54份可可种质资源主要品质性状及相关分析
2021-12-15谢薇赖剑雄秦晓威朱自慧李付鹏
谢薇 赖剑雄 秦晓威 朱自慧 李付鹏

摘要:【目的】對54份可可种质材料的主要品质性状进行比较分析,筛选出品质性状优良的种质,为可可新品种培育提供基础材料,也为探究可可品质性状的影响因素提供理论参考。【方法】以来源于不同国家的54份可可种质为材料,采用超声波超离法、气相色谱法和福林酚法测定可可脂、多酚、脂肪酸组分(棕榈酸、硬脂酸、油酸和亚油)含量及脂肪酸总含量,并对其进行变异分析、相关分析及聚类分析。【结果】54份可可种质中,XYS53的可可脂、棕榈酸、硬脂酸、油酸、亚油酸含量及脂肪酸总含量均最低;XYS52的可可脂含量和脂肪酸总含量均最高;XYS18的棕榈酸含量最高;XYS17的硬脂酸含量最高;XYS52的油酸含量最高;XYS44的亚油酸含量最高;XYS20的多酚含量最高。7个品质性状指标的变异系数为11.52%~18.74%,其中棕榈酸、亚油酸和多酚的变异幅度较大,变异系数分别为18.74%、18.62%和14.42%;可可脂含量变异系数最小,为11.52%。可可脂含量与棕榈酸、硬脂酸和油酸含量呈极显著正相关(P<0.01,下同),棕榈酸含量与硬脂酸含量间、棕榈酸含量与油酸含量间及硬脂酸含量与油酸含量间均呈极显著正相关;亚油酸含量与棕榈酸含量存在显著负相关(P<0.05,下同);多酚含量与可可脂、棕榈酸、硬脂酸、油酸和亚油酸含量均呈负相关,但未达显著水平(P>0.05,下同)。不同国家的可可种质间可可脂、硬脂酸、油酸、多酚含量及脂肪酸总含量无显著差异,但中国与泰国的可可种质间棕榈酸含量和亚油酸含量存在显著差异。主成分分析和聚类分析均发现,归属于类群Ⅰ的15份种质品质性状综合表现较好,且主要来自于中国和越南。【结论】可可种质的主要品质性状与其来源地间存在一定相关性。在可可品种选育过程中,需结合性状间相关性综合考虑可可品质性状表现,可优先选择中国和越南的15个综合性状优良的种质作为育种材料或亲本来培育优良可可品种。
关键词: 可可;鉴定评价;可可脂;脂肪酸;多酚
中图分类号: S571.3 文献标志码: A 文章编号:2095-1191(2021)08-2174-09
Evaluation and correlation analysis of main quality traits of 54 cacao germplasm resources
XIE Wei1,2, LAI Jian-xiong2, QIN Xiao-wei2, ZHU Zi-hui2, LI Fu-peng2*
(1Institute of Horticulture,Hainan University,Haikou 570208, China; 2Institute of Spices and Beverages,Chinese Academy of Tropical Agricultural Sciences/ Hainan Provincial Key Laboratory of Genetic Improvement and Quality Regulation for Tropical Spice and Beverage Crops/Key Laboratory of Genetic Resources for Spiced Beverage Crops, Ministry of Agriculture and Rural Affairs,Wanning,Hainan 571533, China)
Abstract:【Objective】In order to select excellent cacao germplasms with better quality traits, evaluation and correlation analysis were performed by 54 cacao core collections. The results could provide basic materials for cultivation of new cacao varieties, and supply a theoretical basis for the formation of cacao quality traits. 【Method】The main quality traits,cocoa butter content, polyphenol, fatty acid components(palmitic acid content, stearic acid content, oleic acid content, linoleic acid) and total content of fatty acids were performed with 54 different origins cacao germplasms from different countries, by ultrasonic ultra-isolation method, gas chromatography and Folin-Ciocalteus method. Variability analysis, correlation analysis and cluster analysis were also performed. 【Result】Among 54 cacao germplasms, XYS53 showed the lowest cocoa butter content, palmitic acid content, stearic acid content, oleic acid content, linoleic acid content and total fatty acid content. XYS52 showed the highest cocoa butter content and total fatty acid content. XYS18, XYS17, XYS52, XYS44 and XYS20 showed highest phenotype on palmitic acid content, stearic acid content, oleic acid content, linoleic acid content and polyphenol content, respectively. The average of variation coefficient was from 1.52% to 18.74 %, of which palmitic acid, linoleic acid and polyphenols showed larger variations with 18.74%, 18.62% and 14.42%. The variation coefficient of cocoa butter content was the smallest(11.52%). Correlation analysis showed that cocoa butter content was extremely significantly positively correlated with palmitic acid, stearic acid and oleic acid(P<0.01, the same below). There was an extremely significant positive correlation between palmitic acid and stearic acid, between palmitic acid and oleic acid, and between stearic acid and oleic acid. There was significant negative correlation between linoleic acid and palmitic acid (P<0.05, the same below). While polyphenols were negatively correlated with cocoa butter, palmitic acid, stearic acid, oleic acid and linoleic acid contents, but did not reach a significant level(P>0.05, the same below). There were no significant differences among cocoa butter content, stearic acid content, oleic acid content, polyphenol content and four fatty acids contents of different germplasm origin countries. However, there were significant differences of palmitic acid and linoleic acid contents between germplasms from China and Thailand. The results of principal component analysis and cluster analysis indicated that quality traits of 15 germplasm belonging to Group I performed well, these germplasms were maily from China and Vietnam. 【Conclusion】There is certain correlation between the main quality traits and different cacao germplasm origins. In the breeding process of selecting cocao varieties, by combining the correlation between traits and comprehensively considering the performance of cocao quality traits, 15 germplasms with excellent comprehensive traits from China and Vietnam can be preferred as breeding materials or parents to cultivate excellent cocao varieties.
