形态上更为原始和多样。越来越多的研究表明, 这一分类框架中的葱绿藻为深度发生且高度并系的类群。葱绿藻也被认为是绿色植物中最古老的类群[9-11]。
中眼藻属最早被描述于1894年[12], 是一种鞭毛器(Flagellate apparatus)具有多层结构(Multi-layers structure, MLS)且有鳞包被的绿色藻类, 能产生并维持一个复杂的三层胞外基质, 由内部菱形鳞片、中部椭圆形鳞片和外部篮状鳞片三种类型的鳞片组成[13,14]。在葱绿藻中, 中眼藻属被描述为一个特殊的类群, 其多糖质的鳞片与特殊的鞭毛位置使得藻类学家将其与其他葱绿藻并列归入绿藻门中。而基于鞭毛根系统的超微结构研究支持中眼藻与轮藻和高等植物具有更近的亲缘关系, 分子系统发育的研究则进一步肯定了基于超微结构的研究结果[2]。近20年来绝大多数关于中眼藻的超微结构和分子生物学研究证实其为链形植物的一个分支, 而不是处于绿藻门支系中[15—17]。基于细胞器基因组数据和全基因组数据的系统发育基因组学研究更进一步支持中眼藻是链形植物门中最为原始的类群[18—20]。现在藻类学界普遍认为已经发现的单细胞鞭毛藻中, 中眼藻是与高等植物亲缘关系最近的藻类[21]。
中眼藻主要分布于静止的淡水水体中, 诸如池塘、水库和小型湖泊等[22]。本研究对采自重庆合川和福建九龙江水库的标本进行了形态观察和分子系统发育分析, 描述了中眼藻的形态特征和系统发育位置; 同时以三峡水库为例, 应用环境DNA技术检测了中眼藻在水库中的时空分布情况。这是中眼藻属在中国的首次报道。
1 材料与方法
1.1 材料采集与形态观察
采集中眼藻样品使用25号浮游生物网, 采集后的标本一部分用福尔马林加10%的甘油固定用于形态观察, 另一部分用无水乙醇固定用于DNA提取。样品采集信息见表 1。环境DNA采集与样品处理方法参见Hu等[23], 采集信息见表 2。形态观察在光学显微镜(DM5000B: Leica, Wetzlar, Germany)下进行, 由于中眼藻细胞较小, 使用油镜(100×)对其进行观察; 利用Leica DFC320数码相机对藻体形态特征、藻体颜色、长宽比例和藻体内部结构等具有辨识度的特征进行拍摄。

表1 中眼藻标本信息及GenBank登录号Tab. 1 Sample time of the studied specimens and GenBank accession number
