不同品种油茶果实成熟期叶片养分及磷组分的差异
2021-12-17马丽丽兰龙焱方海富牛德奎张文元胡冬南郭晓敏
马丽丽,朱 婷,兰龙焱,方海富,李 宁,张 强,牛德奎,张文元,胡冬南,郭晓敏
(江西农业大学 a.林学院;b.江西省森林培育重点实验室,江西 南昌 330045)
油茶Camellia oleifera为山茶科Theaceae 山茶属Camellia常绿小乔木或灌木,广泛分布于我国长江流域以南的红壤地区,其中以湖南、江西、广西三省(区)面积最大。油茶是我国重要的木本油料树种,其主要特点是栽培历史悠久、经济利用价值高、综合开发利用潜力大,与油橄榄、油棕、椰子并称为世界四大木本油料植物[1-3]。油茶产业是南方农村的特色产业,是乡村振兴和扶贫攻坚的支柱产业,也是我国维护国家粮油安全的新兴主力[4]。因此,提高油茶产量,促进油茶产业的发展尤其重要。
磷肥施入酸性红壤中容易被固定、沉淀,导致有效磷浓度较低,生产上投入的磷肥远远大于植物的需求,磷为江西红壤区油茶林土壤主要养分限制因子[5]。碳、氮、磷是重要的生命元素,有机体干物质的50%左右是碳,这一比例在生物的不同类群中随细胞的结构组成发生变化;地球上有机体蛋白质的16%是氮,核酸组成的9.5%是磷,这2 个比例在不同来源的生物中相对稳定[6]。生态化学计量学是研究生物系统能量平衡和多重化学元素平衡的科学,跨越了个体、种群、群落、生态系统、景观和区域各个层次[7-8]。碳、氮、磷元素含量及其生态化学计量特征的研究对揭示物种的生态策略和适应性具有重要的生态学和生理学意义[9-10]。叶片是植物进行同化作用和蒸腾作用的主要器官,叶片养分浓度能够传递植物生理生态方面的各种信息,也是对环境变化比较敏感和可塑性较大的部位[11-12]。在植物细胞中,叶片磷组分由于组成分子的大小及功能的不同,可分为几个主要组分,核酸中的磷(核酸态磷),磷脂膜中的磷(结构态磷),新陈代谢的磷(代谢态磷),不可溶解的磷(残余态磷)[13]。存在磷限制时,植物会通过优化叶片中的磷组分的分配来维持其生产力和生长,并减少对磷的需求以适应磷贫瘠的土壤[14]。在磷限制下植物吸收的磷十分有限,对植物已获得的磷进行循环利用决定了其能否在低磷环境下生存。有关学者对油茶养分的研究集中在施肥对油茶树体、果实和产量的影响[15-18],以及不同时期、不同器官营养元素的吸收、累积和分配特征等方面[19-20]。但目前对江西红壤地区不同品种油茶果实成熟期叶片养分含量差异、生态化学计量特征、以及叶片磷组分的分配的研究尚未见报道。本试验以8年生油茶‘长林23’‘长林55’‘赣无12’‘赣无16’为研究对象,探讨品种对叶片养分含量、生态化学计量特征的影响,以及不同品种油茶的磷素利用能力,为后期施肥提供指导,以提高油茶产量。
1 材料与方法
1.1 试验地概况
本试验样地位于江西省九江市永修县林木育种中心的油茶基地,油茶良种基因库占地17.13 hm2,种质资源基因库49.07 hm2,优良乡土树种收集区34.74 hm2,同纬度珍稀树种收集区41.00 hm2,林木良种生产园34.47 hm2,生态景观园28.67 hm2。年平均气温17.4℃,年平均降水量1 300~1 600 mm,属于典型的亚热带季风气候,土壤为典型红壤,缓坡地,坡度9°,是油茶的适生区。样地土壤pH 值为5.29,有机碳含量为22.99 g·kg-1,全氮含量为1.79 g·kg-1,全磷含量为1.14 g·kg-1,全钾含量为3.91 g·kg-1,有效磷含量为1.59 mg·kg-1,速效钾含量为85.57 mg·kg-1。
