黏虫保幼激素环氧水解酶基因MsJHEH2的克隆与生物学功能
2021-12-17杨洪佳张春雨胡育硕
李 泽,杨洪佳,张 良,张春雨,胡育硕,樊 东
(东北农业大学农学院,哈尔滨 150030)
在昆虫的生长发育中,昆虫的蜕皮与变态受到多种激素的影响,其中最主要的是保幼激素(Juvenile hormone,JH)和蜕皮激素(molting hormone,MH)[1]。蜕皮激素诱导昆虫的蜕皮和变态,而保幼激素则阻止由蜕皮激素引起的变态,使幼虫蜕皮后仍然维持幼虫形态[2,3]。在昆虫的龄期末期通常观察到JH滴度的急剧下降和蜕皮激素滴度的急剧上升,由于JH与MH在功能上的拮抗作用和在昆虫生长发育过程中其滴度变化呈现相反趋势,暗示了MH对JH的降解可能具有促进作用[4]。昆虫蜕皮过程发生在很短的时间内,因此JH的快速降解对昆虫的发展具有重要意义。
保幼激素降解过程中主要有三种酶参与:保幼激素酯酶(Juvenile hormone esterase,JHE)、保幼激素环氧水解酶(juvenile hormone epoxide hydrolase,JHEH)和保幼激素二醇激酶(Juvenile hormone diol kinase,JHDK)[5-7]。其中,JHEH负责不可逆地打开环氧环,将保幼激素水解为保幼激素二醇,将保幼激素酸水解为保幼激素酸二醇[8-10]。由于JH在昆虫发育和生殖过程中发挥着重要的调控作用[11],长期以来,干扰其生物合成和代谢一直被认为是一种很有前途的替代化学杀虫剂的方法,并且对非目标生物具有低毒性[12]。大多数关于JH降解的研究都集中在JHE的作用机制上,但越来越多的研究表明,JHEH在昆虫发育中和JHE一样重要[13],因为保幼激素二醇和保幼激素酸二醇是昆虫的主要代谢物[14]。因此,由于其在JH调控和不可逆降解中的作用,对于JHEH的研究也是至关重要的[15]。
以往对于昆虫保幼激素环氧水解酶的研究多集中在鳞翅目,自从从烟草天蛾Manduca sexta[16]卵中克隆出第一个JHEH基因后,在粉纹夜蛾Trichoplusia ni[17]、家蚕Bombyx mori[18]、棉铃虫Helicoverpa armigera[19]、小菜蛾Plutella xylostella[20]、斜纹夜蛾Spodoptera litura[21]、赤拟谷盗Tribolium castaneum[14]等昆虫中对JHEH进行了深入研究,发现大多数JHEH均具有水解JH的功能,且在一些昆虫如赤拟谷盗中,不止一种JHEH[14],而是多种JHEH共同参与调控昆虫的蜕皮、繁殖、滞育和变态等一系列重要生物学功能,对昆虫正常的生长发育具有重要意义。
黏虫Mythimna separata属于鳞翅目Lepidoptera夜蛾科Noctuidae,是我国重要的农业害虫,一旦大暴发会对我国粮食作物造成十分严重的危害[22]。目前对于黏虫的防治多采用化学防治,但逐年凸显的抗药性问题让我们不得不寻找其他更为有效的防治方法[23]。本研究以黏虫为研究对象,探究MsJHEH2基因在黏虫体内的表达情况,并利用RNAi技术将MsJHEH2基因沉默,探究基因沉默后黏虫体内JH含量的变化,以及对黏虫生长发育的影响,为今后提出通过保幼激素降解途径防治黏虫的生物技术防治方法提供理论基础。