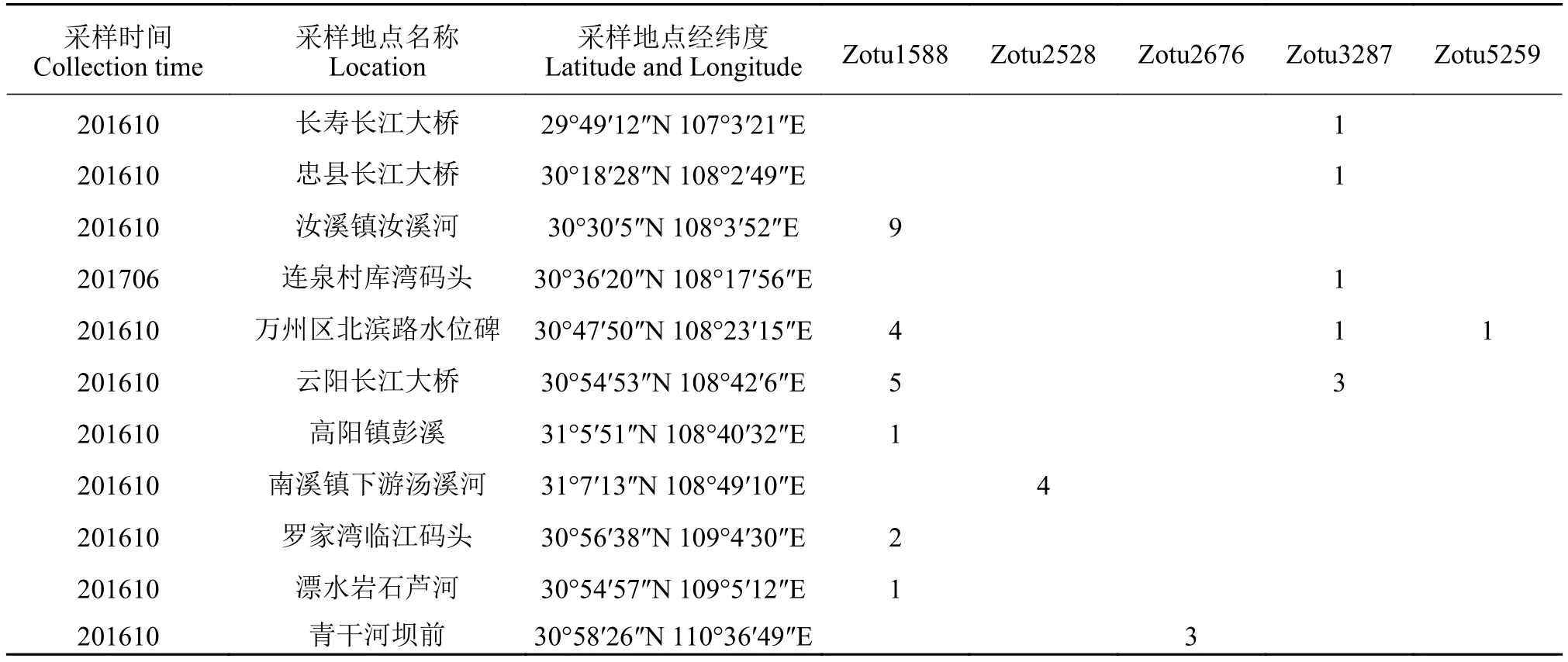
表2 环境DNA采集信息详情Tab. 2 Details of environmental DNA collection
1.2 单细胞PCR扩增及测序
模板DNA制备: 在倒置显微镜下确定目标藻体, 用孔径约0.1 mm的微吸管吸取单细胞直到提取出的细胞仅为中眼藻1种, 用移液枪吸取50 μL含有细胞的缓冲液(PBS)至PCR管中, 加入2 μL裂解缓冲液(Vazyme, Nanjing, China), 95 ℃热处理15min, 处理后将获取的DNA模板保存于-20 ℃冰箱中备用。
PCR扩增体系共50 μL, 其中包含正反向引物各1.5 μL, 引物最终浓度为0.4 μmol/L; 4 μL模板DNA; 25 μL2×TaqMaster Mix(ExTaq, TaKaRa,Dalian, China); 无菌双蒸水18 μL。18S rDNA扩增引物及反应条件参照Honda等[24]。
PCR 扩增产物经1.0%的琼脂糖凝胶电泳, 根据条带长度切下目标条带, 凝胶回收试剂盒回收(E.Z.N.A. Gel Extraction Kit Omega Bio-Tek, America)并纯化PCR产物后送至擎科生物公司测序(TSING KE, Wuhan, China), 测序后的序列上传至GenBank (http://www.ncbi.nlm.nih.gov/genbank/)。环境DNA的提取及测序方法参见Hu等[23]。环境DNA序列上传至NCBI数据库, 其Bioproject Accession为PRJNA496371。
1.3 数据分析
使用NCBI核酸数据库对18S rDNA基因序列进行BLAST相似性对比, 下载相似度高的序列片段,首先使用MAFFT 7.4对序列矩阵进行比对[25], 然后使用MEGA 7.0软件对序列进行手动校正[26], 在DAMBE5.0中对数据质量进行评估[27]。使用ModelFinder软件选择最适模型[28]。采用最大似然法(Maximum likelihood, ML)和贝叶斯法构建系统进化树, 最大似然法使用软件为 RAxML 8.0[29], 贝叶斯法使用 MrBayes 3.2[30]。
2 结果
2.1 形态观察