Key words: cacao; identification and evaluation; cocoa butter; fatty acid; polyphenol
Foundation item: National Natural Science Foundation of China(31670684); Central Public-interest Scientific Institution Basal Research Fund for Chinese Academy of Tropical Agricultural Sciences(1630142019003)
0 引言
【研究意义】可可(Theobroma cacao L.)是生长于热带地区的常绿小乔木,与茶和咖啡并称为世界三大饮料作物(Selmi et al.,2006;李付鹏等,2014),广泛栽培于世界热带地区,在我国主要分布于海南省和云南省。可可的种子(即可可豆)富含可可脂、多酚类化合物、蛋白质及膳食纤维等活性成分,营养丰富,味醇香,是巧克力、糕点、糖果等食品的重要原料,在国际大宗商品贸易中占有重要地位(Motamayor et al.,2013;李付鹏等,2016,2018)。其中,可可脂作为可可豆的主要经济成分,具有独特的理化性质,熔点为35~37 ℃,其含量是决定巧克力品质的重要指标;多酚类化合物具抗氧化、预防心脑血管疾病功能(Shekarchizadeh et al.,2014;Nasiruddin et al.,2014)。因此,筛选鉴定可可脂和多酚含量特异的可可种质资源,可为培育优良品质性状的品种提供基础材料(Ellam and Williamson,2013;周波,2017),对可可产业健康发展具有重要意义。【前人研究进展】目前,国内外学者从可可品质性状鉴定评价及其形成机制方面开展研究。Chaiseri和Dimick(1989)研究发现美洲、亚洲和非洲的可可种质间可可脂含量存在明显差异,且南美洲可可脂特性和组分与北美洲和中美洲的可可脂特性相近,但与亚洲和非洲的可可脂特性存在明显差异。Steinberg等(2003)、Ellam和Williamson (2013)研究发现,可可脂含量约占可可豆干重的50%,可可脂中主要的脂肪酸为棕榈酸、硬脂酸、油酸及少量的亚油酸。Torres-Moreno等(2015)对不同产地的可可和巧克力营养成分与脂肪酸组成进行比较分析,结果发现可可脂肪酸组分因不同地理来源而存在明显差异。Urbańska和Kowalska(2019)比较了不同地区烘焙和未烘焙可可豆的多酚含量,结果发现多酚含量受基因型和生长地区环境的影响,其中南美洲哥伦比亚和秘鲁可可豆的多酚含量最高。此外,研究证实,多酚类物质是可可豆的重要组成成分,占可可豆干重的5%~6%(Bertazzo et al.,2011;Kruszewski and Obiedzinski,2018);多酚类物质是形成可可和巧克力涩味的主要成分,主要以儿茶素、表儿茶素、花青素和原花青素形式存在(Borchers et al.,2000;Sanchez-Rabaneda et al.,2003;Jolic et al.,2011)。【本研究切入点】虽然已对世界可可主产区的可可豆中可可脂含量进行比较分析,但鲜见基于主要品质性状进行可可种质的综合评价及分析不同来源地(东南亚与大洋洲)对主要品质性状影响的研究报道。【拟解决的关键问题】以54份可可核心种质为材料,对其可可脂含量、多酚含量、4种脂肪酸组分含量及脂肪酸总含量等品质性状进行鉴定评价及相关分析,并分析种质来源地与主要品质性状间相关性,筛选出综合性状优良的可可种质,为可可新品种培育提供基础材料。