1 材料与方法
1.1 供试昆虫
试验所用黏虫采自东北农业大学向阳试验示范基地,在室内连续培养数代。将黏虫幼虫放入人工气候箱中饲养,温度为(25±1)℃、相对湿度为(65±5)%、光周期为14L:10D,期间喂食新鲜玉米叶片。将预蛹期幼虫放入装有湿润土壤的培养盒中,待其羽化后,放入养虫笼中,喂食蜂蜜水,在笼中放入用于产卵的塑料条,每2 d更换一次。待其产卵后,每天更换塑料条并将带有卵块的塑料条放到通风湿润的培养盒内,待其孵化后继续饲养。
1.2 黏虫JHEH基因的克隆及序列分析
1.2.1 黏虫转录组测序 收集黏虫卵、1~6龄各龄期的幼虫、预蛹、蛹和成虫,用液氮速冻后将样本送到安诺优达基因科技有限公司进行转录组测序。
1.2.2 cDNA 序列的克隆与鉴定 在获得的转录组数据库中进行保幼激素环氧水解酶基因序列的筛选,并将筛选出的基因序列在NCBI(https://blast.ncbi.nlm.nih.gov/Blast.cgi)网站上使用BLAST进行进一步的同源性比对。使用Primer 5软件设计引物MsJHEH2-F和MsJHEH2-R(表1),通过PCR扩增、胶回收和测序,对在转录组数据库中筛选出的保幼激素环氧水解酶基因序列进行准确性校正。将获得的完整的保幼激素环氧水解酶基因cDNA序列在GenBank上登录。

表1 引物名称、序列及用途Table 1 Primer name, sequence and purpose
1.2.3 序列分析及系统进化树的构建 利用在线网站ExPASy(http://ca.expasy.org/tools/)预测氨基酸的等电点和分子量,利用在线网站SMART(http://smart.embl-heidelberg.de/)、HMMER(https://www.ebi.ac.uk/Tools/hmmer)和 InterProScan(http://www.ebi.ac.uk/interpro)预测氨基酸的结构域,利用在线网站ProtParam(https://www.expasy.org/resources/protparam)对氨基酸的组成及理化性质进行分析,亚细胞定位使用网站CELLO(http://cello.life.nctu.edu.tw/)进行预测,蛋白质跨膜结构预测采用TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)网站进行。在NCBI网站使用BLAST搜索昆虫JHEH序列,利用MEGA7.0软件中邻位相连(Neighbor-joining)法构建系统进化树。
1.3 MsJHEH2基因表达模式研究