中眼藻属
Mesostigma Lauterborn 1894
Lauterborn, R. Ueber die Winterfauna einiger Gewässer der Oberrheinebene. Mit Beschreibungen neuer Protozoen. Biologische Centralblatt, 1894, 14:390—398
Holotype species:Mesostigma virideLauterborn 1894
细胞扁平到鞍形, 从背侧凹陷处伸出2条鞭毛。腹面略凹。鞭毛基部位置靠近细胞前端, 鞭毛或多或少地指向运动细胞的前部, 该运动细胞在运动过程中会快速旋转。叶绿体单个, 覆盖腹侧和外侧, 并延伸到背侧。腹侧具单个眼点, 靠近细胞的中轴。鞭毛基部附近有几个伸缩泡。在模式种中存在2个蛋白核, 而其他物种则缺乏蛋白核。细胞和鞭毛覆盖着多种类型的鳞片。原生质体最外层的鳞片足够大, 以至于在光学显微镜下可见。繁殖方式未知。
该属为中国新记录属。根据Algaebase在线数据库的记录有Mesostigma virideLauterborn和Mesostigma grandeKorshikov两种, 区分这两种的关键特征在于蛋白核数量不同。亦有藻类学家认为中眼藻属为单种属,M. grande为M. viride的异名。
绿色中眼藻(图 1a—i)
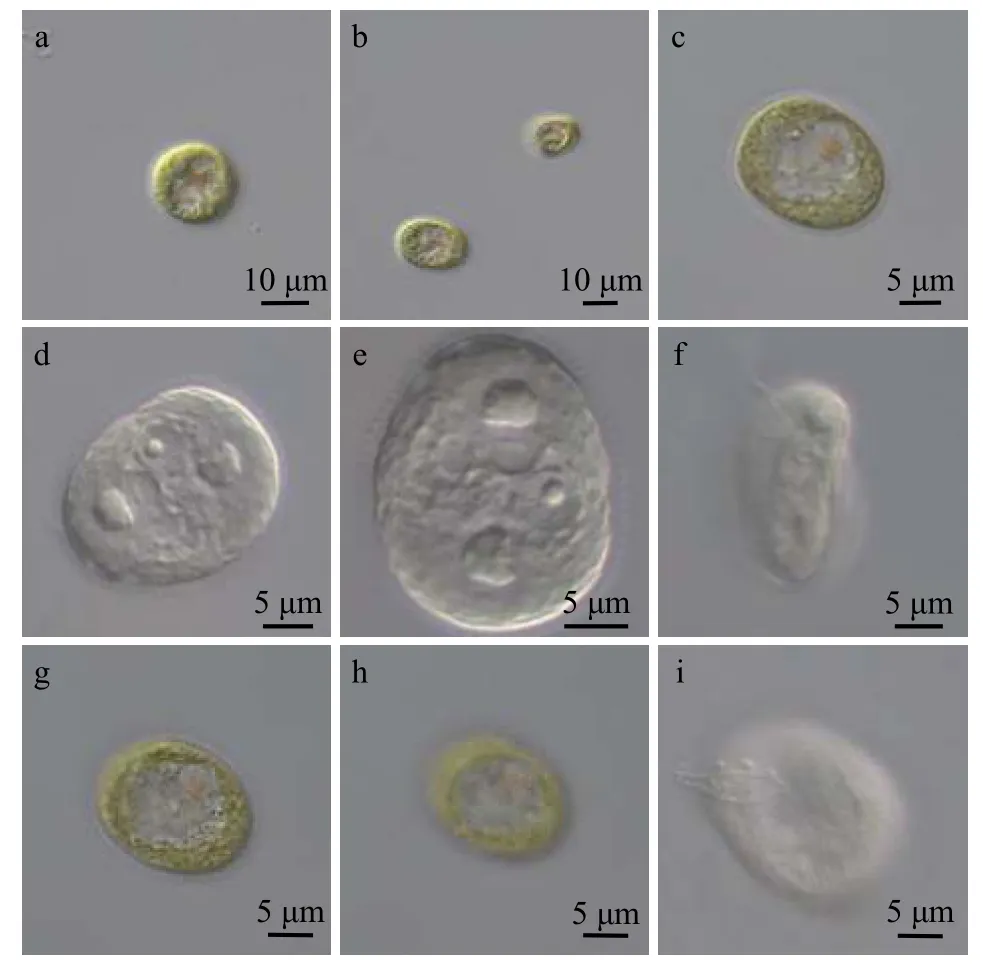
图1 本研究中采集的绿色中眼藻的形态特征Fig. 1 The morphological characteristics of Mesostigma viride cells
Mesostigma viride Lauterborn 1894
Lauterborn, R. Ueber die Winterfauna einiger Gewässer der Oberrheinebene. Mit Beschreibungen neuer Protozoen. Biologische Centralblatt, 1894, 14:390—398