1 材料与方法
1. 1 试验材料
以54份可可核心种质为供试材料,保存于中国热带农业科学院香料饮料研究所农业农村部热带香料饮料作物种质资源圃,其来源地如表1所示。
1. 2 样品采集及处理
在可可果实成熟期,分别选取同一株树干上不同位置的3个生长状况均一、正常發育的成熟果实,将每个可可果实单独处理,剖开果实取出种子洗净后,于50 ℃恒温烘箱干燥48 h,充分粉碎后过40目筛。并对3个可可样品的可可脂含量、多酚含量、脂肪酸组分含量等6个品质性状进行测定,计算平均值。3个可可样品即为同一份种质材料的3次重复,各品质性状的平均值即为该份可可种质的品质性状表型。
1. 3 品质性状测定
1. 3. 1 可可脂含量测定 参考范睿等(2019)的方法提取可可脂。具体步骤:(1)称取可可粉末2 g左右(记为M1),置于干燥洁净的50 mL离心管中再次称量(记为M2);(2)向离心管中加入石油醚,在超声波清洗机中浸提10 min,取出离心管,冷却至室温;(3)将离心管放入转速8000 r/min的离心机中离心后弃上清液,重复提取并离心2次;(4)弃去上清液样品在通风橱挥发多余石油醚后放入95 ℃的鼓风干燥箱内干燥4 h,冷却后称量(记为M3),运用减重法计算得出可可脂含量(X),计算公式为X=(M2-M3)/M1。
1. 3. 2 脂肪酸含量测定 参照农业行业标准(NY/T 3110─2017),采用气相色谱法规定棕榈酸、硬脂酸、油酸和亚油酸4种主要脂肪酸组分含量及脂肪酸总含量。
1. 3. 3 多酚含量测定 参照汤雯(2012)的福林酚法测定可可多酚含量。
1. 4 统计分析
利用Excel 2003对品质性状的平均值、标准差及变异系数进行分析,并利用SPSS 22.0对品质性状进行相关分析、主成分分析与聚类分析。
2 结果与分析
2. 1 可可种质材料的品质性状分析结果
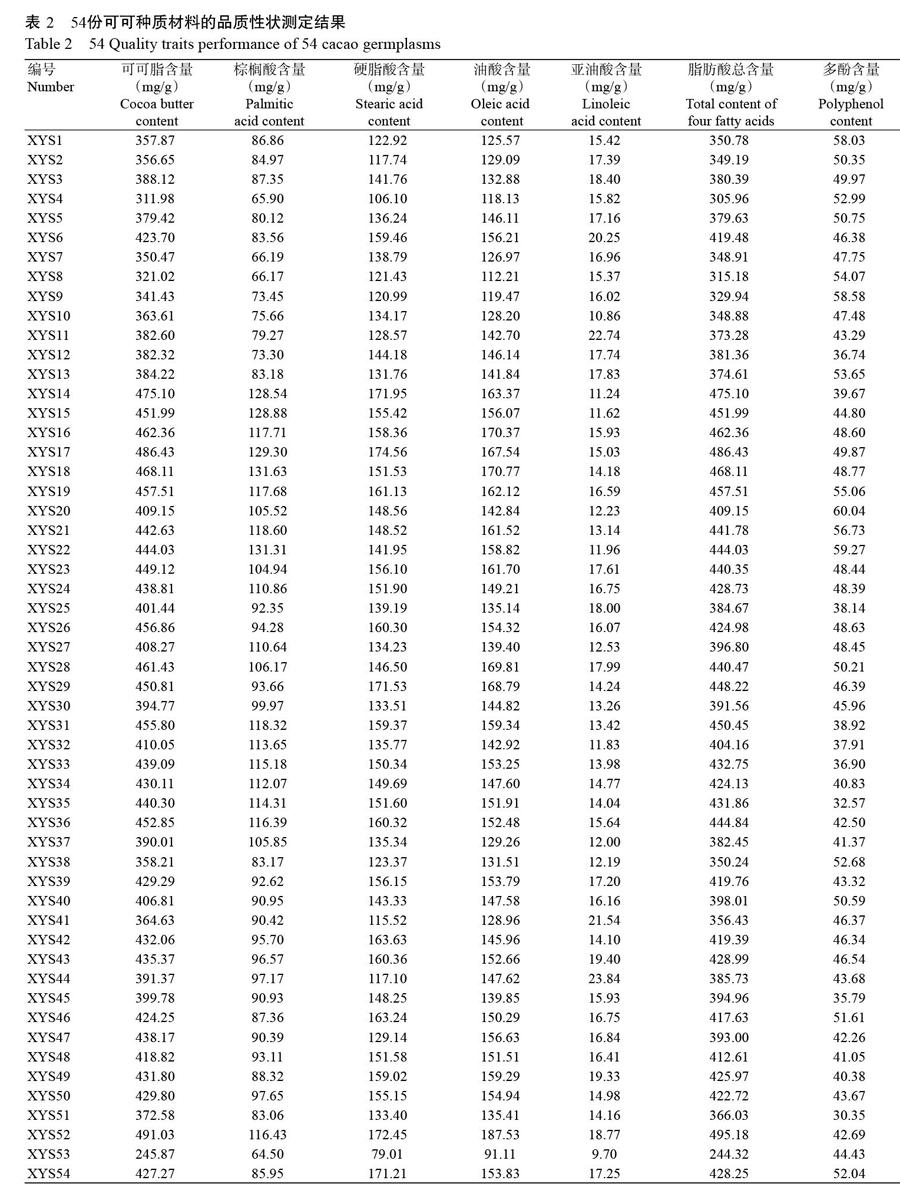
由表2可知,54份可可种质材料中,XYS53的可可脂、棕榈酸、硬脂酸、油酸、亚油酸含量及脂肪酸总含量均最低,分别为245.87、64.50、79.01、91.11、9.70和244.32 mg/g;XYS52的可可脂含量、油酸含量和脂肪酸总含量均最高,分别为491.03、187.53和495.18 mg/g;XYS18的棕榈酸含量最高,为131.63 mg/g;XYS17的硬脂酸含量最高,为174.56 mg/g;XYS44的亚油酸含量最高,为23.84 mg/g;XYS20的多酚含量最高,为60.04 mg/g。
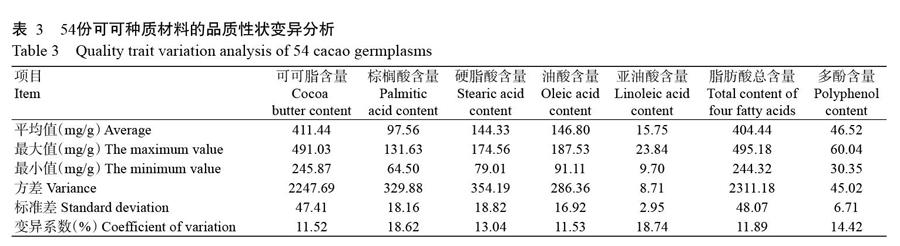