1.3.1 不同龄期表达模式分析 分别选取生长情况良好的黏虫卵、1~6龄幼虫、预蛹、蛹和成虫的第1 d虫体,放入液氮中速冻后置于-80 ℃冰箱中备用。利用TRIzol法提取黏虫总RNA,每100 mg组织中加入1 mL Invitrogen公司生产的TRIzol®Reagent,提取得到的总RNA用紫外分光光度计鉴定其纯度和浓度,检测合格的RNA放入-80 ℃冰箱备用。取2 µg总RNA,使用TOYOBO公司生产的反转录试剂盒(Revertra Tra Ace qPCR RT Master Mix with gDNA Remover)对提取的RNA进行反转录,合成得到cDNA第1链,放入-20 ℃冰箱保存备用。利用获得的MsJHEH2cDNA序列设计荧光定量引物MsJHEH2-q-F和MsJHEH2-q-R,使用两个内参基因G-3-P和β-actin,荧光定量引物G-3-P-F、G-3-P-R、β-actin-F、β-actin-R见表1,黏虫不同发育阶段的cDNA作为模板进行qRT-PCR,使用荧光染料SYBR Primer Script RT−PCR Kit Mix(TOYOBO,日本)在Bio−Rad荧光定量PCR仪上通过三步法进行。反应体系为20 μL:SYBR qPCR Mix 10 μL,上、下游引物各 0.6 μL,底物 cDNA 2 μL,ddH2O 6.8 μL。反应程序为:94 ℃ 1 min,95 ℃ 30 s,59 ℃ 30 s,72 ℃ 30 s,共进行40个循环。每个样品重复3次,生物学重复3次。
1.3.2 3龄幼虫不同发育时间表达模式分析 为了进一步明确MsJHEH2在一个龄期的不同发育时间的表达模式,对即将蜕皮的2龄幼虫进行观察,选取生长情况良好并一致的3龄3、6、12、24、48 h幼虫各3头,放入液氮中进行速冻,保存于-80 ℃中。对各样品分别进行总RNA提取,反转录得到cDNA第1链,通过qRT-PCR检测MsJHEH2基因表达量,操作方法同1.3.1。
1.3.3 不同组织内表达模式分析 选取生长状况良好的4龄第1 d黏虫幼虫,将其在RNA组织保存液中进行解剖,获得前肠、中肠、后肠、脂肪体、马氏管、唾腺和体壁7个组织。分别提取各组织RNA,反转录得到cDNA第1链,并通过荧光定量PCR检测基因表达量,操作方法参照1.3.1。设3次生物学重复。
1997年,朗格推出先进而精密的自动上链机心——屡获殊荣的Sax-0-Mat型机心,内置归零装置,于表冠拉起时掣停摆轮,秒针即时返回零位,就如计时码表一样。Sax-0-Mat型机心仍为Langematik Perpetual和Saxonia ANNUAL CALENDAR等表款的基本机心。1815 Tourbillon和Richard Lange Jumping Seconds的手动上链机心亦设有归零功能,简化了时间设定过程。往后的表款中,归零装置采用独立模组设计,附设由多个圆盘组成的离合器,使秒针在突然的震荡或反弹中仍平稳运作。
1.4 MsJHEH2基因对不同浓度保幼激素的表达响应