细胞扁平, 椭圆形或圆形, 稍不对称, 呈黄绿色,细胞长14—20 μm, 宽11—16 μm(图 1a、1b和1c);原生质体外无细胞壁, 而是由多糖质鳞片包被(图 1e),且鳞片呈网状(图 1h); 细胞内具1个大液泡(图 1c),2—4个伸缩泡(图 1g)及2个蛋白核(图 1d); 叶绿体1个, 暗绿色, 片状, 周生; 细胞腹侧中轴处具1个眼点(图 1a); 背侧凹陷处具两根鞭毛(图 1i), 呈螺旋形前进(可见其侧面观图 1f)。以上特征与中眼藻(Mesostigma viride)形态特征符合, 故可判断该藻为中眼藻。
该种为中国新记录种。
2.2 分子系统发育分析
两株中眼藻的18S rDNA序列长度分别为1649 bp(MW413341)和1605 bp(MW413342)。从环境样本中获得的18S rDNA V4区的5条序列长度分别为381 bp(Zotu1588)、382 bp(Zotu2528)、381 bp(Zotu 2676)、380 bp(Zotu3287)和380 bp(Zotu5259)。NCBI核酸数据库对基因序列进行相似性对比显示以上7条序列与GenBank中已有的中眼藻序列相似度为100%。用于构建系统发育树的序列矩阵信息见表 3。

表3 构建系统发育树数据详情Tab. 3 Details of the data used to build phylogenetic trees
基于中眼藻18S rDNA序列构建的系统进化树(图 2), 运用最大似然法和贝叶斯法得到的拓扑结构基本一致。本实验所采集到的2株藻与Gen-Bank中已有的中眼藻KJ808698Mesostigma viride聚在一起, 其自展支持值和后验概率分别为100和1.00, 支持本实验所采集藻类为绿色中眼藻。从系统进化树可以明显地看出, 中眼藻纲(Mesostigmatophyceae)与绿方藻纲 (Chlorokybophyceae)以较高的自展支持值和后验概率聚为一枝。环境DNA系统发育树结果显示(图 3), 5个不同的环境序列均以较高的自展支持值与绿色中眼藻聚在一起。另外, 通过对三峡水库4个季节的环境DNA调查, 我们发现中眼藻在秋季出现的频率最高,夏季仅在1个样品中检测到, 而冬春季节均未发现该藻。
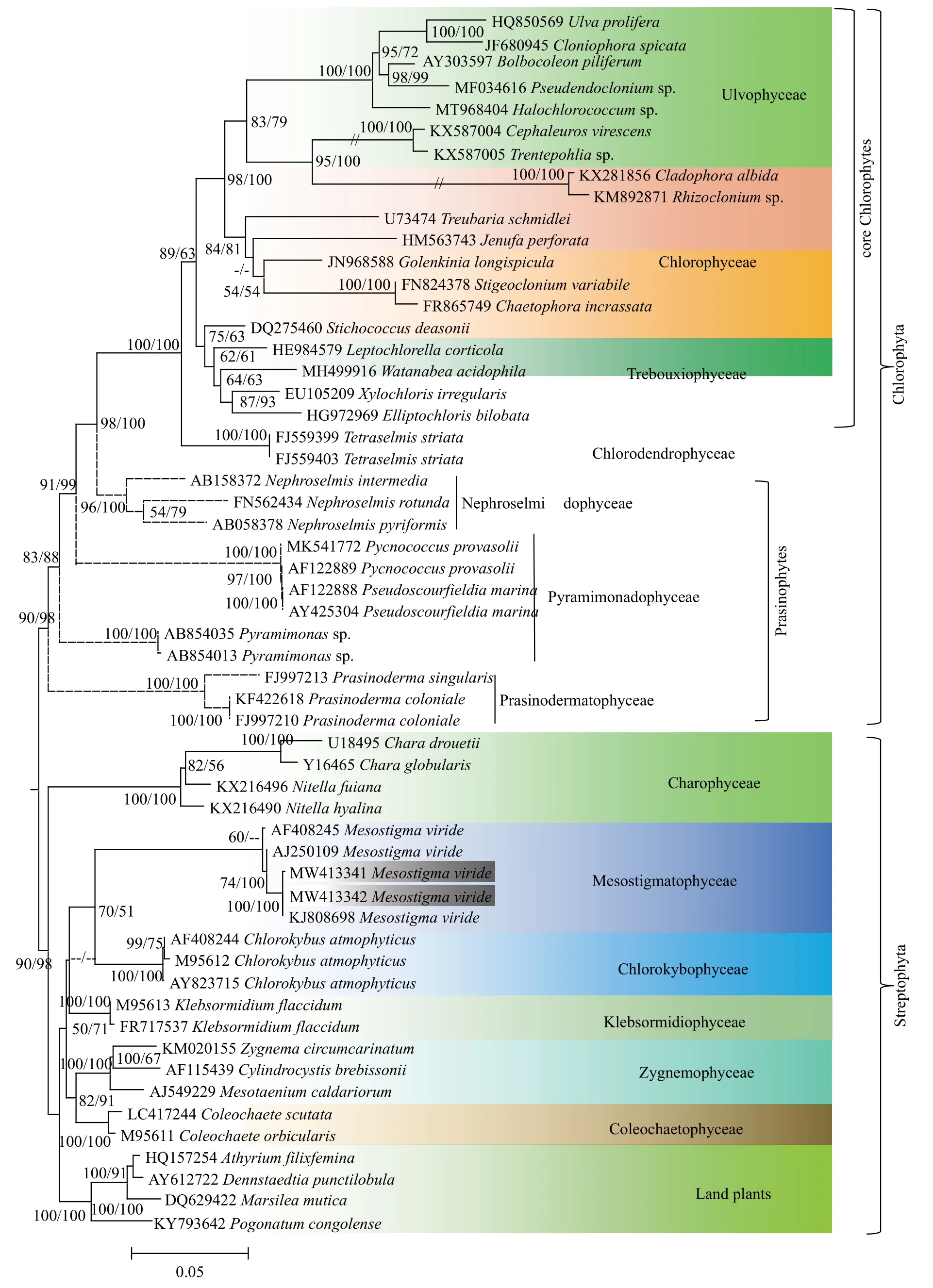
图2 基于中眼藻18S rDNA序列构建的系统进化树Fig. 2 The phylogram inferred from18S rDNA sequences of Mesostigma viride