54份可可种质材料的品质性状变异情况如表3所示。可可脂含量为245.87~491.03 mg/g,多酚含量为30.35~60.04 mg/g,棕榈酸、硬脂酸、油酸和亚油酸含量分别为64.50~131.63 mg/g、79.01~174.56 mg/g、91.11~187.53 mg/g和9.70~23.84 mg/g。7个指标的变异系数为11.52%~18.74%,其中棕榈酸、亚油酸和多酚含量的变异幅度较大,变异系数分别为18.62%、18.74%和14.42%;脂肪酸总含量的变异系数较小,为11.89%;可可脂变异系数最小,为11.52%。可见,可可脂、多酚和总脂肪酸含量变异系数排序为:多酚(14.42%)>脂肪酸总含量(11.89%)>可可脂(11.52%),其中4种脂肪酸组分含量变异系数排序为:亚油酸(18.74%)>棕榈酸(18.62%)>硬脂酸(13.04%)>油酸(11.53%)。
2. 2 品质性状的相关分析结果
由表4可知,可可脂含量与棕榈酸、硬脂酸和油酸含量呈极显著正相关(P<0.01,下同);棕榈酸含量与硬脂酸含量间、棕榈酸含量与油酸含量间及硬脂酸含量与油酸含量间均呈极显著正相关;亚油酸含量与棕榈酸含量存在显著负相关(P<0.05,下同);亚油酸含量与可可脂、硬脂酸和油酸含量呈正相关,但未达显著水平(P>0.05,下同);多酚含量与可可脂、棕榈酸、硬脂酸、油酸和亚油酸含量均呈负相关,但未达显著水平。
2. 3 不同来源地的可可种质材料品质性状方差分析结果
不同来源地的可可种质间品质性状进行方差分析,结果如表5所示。不同来源地的可可种质材料在可可脂、硬脂酸、油酸、多酚含量及脂肪酸总含量方面均无显著差异,但泰国和中国可可种质的棕榈酸和亚油酸含量存在顯著差异,其中,中国的可可棕榈酸含量平均值显著高于泰国的可可棕榈酸含量平均值,泰国的可可亚油酸含量平均值显著高于中国的可可亚油酸含量平均值。
2. 4 品质性状的主成分分析结果
对54份可可种质材料的6个品质性状进行主成分分析,结果如表6所示。6个品质性状综合为3个主要成分,累积贡献值为92.094%,说明这3个主成分包含了可可品质性状的主要信息。主成分1的特征值为3.404,贡献值为56.739%;主成分2的特征值为1.151,贡献值为19.184%;主成分3的特征为0.970,贡献值为16.171%。根据主成分分析中因子旋转后的载荷矩阵可知,主成分1中可可脂(0.991)、棕榈酸(0.812)、硬脂酸(0.896)和油酸(0.949)的指标值较大,表示主成分1主要代表可可脂和3种主要的脂肪酸;主成分2中亚油酸的指标值最大(0.979),表示主成分2主要代表亚油酸;主成分3中多酚的指标值(0.997)最大,表示主成分3主要代表多酚。3个主成分分别用Y1、Y2和Y3表示,根据SPSS 22.0分析得出的得分系数矩阵可知,3个主成分与可可6个品质性状间的关系:
Y1=0.296X1+0.234X2+0.268X3+0.292X4+
0.038X5+0.059X6
Y2=0.031X1+(-0.335)X2+0.079X3+0.149X4+
0.855X5+0.016X6
Y3=0.010X1+0.035X2+(-0.003)X3+0.042X4+
0.017X5+1.006X6
其中,X1~X6为所有标准化的原始变量。
结合贡献率可得出品质性状综合得分与3个主成分之间的关系:Y=0.567Y1+0.192Y2+0.162Y3。根据品质性状综合得分公式,对54份可可种质进行综合评价,得到综合得分,并将综合得分前15的种质材料列于表7。可见,XYS52、XYS17、XYS14等15个可可种质综合性状表现较好,且主要来自于中国和越南。
2. 5 聚类分析结果