选用生长状况良好的4龄第一天黏虫幼虫,使用注射法对黏虫进行保幼激素类似物(Juvenile hormone analogue,JHA)处理,保幼激素类似物烯虫酯(S-(+)-Methoprene)[24]购自aladdin公司,用1%的DMSO(购自coolaber公司)稀释成五个浓度梯度,0.3125、0.625、1.25、2.5和5 µg/µL,1%的DMSO作为对照,在每头幼虫腹部2~3节注射2 µL不同浓度保幼激素类似物。将注射后的幼虫进行单头饲养,并在处理后3、6、12、24和48 h分别收集存活的幼虫各3头,放入液氮中速冻致死后放于-80 ℃冰箱保存备用。对各样品分别进行总RNA提取,反转录得到cDNA第1链,并通过荧光定量PCR检测基因表达量,操作方法同1.3.1。
1.5 MsJHEH2基因生物功能研究
1.5.1 RNA干扰处理与MsJHEH2基因表达量检测 将MsJHEH2基因完整的cDNA序列发送至上海吉玛公司分别设计并合成MsJHEH2基因的siRNA和与靶基因无同源性且在黏虫体内没有生物学效应的阴性对照(Negative Control,NC),引物序列siMsJHEH2-F、siMsJHEH2-R、Negative control-F、Negative control-R见表1,用对应体积ddH2O分别稀释成浓度为20 µmol/L备用。选取生长状况良好的4龄第1 d黏虫幼虫,对其进行siRNA注射,操作方法同1.4。将注射后的幼虫进行单头饲养,并在处理后的3、6、12、24、48、72和96 h后分别收集存活的幼虫各3头,放入液氮中速冻致死后放于-80 ℃冰箱保存备用。将处理后的黏虫幼虫进行总RNA提取,反转录得到cDNA第1链,并通过qRT-PCR检测基因表达量,操作方法如1.3.1,检测干扰不同时间后基因沉默效率。
1.5.2 RNA干扰后保幼激素含量检测 首先参照考马斯亮蓝G-250染色法测定RNA干扰不同时间后黏虫体内总蛋白质含量[25],利用昆虫保幼激素(JH)酶联免疫分析(ELISA)试剂盒(江苏酶标生物科技有限公司)测定RNA干扰后不同时间黏虫体内保幼激素(JH)含量,具体操作见说明书。样品折算保幼激素含量=样品实际保幼激素含量/样品总蛋白质含量,以样品折算保幼激素含量作为结果中样品的最终保幼激素含量。
1.5.3MsJHEH2基因对黏虫生长发育的调控作用研究 选取体质量基本一致,生长状况良好的4龄第1 d黏虫幼虫,用万分之一天平分别称重后注射 siRNA,操作方法同 1.4,注射后将每头黏虫幼虫单独放于塑料平皿中,放入恒温培养箱中继续饲养,每个处理注射30头黏虫幼虫,设3次生物学重复。注射后的黏虫幼虫每24 h观察1次并记录体重变化,并用万分之一天平称量单头蛹(化蛹第二日)的重量。观察并记录幼虫4、5、6龄的发育历期,统计幼虫化蛹率、蛹历期及羽化率。
1.6 数据统计与分析
采用2-ΔΔCt法计算MsJHEH2基因相对表达量[26]。利用IMB SPSS Statistics 23软件单因素方差分析的方法进行黏虫体内各组织、各龄期及JHA处理后不同时间点MsJHEH2基因相对表达量的差异显著性分析,利用独立样本T检验的方法进行RNA干扰后MsJHEH2基因表达量、保幼激素含量、化蛹率和羽化率各数据间差异显著性分析,显著性水平为P<0.05。并在Excel 2010中计算数值及制作图表。