1.2 试验设计与方法
本试验材料为江西省林木育种中心提供的‘长林23’‘长林55’‘赣无12’‘赣无16’四个品种,树龄为8 a,已进入结果期。4 个品种都位于2012年造林的种质资源收集圃,在收集圃中每个品种随机选取9 株长势相当的油茶树作为采样目标树,品种之间设置隔离行,自造林后抚育管理措施一致。
1.3 样品采集与测定
试验于2019年10月下旬采集当年生枝(春梢)上3~4 片叶,每株油茶树分4 个方向各取2片叶子,装入自封袋中带回实验室,一部分叶片在105℃条件下杀青15 min 后于65℃下烘干,后进行养分测定。另一部分冷冻干燥,用于测定叶片磷组分,测定指标和方法如下:
叶片形态指标用手持式叶面积仪测定(型号:CI-203)。
全氮全磷测定:HClO4—H2SO4法[21],所得待测液全氮全磷用全自动间断化学分析仪(Smartchem 200 Alliance Corp.France)测定。
叶片有机碳测定[21]:重铬酸钾容量法-外加热法。
叶片磷分级:采用三氯乙酸连续浸提法[13,14]。将冷冻干燥后研磨的样品放入离心管1 中,用15 mL 12∶6∶1CMF(氯仿∶甲醇∶甲酸)提取2次,离心后将上清液转入离心管2,继续用19 mL 1∶2∶0.8CMW(氯仿∶甲醇∶水)提取2 次,将上清液转移到离心管2 中,管1 中加入9.5 mL 氯仿,混匀后将上清液转移到离心管2 中。将离心管2放入离心机离心10 min,将离心后的上层液体转移到离心管1 中,下层液体用于测定组分1。在离心管1 中加入5 mL 85%甲醇,置于真空下48 h,从水相层中除去溶解的氯仿和部分甲醇,将离心管冷却至4℃后,加入1 mL 5%TCA 溶液,再加10 mL 冷却的5%TCA 溶液进行第2 次萃取,(放入4℃冰箱,持续1 h,10 min 振荡1 次),离心后将上清液转移到离心管3(组分2)。在离心管1 中加入35 mL 2.5%TCA,放入95℃水浴锅中持续加热1 h,离心后将上清液转移到离心管4(组分3)。离心管1 中剩余的植物样品即为组分4,最后将分离出的4 种磷组分用H2SO4-H2O2消化后测定磷含量。在该过程中,结构磷、代谢磷、核酸磷和残余磷分别对应于组分1、2、3 和4。
1.4 数据处理与分析
使用Excel 2010 软件和SPSS 22.0 软件数据进行整理和方差分析以及相关性分析,用Origin 18.0软件作图。
2 结果与分析
2.1 不同品种油茶叶片形态指标分析
由表1中可得,‘赣无12’的叶面积、叶片长度、叶片宽度都显著高于‘赣无16’‘长林23’(P<0.05),‘长林23’与‘长林55’之间的叶面积、叶片长度和叶片宽度之间无显著性差异,‘长林55’与‘赣无16’的叶片长度之间也无显著性差异。‘长林55’(2.35)的叶形指数显著高于‘赣无12’(2.19)和‘赣无16’(2.16),其中‘长林55’和‘长林23’之间无显著性差异,‘赣无12’与‘赣无16’之间无显著性差异。