采用欧氏距离类间平均聚类法对54份可可种质材料进行聚类分析,并对各类群的品质性状含量平均值进行统计分析,结果如图1和表8所示。在欧氏距离为3时,54份可可种质材料被聚为四大类群,其中类群I包含28份来自于越南、泰国、巴布亚新几内亚和中国的可可种质,该类群特点:可可脂、棕榈酸、硬脂酸和油酸含量及脂肪酸总含量平均值均最高,分别为446.67、108.11、158.33、158.75和440.86 mg/g;类群Ⅱ包含16份来自于越南、泰国、巴布亚新几内亚和中国的可可种质,该类群特点:亚油酸含量平均值最高,为16.29 mg/g;类群Ⅲ有9份来自于泰国、越南和中国的可可种质,多酚含量平均值最高,为52.03 mg/g;类群Ⅳ只含1份来自于越南的可可种质,其可可脂含量、4种脂肪酸组分含量及脂肪酸总含量均最低。结合主成分分析结果可知,综合品质性状优良的15份可可种质材料均聚于类群I,可作为优异的可可育种基础材料。
3 讨论
中国热带农业科学院香料饮料研究所对世界主产区的可可种质资源进行全面收集保存(李付鹏等,2014)。本研究对这些收集保存的可可种质材料进行品质性状分析,结果发现其具有丰富的表型遗传变异性,变异系数以亚油酸含量最高,其次为棕榈酸含量和多酚含量。此外,研究发现XYS52的可可脂含量和脂肪酸总含量最高,且综合性状最优,是可可脂含量特异的优良种质;XYS53的可可脂、4种脂肪酸组分含量及脂肪酸总含量均为最低,尤其是可可脂含量非常低,仅为可可脂含量平均值的50%左右,是可可脂和脂肪酸含量特异的可可种质,可作为品质性状特异性研究的特殊材料(Mustiga et al.,2019;Urbańska and kowalska,2019)。可可脂中主要的饱和脂肪酸为棕榈酸(含量约为24.5%~33.7%)和硬脂酸(含量约为33.7%~40.2%),主要的不饱和脂肪酸为油酸(含量约为26.3%~35.0%)和亚油酸(含量约为1.7%~3.0%),且低饱和脂肪酸和高不饱和脂肪酸的可可常用于食品工业(Mozaffarian et al.,2010;Bindu and Vijay,2014;Mustiga et al.,2019)。XYS44的棕榈酸和硬脂酸的总含量最低,油酸和亚油酸的总含量最高,因此XYS44可作为低饱和脂肪酸、高不饱和脂肪酸的特殊可可种质。由此可见,54份可可种质材料的6个主要品质性状具有较大的选择空间,可用于拓宽可可种质资源遗传背景及品质性状遗传改良。
可可为常异花授粉作物,其果实品质性状受多个基因的调控,且不同品质性状间存在一定的相关性(Jan and Elke,2000;Mustiga et al.,2019)。由于葡萄糖分解代谢生成的乙酰辅酶A参与脂肪酸合成途径和多酚合成途径,因此,脂肪酸和可可脂与多酚存在一定的负相关性(Jan and Elke,2000;Beppu et al.,2013)。结合本研究可可品质性状的相关分析结果可知,可可脂含量与棕榈酸、硬脂酸和油酸含量呈极显著正相关,说明可通过改良可可种质的脂肪性状提高脂肪酸组分含量;多酚含量与可可脂含量和4种脂肪酸组分含量均呈负相关性,与根据脂肪酸和多酚合成途径所得结论相一致,因此,筛选高多酚含量的可可种质为育种材料时,应关注其可可脂和4种脂肪酸组分含量。可见,在选育优异可可品种过程中,需结合性状间相关性,综合考虑可可品质性状表现。