2 结果与分析
2.1 MsJHEH2基因的克隆与分析
本试验通过对黏虫转录组数据库搜索、NCBI比对、保守结构域查找以及序列验证,得到1条包含完整开放阅读框(ORF)序列的保幼激素环氧水解酶基因cDNA序列。序列验证后,在NCBI上进行登录并命名为MsJHEH2(登录号:MT802192)。利用在线软件ProtParam对MsJHEH2基因编码氨基酸的理化性质进行了分析,结果表明,MsJHEH2基因cDNA长度为1533 bp,ORF 1389 bp,编码462个氨基酸,第 51~160个氨基酸之间存在EHN环氧水解酶超家族结构域(图1)。该蛋白质共有 7443个原子,分子式为 C2455H3726N598O653S11,分子量为 52.42 kD;该蛋白质的负电荷残基(Asp+Glu)数为50个,正电荷残基(Arg+Lys)为50个,理论等电点为7.07;该蛋白质的不稳定指数为30.85,据此可将其归为稳定蛋白;脂肪族指数为90.32,平均疏水性(GRAVY)为−0.023;蛋白中含量最多的氨基酸为Ala(6.5%)。利用InterProScan和Hmmer工具进行的氨基酸分析得到3个环氧水解酶家族标签,即序列催化三联体Asp226、Glu402、His429残基。此外发现MsJHEH蛋白保守基序HGWP151花样结构,2个作为稳定剂结合环氧环中氧离子洞结构的Tyr297、Tyr372残基,N-端的跨膜基序XWG37(其中X为芳香族残基)。亚细胞定位预测结果显示,MsJHEH2基因编码蛋白位于质膜(PlasmaMembrane)和细胞质(Cytoplasmic)的可能性最大。TMHMM网站(http://www.cbs.dtu.dk/services/TMHMM/)预测结果显示,MsJHEH2编码蛋白存在一个跨膜结构,位于第 5~23个氨基酸之间。

图1 MsJHEH2基因 cDNA 序列及推导的氨基酸序列Fig. 1 cDNA and deduced amino acid sequences of MsJHEH2
2.2 MsJHEH2基因系统进化树构建
利用系统进化树分析MsJHEH2与其他昆虫JHEH基因的亲缘关系(图2)。分析发现,黏虫M. separata(QOI60666)首先与夜蛾科昆虫,如草地贪夜蛾Spodoptera frugiperda(XP_035454913)、斜纹夜蛾S. litura(QDP42285)、棉铃虫H. armigera(XP_021199401)和粉纹夜蛾T. ni(XP_026738426)聚类,显示它们的亲缘关系最近;再与蛾类其他昆虫如亚洲玉米螟Ostrinia furnacalis(XP_028170523)、烟草天蛾M. sexta(XP_037300979)、草地夜蛾Bombyx mandarina(XP_028039332)、家蚕B, mori(NP_001037201)聚类,显示它们的亲缘关系较近;最后与蝶类如金凤蝶Papilio machaon(XP_014361683)、柑橘凤蝶Papilio xuthus(KPI98602)、黑脉金斑蝶Danaus plexippus plexippus(XP_032521996)等昆虫聚类,显示它们的亲缘关系相对较远。