表1 不同品种油茶叶片形态指标差异†Table 1 Differences of morphological indexes in different Camellia varieties
2.2 不同品种油茶叶片的C、N、P 含量及化学计量特征
由表2可知,在果实成熟期,不同品种油茶比较,‘赣无12’的有机碳含量(499.90 g·kg-1)显著高于‘赣无16’(457.20 g·kg-1),‘长林23’与‘长林55’之间无显著性差异。‘赣无16’的全氮含量(17.23 g·kg-1)显著低于‘长林23’‘长林55’和‘赣无12’(19.38、19.55、19.80 g·kg-1),其中‘长林23’‘长林55’‘赣无12’三个品种之间无显著性差异。4 个品种的全磷含量无显著性差异。该结果说明品种对有机碳含量和氮含量的影响显著。

表2 不同品种油茶叶片C、N、P 含量及化学计量比Table 2 C,N,P content and stoichiometric ratio of different varieties of Camellia leaves
碳、氮、磷养分的化学计量比可以判断限制性元素和养分利用效率的高低,4 个品种的C∶N 和N∶P 之间无显著性差异。‘长林55’(395.29)与‘赣无12’(395.68)的C∶P显著高于‘长林23’(368.98)与‘赣无16’(370.57),其中‘长林55’与‘赣无12’之间无显著性差异,‘长林23’与‘赣无16’之间无显著性差异。该结果说明品种对化学计量比影响较小,仅对C∶P 影响显著。
2.3 不同品种油茶叶片磷组分及磷组分占比的差异
由图1可知,果实成熟期,不同品种比较,‘长林23’(0.21 g·kg-1)的结构磷含量显著高于‘赣无16’(0.16 g·kg-1),‘长林23’与‘赣无12’之间无显著性差异,‘长林55’与‘赣无16’之间无显著性差异。4 个品种的代谢磷含量无显著性差异。‘长林23’(0.21 g·kg-1)、‘长林55’(0.22 g·kg-1)、‘赣无12’(0.19 g·kg-1)的核酸磷含量显著高于‘赣无16’(0.16 g·kg-1),说明‘长林23’‘长林55’‘赣无12’满足在光合过程中所需蛋白质合成的核酸物质。4 个品种的残余磷含量之间无显著性差异。该结果说明不同品种油茶的磷组分含量具有差异性,品种对代谢磷和残余磷的含量影响不显著。
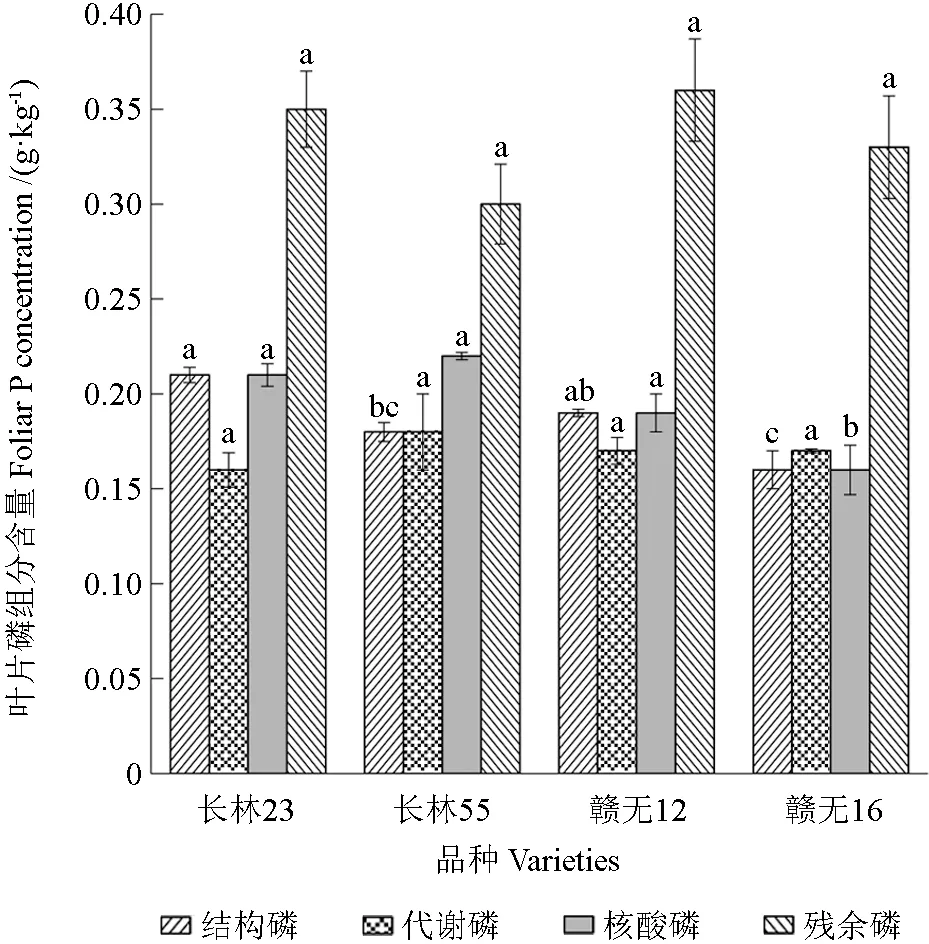
图1 不同品种油茶叶片磷组分的含量Fig.1 Difference of foliar phosphorus fraction in different Camellia varieties
由图2可得,在果实成熟期,‘长林23’叶片磷组分分配比例为:残余磷>结构磷>核酸磷>代谢磷;‘长林55’:残余磷>核酸磷>代谢磷>结构磷;‘赣无12’:残余磷>核酸磷>结构磷>代谢磷;‘赣无16’:残余磷>代谢磷>结构磷>核酸磷。‘长林23’(22.58%)的结构磷占比最高,‘赣无16’(20.03%)的最低,代谢磷占比‘赣无16’(20.45%)最高,‘长林23’(17.14%)最低;核酸磷占比‘长林55’(25.07%)的最高,‘赣无16’(19.39%)最低;残余磷占比‘赣无16’(40.13%)最高,‘长林55’(34.48%)最低,该结果说明不同品种油茶的叶片磷组分的分配方式不同。