前人研究已证实,可可品质性状间呈一定的相关性,且品质性状与来源地存在一定的关联(Urbańska et al.,2019)。但本研究通过分析不同国家可可种质的品质性状差异,发现不同来源地的可可种质在可可脂、硬脂酸、油酸、多酚含量及脂肪酸总含量方面均无显著差异,但来自中国与泰国的可可种质间棕榈酸和亚油酸含量存在显著差异。由于可可脂含量与遗传物质相关,因此在以后研究中可通过转录分析筛选不同国家可可种质中与可可脂相关的差异基因,以进一步探寻可可脂含量无显著差异的分子机制(Mustiga,et al.,2019)。来自中国与泰国的可可种质间棕榈酸和亚油酸含量存在显著差异可能是由于遗传变异所致(Bucheli et al.,2001)。今后可通过基因水平检测分析遗传变异是否为导致中国与泰国可可种质亚油酸含量差异的主要因素。多酚含量不仅与脂肪酸组分相关联,还取决于可可种质的基因型、产地及生长条件。此外,Urbańska和Kowalska(2019)对不同地区烘焙和未烘焙可可豆巧克力中总多酚含量进行比较分析,结果发现来自厄瓜多尔、哥伦比亚和委内瑞拉等美洲国家的可可种质多酚含量间存在显著差异,表明不同地理环境会影响多酚含量(Rusconi and Conti,2010;Katz et al.,2011;Kothe et al.,2013)。但本研究中得出中国、越南、泰国和巴布亚新几内亚的可可种质多酚含量无显著差异。造成与前人研究结论存在差异的原因可能是本研究的可可种质来源于东南亚和大洋洲国家,其地理环境与美洲国家存在明显差异。
本研究根据品质性状综合表现将可可种质分为四大类群,其中综合性状较优的15份可可种质均聚于类群I,且大多数来自于中国和越南,说明中国和越南的可可种质比巴新和泰国可可种质的综合品质性状更优良。由于可可品质性状易受基因、环境等因素影响,今后应结合转录分析对可可品质性状开展鉴定评价。
4 结论
可可种质品质性状间具有一定的相关性,且可可种质的主要品质性状表现与其来源地间显著相关。因此在品种选育过程中,需综合考虑可可品质性状表现,并可优先筛选来自中国或越南的种质,作为育种材料或亲本来培育优良可可品种。
参考文献:
范睿,陈冬梅,周利萍,龙雪冰,郑俏然. 2019. 胡椒脂肪的快速提取方法研究[J]. 中国热带农业,(3):42-46. doi:10. 3969/j.issn.1673-0658.2019.03.012. [Fan R,Chen D M,Zhou L P,Long X B. Zheng Q R. 2019. Study on rapid extraction of fat in black piper[J]. China Tropical Agriculture,(3):42-46.]
李付鹏,秦晓威,郝朝运,闫林,伍宝朵,赖剑雄. 2016. 可可核心种质遗传多样性及果实性状与SSR标记关联分析[J]. 热带作物学报,37(2):226-233. doi:10.3969/j.issn.1000-2561.2016.02.003. [Li F P,Qin X W,Hao C Y,Yan L,Wu B D,Lai J X. 2016. Genetic diversity and association analysis of cacao germplasm using SSR markers[J]. Journal of Tropical Crops,37(2):226-233.]