说明在昆虫中,JHEH在鳞翅目中的同源性具有高度保守性,昆虫保幼激素环氧水解酶之间的系统进化关系与形态特征的系统进化关系具有一定的相关性。

图2 基于氨基酸序列构建的黏虫与其他昆虫JHEH蛋白的系统进化树Fig. 2 Phylogenetic tree of JHEH proteins from Mythimna separata and other insects
2.3 黏虫MsJHEH2基因表达模式分析
2.3.1 在不同发育阶段表达模式分析 qRT-PCR分析结果显示MsJHEH2基因在黏虫不同发育阶段内均有表达,其中在蛹期表达量最高,是成虫体内表达量的40.5倍,在5龄和预蛹期内表达量相对较高,二者差异不显著但均与其他发育阶段差异显著。1龄幼虫中相对表达量也较高,其他发育阶段表达量均较低,没有显著性差异(图3)。
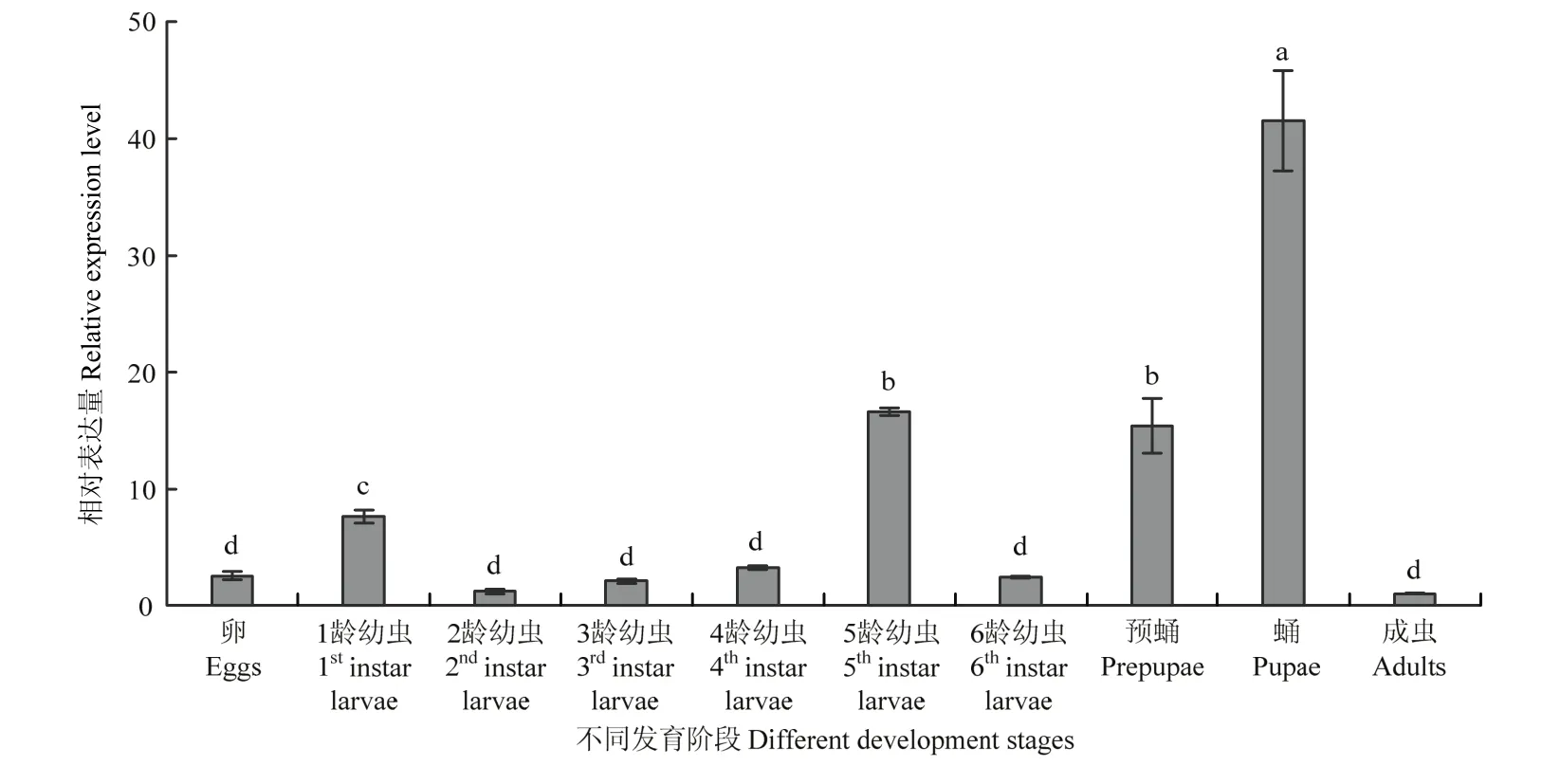
图3 黏虫不同发育阶段MsJHEH2表达模式Fig. 3 MsJHEH2 expression pattern at different developmental stages of M. separata
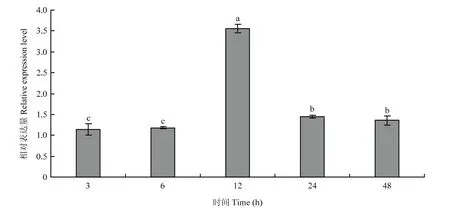
图4 黏虫3龄期内不同时间MsJHEH2表达模式Fig. 4 MsJHEH2 expression pattern at different time points in the 3rd instar larvae of M. separata
2.3.3在不同组织中表达模式分析 qRT-PCR分析结果显示,MsJHEH2基因在黏虫中肠和体壁中相对表达量最高,与其他组织中表达量相比差异显著,分别为唾腺中表达量的181.04和168.09倍;在脂肪体中基因表达量较高;在前肠、后肠和马氏管中表达量相对较低,无明显差异,其中唾腺内MsJHEH2基因表达量最少(图5)。
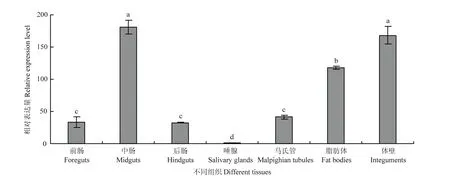
图5 黏虫不同组织MsJHEH2表达模式Fig. 5 MsJHEH2 expression pattern in different tissues of M. separata
2.4 不同浓度JHA处理对黏虫MsJHEH2基因表达的影响
JHA处理后不同时间点MsJHEH2基因表达均有显著性差异,不同浓度JHA处理后基因MsJHEH2表达量随时间变化成上升趋势,并在12 h达到最高,后缓慢下降。低浓度JHA(0.3125、0.625、1.25 µg/µL)处理12 h后均促进MsJHEH2基因表达,而高浓度JHA(2.5、5 µg/µL)处理后基因上调程度较低,并在48 h抑制其表达(图6)。