图2 不同品种油茶叶片的磷组分占比Fig.2 Percentage of foliar phosphorus fraction in different Camellia varieties
2.4 不同品种油茶叶片形态指标与养分含量及磷组分含量之间的相关性分析
对不同品种油茶叶片的养分含量与磷组分进行相关性分析,由表3可得,叶面积与叶形指数有显著的负相关关系(P<0.05);叶片有机碳与叶片全磷有显著的正相关关系;叶片C∶N 与叶片全氮有极显著的负相关关系(P<0.01);叶片N∶P与叶片全氮有极显著的正相关关系,与叶片全磷有显著的负相关关系,与叶片的C∶N 有极显著的负相关关系,与叶片C∶P 有显著的正相关关系;结构磷与叶片全氮有显著的正相关关系;核酸磷与叶形指数、叶片全氮、叶片N∶P 有显著的正相关关系;残余磷与叶片全磷有显著的正相关关系。

表3 不同品种油茶叶片形态指标、养分含量与磷组分之间的相关性分析†Table 3 Correlation analysis among leaf morphological indexes,nutrient content and foliar phosphorus fraction in different Camellia varieties
3 讨论与结论
3.1 讨 论
叶片是植物进行光合作用的主要器官,在维持植物的生长发育方面具有不可替代的作用。植物对养分的吸收分配特性,既是植物本身的特点,也是植物的品种特点[22]。很多研究表明,叶片中的氮含量与光合作用和酶的合成过程有关[23],本试验结果表明,4 个品种油茶的叶面积和叶形指数之间存在一定的差异,叶片中的养分含量因品种不同也存在一定的差异。4 个品种的叶片全氮含量介于17.23~19.38 g·kg-1,都低于中国区域氮含量的平均值(20.20 g·kg-1)[24-25]。4 个品种叶片的全磷含量介于1.21~1.30 g·kg-1,高于中国区域的1.21 g·kg-1[24],低于全球区域的1.42 g·kg-1[26]。‘赣无16’的叶片全氮含量和有机碳含量低于‘长林23’‘长林55’和‘赣无12’,且低于全球尺度的碳含量(461.60 g·kg-1)[27],表明‘赣无16’固碳能力与光合作用能力低于其他3 个品种,主要原因可能与不同区域对不同元素的可利用性[28]以及油茶自身的遗传特性有关。植物叶片C∶N、C∶P代表了植物对氮磷元素的吸收利用能力,而N∶P临界比值是判断环境对植物生长养分供应状况的关键指标[29]。当叶片N∶P<14 时,植物生长主要受土壤氮限制;叶片N∶P>16 时,植物生长主要受土壤磷限制;当N∶P 比值介于14~16 时,植物生长受土壤中氮和磷共同限制或均不限制[30]。‘长林55’和‘赣无12’的C∶P 显著高于‘长林23’和‘赣无16’,但C∶N 之间无显著性差异,表明‘长林55’和‘赣无12’比‘长林23’和‘赣无16’有较高的养分利用效率。本研究中,4 个品种的N∶P 之间无显著性差异,但‘赣无16’的N∶P为13.86,小于14,受氮的限制;而‘长林55’的N∶P 为16.29,大于16,受到磷的限制。