李付鹏,谭乐和,秦晓威,郝朝运,宋应辉,赖剑雄. 2018. 可可嫁接成活率研究[J]. 种子,37(2):94-97. doi:10.16590/j.cnki.1001-4705.2018.02.094. [Li F P,Tan L H,Qin X W,Hao C Y,Song Y H,Lai J X. 2018. Study on grafting survival rate of cacao tree[J]. Seed,37(2):94-97.]
李付鵬,王华,伍宝朵,赵溪竹,秦晓威,赖剑雄. 2014. 可可果实主要农艺性状相关性及产量因素的通径分析[J]. 热带作物学报,35(3):448-453. doi:10.3969/j.issn.1000-2561.2014.03.006. [Li F P,Wang H,Wu B D,Zhao X Z,Qin X W,Lai J X. 2014. Correlation analysis among main pod agronomic traits and path analysis on yield factor in cacao[J]. Journal of Tropical Crops,35(3):448-453.]
汤雯. 2012. 可可多酚主要成分鉴定及其体外抗氧化活性评价[D]. 杭州:浙江大学. [Tang W. 2012. Verification and analysis of in vitro antioxidation activity of main ingre-dients in cocoa polyphenols[D]. Hangzhou:Zhejiang University.]
周波. 2017. 海南可可中多酚和可可堿的提取分离技术及成分分析研究[D]. 湛江:广东海洋大学. [Zhou B. 2017. Study on extraction separation technology and component analysis of polyphenol and theobromine in Theobroma cacao(Hainan)[D]. Zhanjiang:Guangdong Ocean University.]
Beppu F,Nagai T,Yoshinaga K,Mizobe H,Kojima K,Gotoh N. 2013. Quantification of triacylglycerol molecular species in cocoa butter using high-performance liquid chromatography equipped with nano quantity analyte detector[J]. Journal of Oleo Science,62(10):789-794. doi:10.5650/ jos.62.789.
Bertazzo A,Agnolin F,Comai S,Zancato M,Costa C V L,Seraglia R,Traldi P. 2011. The protein profile of Theobroma cacao L. seeds as obtained by matrix-assisted laser desorption/ionization mass spectrometry[J]. Rapid Communications in Mass Spectrometry,25(14):2035-2042. doi:10.1002/rcm.5080.
Bindu N,Vijay K. 2014. Cocoa butter and its alternatives:A review[J]. Journal of Bioresource Engineering and Technology,1(3):7-17.
Borchers A T,Keen C L,Hannum S M,Gershwin M E. 2000. Cocoa and chocolate:Composition,bioavailability,and health implications[J]. Journal of Medicinal Food,3(2):77-105. doi:10.1017/S0029665116000835.
Bucheli P,Rousseau G,Alvarez M,Laloi M,McCarthy J. 2001. Developmental variation of sugars,carboxylic acids,purine alkaloids,fatty acids,and endoproteinase activity during maturation of Theobroma cacao L. seeds[J]. Journal of Agricultural and Food Chemistry,49(10):5046-5051. doi:10.1021/jf010620z.
Chaiseri S,Dimick P S. 1989. Lipid and hardness characteristics of cocoa butters from different geographic regions[J]. Journal of the American Oil Chemists Society,66(12):1771-1780. doi:10.1007/BF02660745.
Ellam S,Williamson G. 2013. Cocoa and human health[J]. Annual review of nutrition,33(1):105-128. doi:10.1146/annurev-nutr-071811-150642.
Jan W,Elke A. 2000. Review on polyphenols in Theobroma cacao:Changes in composition during the manufacture of chocolate and methodology for identification and quantification[J]. Food Research International,33(6):423-447. doi:10.1016/S0963-9969(00)00068-5.
Jolic S M,Redovnikovic I R,Markovic K,Sipusic I D,Delonga K. 2011. Changes of phenolic compounds and antioxidant capacity in cocoa beans processing[J]. International Journal of Food Science and Technology,46(9):1793-1800. doi:0.1111/j.1365-2621.2011.02670.x
Katz D L,Doughty K,Ali A. 2011. Cocoa and chocolate in human health and disease[J]. Antioxidants & Redox Signaling,15(10):2779-2811. doi:10.1089/ars.2010.3697.
Kothe L,Zimmermann B F,Galensa R. 2013. Temperature influences epimerization and composition of flavanol monomers,dimers and trimers during cocoa bean roasting[J]. Food Chemistry,141(4):3656-3663. doi:10.1016/j.foodchem.2013.06.049.