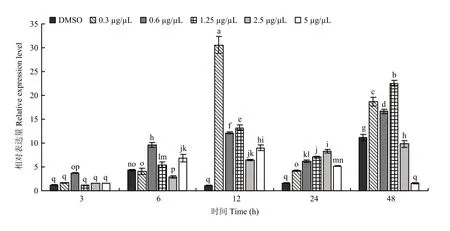
图6 JHA对黏虫MsJHEH2表达的影响Fig. 6 Effects of JHA on the expression of MsJHEH2 in M. separata
2.5 黏虫MsJHEH2基因生物学功能研究
2.5.1 RNA干扰对MsJHEH2基因表达量的影响 RNA干扰后,MsJHEH2基因被成功沉默。MsJHEH2基因表达量在处理后的3~24 h均有不同程度的下降,沉默效率分别为32.60%、64.86%、43.88%和15.36%,在6 h时干扰效果最好。在处理48 h后无干扰效果,72 h后,MsJHEH2基因表达量出现上调趋势,但与对照差异不显著(图7)。

图7 RNAi对黏虫MsJHEH2基因表达的影响Fig. 7 MsJHEH2 expression after treated with RNAi
2.5.2 RNA干扰对保幼激素含量的影响 RNA干扰后,随着MsJHEH2基因表达量的降低,不同时间内黏虫体内保幼激素含量与对照相比均有明显上调,分别为对照的1.51、1.58、1.52、1.39、1.36、1.41和1.38倍,且差异均显著(图8)。

图8 RNAi对黏虫体内保幼激素含量的影响Fig. 8 Effects of juvenile hormone content in M. separata treated with RNAi
2.5.3MsJHEH2基因生物功能研究 为探究MsJHEH2基因的生物学功能,对RNA干扰后黏虫生长发育情况进行了观察。黏虫的体重在基因沉默后随着时间的变化呈现出增长的趋势,其中第5、6 d,体质量增长最为明显,均有显著性差异(图9)。

图9 RNAi对黏虫体质量的影响Fig. 9 Effects of the M. separata weight treated with RNAi
RNA干扰后,随着黏虫体质量增长的同时,黏虫幼虫的发育进程虽然没有显著性变化,但化蛹率、羽化率均降低,羽化所需时间延长,前两天羽化率差异显著,4 d后羽化率未发生变化(表2,3)。
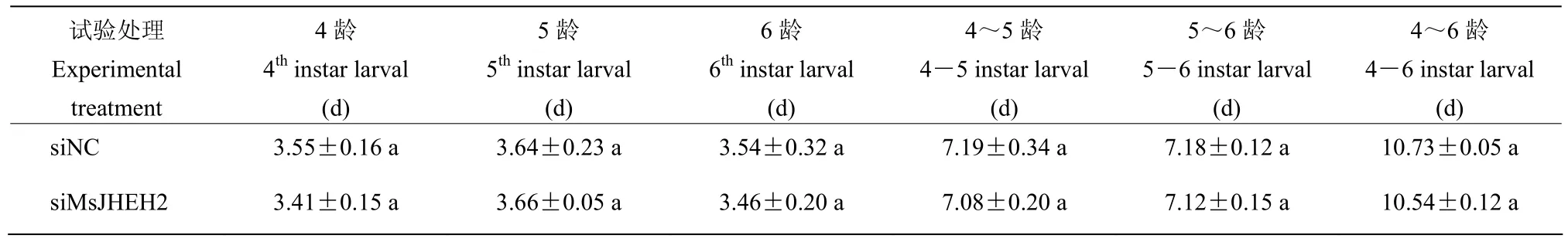
表2 不同处理下黏虫的幼虫发育历期Table 2 Developmental duration of M. separata under different treatments

表3 不同处理下黏虫生长发育参数Table 3 Growth and development parameters of M. separata under different treatments
3 讨论