植物体内的磷主要分为无机磷酸盐和有机磷酸酯,作为叶片总磷的主要部分,叶片有机磷是细胞核和细胞膜的基本组成部分,有机磷浓度在成熟叶片中相对稳定,并且在衰老之前就被水解为无机磷[31]。本试验结果表明,在果实成熟期,磷组分的分配因油茶品种不同有一定的差异,4 个品种油茶叶片的结构磷和核酸磷之间有显著性差异,而代谢磷和残余磷之间无显著性差异。研究表明,施肥会对叶片中的氮、磷含量产生一定的影响,叶片N∶P 与植物生长速率相关,叶片N∶P随叶片磷浓度的降低而增加,生态化学计量学的研究重点是将磷分配到rRNA 上,是控制叶片N∶P变化的重要因素[32]。结构态磷是植物细胞磷脂膜中的磷,属于难溶态的磷;核酸磷是核酸中含有的磷,合成光合作用所必需的蛋白质,存在磷限制时,植物会将更多的磷分配给核糖体RNA,用来合成蛋白质,使得核酸磷的含量升高。不同的植物具有不同的磷吸收和利用策略,物种间的差异最终带来植物磷组分分配的差异。有学者的研究表明,在低磷胁迫下,磷高效植株通过提高下部叶酸性磷酸酶活性加强酯磷和核酸态磷的分解,分解的磷被上部新生叶片再利用,以提高后期磷素的再利用能力[33-34]。Wieneke 等[35]的研究表明,低磷胁迫下,高粱磷高效品种叶片中TCA(三氯乙酸)可溶性磷含量迅速下降,在该组分完全耗尽前,TCA 不溶性磷组分也显著降低,根中总磷含量基本保持不变,不溶性组分有明显的变化。在本试验中,核酸磷含量与叶片全氮含量和N∶P 呈显著正相关,结构磷含量与叶片全氮呈显著正相关,表明核酸磷和结构磷与全氮的关系较为紧密。4 个品种的全磷含量无显著性差异,但‘长林23’的叶片比其他3 个品种能维持较高的结构磷和核酸磷含量,说明‘长林23’比‘长林55’‘赣无12’‘赣无16’有更高的磷利用能力。有研究表明,土壤氮素的增加提高了丝栗栲幼树结构磷的比例,降低了代谢磷的比例[36],本研究中‘赣无16’的结构磷和核酸磷含量最低,后续可在果实生长高峰期通过施氮肥提高‘赣无16’油茶叶片中结构磷和核酸磷的含量。残余态磷主要是一些未被识别的含磷蛋白,是不可溶解的磷,受基因型差异的影响较小。许多研究表明,植物组织中的全磷浓度对可利用的土壤磷和植物生长的变化无响应,而植物组织中的无机磷浓度提供了植物中磷状况的最佳指标[37-38],本试验中,残余磷与叶片全磷的关系较为紧密。因此,不能仅以全磷作为营养诊断的指标,要考虑各磷组分对全磷的响应,这与刘兴宇等[39]的研究结果相似。
由于土壤磷有效度低,植株通常会降低叶片的磷浓度。植物细胞内的磷酸酶能够水解植物体内的酯磷化合物,可加快磷的代谢速率[33],而不同品种油茶叶片的磷酸酶与磷组分的关系需进一步深入研究。较低的叶片磷浓度与叶片性状[40]及自身的遗传特性有关,所以对油茶的后续研究应注重叶片磷吸收效率和磷功能基因的关系,筛选磷高效基因型油茶,提高油茶体内磷素的利用能力,以克服南方红壤磷素缺乏的养分限制因子的不利影响。
3.2 结 论
在果实成熟期,品种对油茶叶片的碳含量、氮含量、C∶P、结构磷和核酸磷含量有显著影响,对全磷和N∶P、C∶N、代谢磷和残余磷无显著影响;‘长林23’的磷利用能力最高,‘长林55’和‘赣无12’次之,‘赣无16’最低。因此,在油茶施肥和养分管理过程中,应根据油茶品种的叶片养分特性和磷组分差异调整施肥比例,科学合理增施氮肥和磷肥,以克服南方红壤磷素缺乏的养分限制因子的不利影响,提高油茶产量。