Kruszewski B,Obiedzinski M W. 2018. Multivariate analysis of essential elements in raw cocoa and processed chocolate mass materials from three different manufacturers[J]. Lwt-Food Science and Technology,98(12):113-123. doi:10.1016/j.lwt.2018.08.030.
Motamayor J C,Mockaitis K,Schmutz J,Haiminen N,Livingstone D,Cornejo O,Findley S D,Zheng P,Utro F,Royaert S,Saski C,Jenkins J,Podicheti R,Zhao M X,Scheffler B E,Stack J C,Feltus F A,Mustiga G M,Amores F,Phillips W,Marelli J P,May G D,Shapiro H,Ma J X,Bustamante C D,Schnell R J,Main D,Gilbert D,Parida L,Kuhn D N. 2013. The genome sequence of the most widely cultivated cacao type and its use to identify candidate genes regulating pod color[J]. Genome Biology,14(6):1-25. doi:10.1186/gb-2013-14-6-r53.
Mozaffarian D,Micha R,Wallace S. 2010. Effects on coronary heartdisease of increasing polyunsaturated fat in place of saturated fat:A systematic review and meta-ana-lysis of randomized controlled trials[J]. PLoS Medicine,7(3):1-8. doi:10.1371/journal.pmed.1000252.
Mustiga G M,Morrissey J,Stack J C,DuVal A,Royaert S,Jansen J,Bizzotto C S,Villeladias C,Mei L K,Cahoon E B,Seguine E,Marelli J P,Motamayor J C. 2019. Identification of climate and genetic factors that control fat content and fatty acid composition of Theobroma cacao L. beans[J]. Frontiers in Plant Science,10(10):1159. doi:10.3389/fpls.2019.01159. eCollection 2019.
Nasiruddin K,Olha K,Mireia U S,Sara T,Mar G A,María M,Ximena M C,Rafael L,Cristina A L. 2014. Cocoa polyphenols and inflammatory markers of cardiovascular disease[J]. Nutrients,6(2):844-880. doi:10.3390/nu60 20844.
Rusconi M,Conti A. 2010. Theobroma cacao L. the Food of the Gods:A scientific approach beyond myths and claims[J]. Pharmacological Research,61(1):5-13. doi:10.1016/j.phrs.2009.08.008.
Sanchez-Rabaneda F,Jauregui O,Casals I,Andrés-Lacueva C,Izquierdo-Pulido M,Lamuela-Raventós R M. 2003. Liquid chromatographic/electrospray ionization tandem mass spectrometric study of the phenolic composition of cocoa (Theobroma cacao)[J]. Journal of Mass Spectrometry,38(1):35-42. doi:10.1002/jms.395.
Selmi C,Mao T K,Keen C L,Schmitz H H,Eric G M. 2006. The anti-inflammatory properties of cocoa flavanols[J]. Journal of Cardiovascular Pharmacology,47(S2):S163-S176. doi:10.1097/00005344-200606001-00010.
Shekarchizadeh H,Tikani R,Kadivar M. 2014. Optimization of cocoa butter analog synthesis variables using neural networks and genetic algorithm[J]. Journal of Food Scien-ce and Technology,51(9):2099-2105. doi:10.1007/s13197- 012-0695-y.
Steinberg F M,Bearden M M,Keen C L. 2003. Cocoa and chocolate flavonoids:Implications for cardiovascular health[J]. Journal of the American Dietetic Association,103(2):215-223. doi:10.1053/jada.2003.50028.
Torres-Moreno M,Torrescasana E,Salas-Salvadó J,Blanch C. 2015. Nutritional composition and fatty acids profile in cocoa beans and chocolates with different geographical origin and processing conditions[J]. Food Chemistry,166(1):125-132. doi:10.1016/j.foodchem.2014.05.141.
Urbańska B,Kowalska J. 2019. Comparison of the total polyphenol content and antioxidant activity of chocolate obtained from roasted and unroasted cocoa beans from different regions of the world[J]. Antioxidants(Basel,Switzerland),8(8):283. doi:10.3390/antiox8080283.
(責任编辑 陈 燕)
收稿日期:2020-07-21
基金项目:国家自然科学基金面上项目(31670684);中国热带农业科学院基本科研业务费专项(1630142019003)
通讯作者:李付鹏(1984-),https://orcid.org/0000-0001-7521-3412,博士,副研究员,主要从事热带作物种质资源与遗传育种研究工作,E-mail:peterlfp@163.com
第一作者:谢薇(1996-),https://orcid.org/0000-0002-7610-6202,研究方向为种质资源与遗传育种,E-mail:mariaxw@163.com