保幼激素降解酶对保幼激素滴度的调节被认为是昆虫正常发育的关键。目前对于保幼激素降解酶的研究主要集中在JHE上,而对JHEH的研究较少。JHEH是保幼激素代谢途径中必不可少的,在保幼激素代谢过程中发挥着重要的作用[27],JHEH在黏虫中的表达情况和功能意义也有待进一步阐明。通过对黏虫转录组数据库搜索比对以及序列验证,得到1条包含完整开放阅读框序列的保幼激素环氧水解酶基因cDNA序列MsJHEH2,并对其进行生物信息学分析。MsJHEH2保守区域与其他昆虫一样具有真核生物环氧水解酶的典型结构特征,对近源物种进行同源比较,通过构建系统进化树分析表明,黏虫保幼激素环氧水解酶MsJHEH2与同属鳞翅目夜蛾科草地贪夜蛾和斜纹夜蛾中保幼激素环氧水解酶聚类到同一分支,说明其进化关系相近。MsJHEH2在黏虫各龄期与各组织中均有表达,在蛹期和中肠中表达量最高,预蛹、体壁和脂肪体中较高,这一结果与蟋蟀中的研究结果相似[28],表明了其在黏虫不同时期和不同器官内对JH的不同代谢作用,另一方面,各组织中MsJHEH2表达量的不同也可能是MsJHEH2组织特异性调节JH水平的重要因素。
保幼激素类似物烯虫酯作为一种新型生物杀虫剂有较高的生物安全性和环境安全性[29,30],本研究使用不同浓度梯度烯虫酯对黏虫处理后发现,JHA对MsJHEH2基因表达量有促进作用,由此可以推测在一定浓度范围内的保幼激素类似物处理后,昆虫体内保幼激素升高,为了维持体内的激素平衡,黏虫体内的保幼激素代谢酶活性增强来降解多余的保幼激素类似物有关,也进一步验证了MsJHEH2参与了黏虫体内保幼激素的代谢过程,表明保幼激素对黏虫生长发育的调控作用,为使用保幼激素防治黏虫提供理论基础。
目前,RNA干扰是昆虫基因功能研究领域的一项广泛应用的技术方法,特别是在昆虫靶基因功能研究中非常高效可靠[31]。通过RNA干扰技术沉默保幼激素环氧水解酶基因后,绿盲蝽若虫的存活率显著降低,并出现蜕皮阻滞,从而达到防治半翅目绿盲蝽的目的[27];RNA干扰褐飞虱保幼激素环氧水解酶会影响其短翅的形成[32],这些试验均通过RNA干扰技术证实了JHEH基因对昆虫生长发育有着重要的调控作用。本研究为进一步明确基因MsJHEH2在黏虫生长发育中的作用,利用RNA干扰技术使MsJHEH2基因沉默,对其进行生物功能研究。结果显示,RNAi介导的MsJHEH2基因沉默显著降低了基因的表达量,导致黏虫体内保幼激素含量显著增加,意味着MsJHEH2介导的 JH降解可能参与下调黏虫体内的 JH含量。干扰MsJHEH2基因后,对黏虫无明显的致死作用,且其发育历期也没有显著变化,推测由于JH降解是一个及其复杂过程,其代谢受多种酶共同调控,且黏虫体内含有多条保幼激素环氧水解酶基因,本试验对一条基因进行干扰后虽然使JH滴度一定程度上升,但并未逆转其幼虫发育历期。另一方面,RNAi后,导致其蛹历期延长,羽化率降低,因此推测MsJHEH2也可能在成虫生长发育过程中发挥重要的调控作用,这需要进一步研究以验证这一假设。
本试验发现干扰黏虫MsJHEH2基因的表达后,黏虫体内JH滴度显著上升并抑制其化蛹及羽化,说明MsJHEH2能够通过调节保幼激素含量来调控其正常的生长发育,研究结果为保幼激素环氧水解酶的表达情况和功能研究提供了重要理论依据,为达到通过分子生物学手段防治害虫、针对黏虫保幼激素代谢途径的新型杀虫剂的开发及应用提供理论基础。