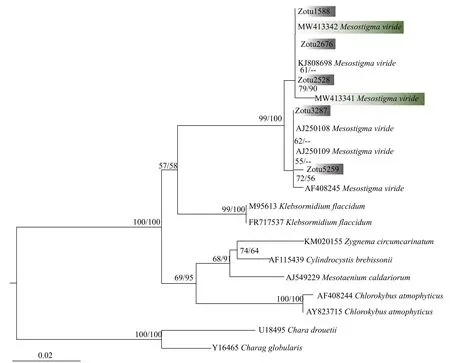
图3 基于环境中18S rDNA构建的系统发育树Fig. 3 The phylogram inferred from 18S rDNA sequences of environmental DNA
3 讨论
不同于鞭毛位置位于细胞前端的绿藻, 中眼藻鞭毛位于背部凹陷处, 这使得其运动方式表现为特殊的螺旋式前进, 可作为区分中眼藻与其他常见单细胞鞭毛绿藻的重要特征。由于原生质体外不具由纤维素和果胶组成的细胞壁, 中眼藻最初被认为隶属于葱绿藻纲[31], 但分子系统发育研究表明这一类群并不是自然发生的。本研究结果表明, 中眼藻与链形植物门中的轮藻纲和绿方藻纲具有更近的亲缘关系, 进一步支持中眼藻在系统发育位置上处于链形植物门的基部, 这与Bhattacharya等[18]对中眼藻肌动蛋白的研究和Nedelcu 等[16]对中眼藻表达序列标签(Expressed Sequence Tags, EST)的研究结果一致。
依据环境样本中获取的18S rDNA V4区片段,我们发现三峡水库多个区域存在中眼藻, 且主要出现在秋季。然而我们通过显微观察却并未在这些水体中发现中眼藻, 这表明中眼藻可能比我们预期中要更难发现, 同时也说明秋季可能是在中国内陆采集中眼藻的最佳季节。不同18S rDNA V4区序列在系统发育树上的位置略有区别, Zotu 3287和Zotu5259以72的自展支持值与AF408245Mesostigma viride聚为一枝; Zotu1588、Zotu2676和Zotu2528以79的自展支持值及90%的贝叶斯后验概率与本研究中采集到的中眼藻聚为一枝; 而以上两大分枝以99的自展支持值及100%的后验概率分为两枝, 这暗示了中眼藻属中有可能存在隐存种。在以后的研究工作中, 应用单细胞PCR手段获取多个分子标记进行详细比较, 同时获取更为精细的显微特征数据, 对于解决这一问题可能较为有效。
目前, 对于中眼藻系统发育的研究结果表明中眼藻在链形植物类群中单独形成一个深度发生的枝系, 与陆生植物有着密不可分的联系, 是研究绿色植物从水生绿藻向陆生植物过渡的理想材料。本研究首次在中国发现并记录了中眼藻属,对中眼藻在中国内陆水体的分布和出现时间进行了初步推测, 为在中国开展中眼藻相关研究奠定基础。
