桑基鱼塘池塘子系统氮、磷污染特征及迁移转化研究进展
2021-12-15陈金洁章昕颖刘科学
程 瑶,陈金洁,谢 湘,章昕颖,刘科学,戴 军,张 池
(1.广州新华学院资源与城乡规划系,广东 广州 510520;2.广东华南空间规划研究院,广东 广州 510630;3.华南农业大学资源环境学院,广东 广州 510642)
桑基鱼塘系统是我国传统的生态农业生产模式,其所形成的陆基种桑、桑叶养蚕、蚕沙喂鱼、塘泥培桑的良性循环立体生态系统,充分利用了土地资源、发挥了生态效益与生产潜力,是一种高效农业物质循环的良性典范。随着时代变迁和科技发展,过度发展现代化高效养殖、极度追求经济效益以及生态保护意识淡薄等因素共同导致桑基鱼塘生态农业发展面临瓶颈,造成桑基鱼塘大面积萎缩、生态系统调节能力下降以及水产养殖安全和水域环境污染问题日益突出。目前,有关桑基鱼塘的研究仍局限于空间演变格局、经营模式评估、水陆相互作用过程与机理、投入产出效率、物质循环与能量流动等方面,而对桑基鱼塘内部营养物质的迁移转化规律缺乏全面了解。
鱼塘子系统是桑基鱼塘生产模式中的主要经济来源,其安全性和效益值得深入研究。N、P 元素作为鱼塘子系统中必不可少的营养成分,为鱼塘水产养殖提供大量的营养物质,但过剩的N、P元素会导致鱼塘水体富营养化等水质污染问题,从而造成鱼塘经济损失严重的后果。由于蚕沙与养殖鱼类投入和生物固氮等原因,大量的N、P营养物质,病原菌,寄生虫,有毒重金属和有机污染物等物质在水体和底泥中大量富集。因此系统梳理池塘子系统中N、P 的赋存形态,污染特征及迁移转化机制,对科学认识桑基鱼塘的微观特征和运行规律意义重大。
本综述通过梳理2010—2020 年桑基鱼塘的相关理论研究,以期从微观角度总结其鱼塘子系统N、P 营养元素的基本特征、赋存形态和物质循环规律,并解释其陆面子系统中水体-底泥界面N、P 营养元素的迁移转化机制及其影响因素。
1 桑基鱼塘生态系统鱼塘子系统基本特征
桑基鱼塘生态系统由陆面子系统和鱼塘子系统两部分构成,子系统间相互配合,相互补充,在桑基鱼塘生态内部实现物质循环和能量流动。具体模式表现为蚕排泄产生的蚕沙经过处理后进入鱼塘子系统喂养鱼类,每年鱼塘积累的塘泥又循环进入陆面子系统,为桑树的生长提供丰富的有机肥和营养养分,系统的组合伴随新的能量流动(图1)[1-2]。
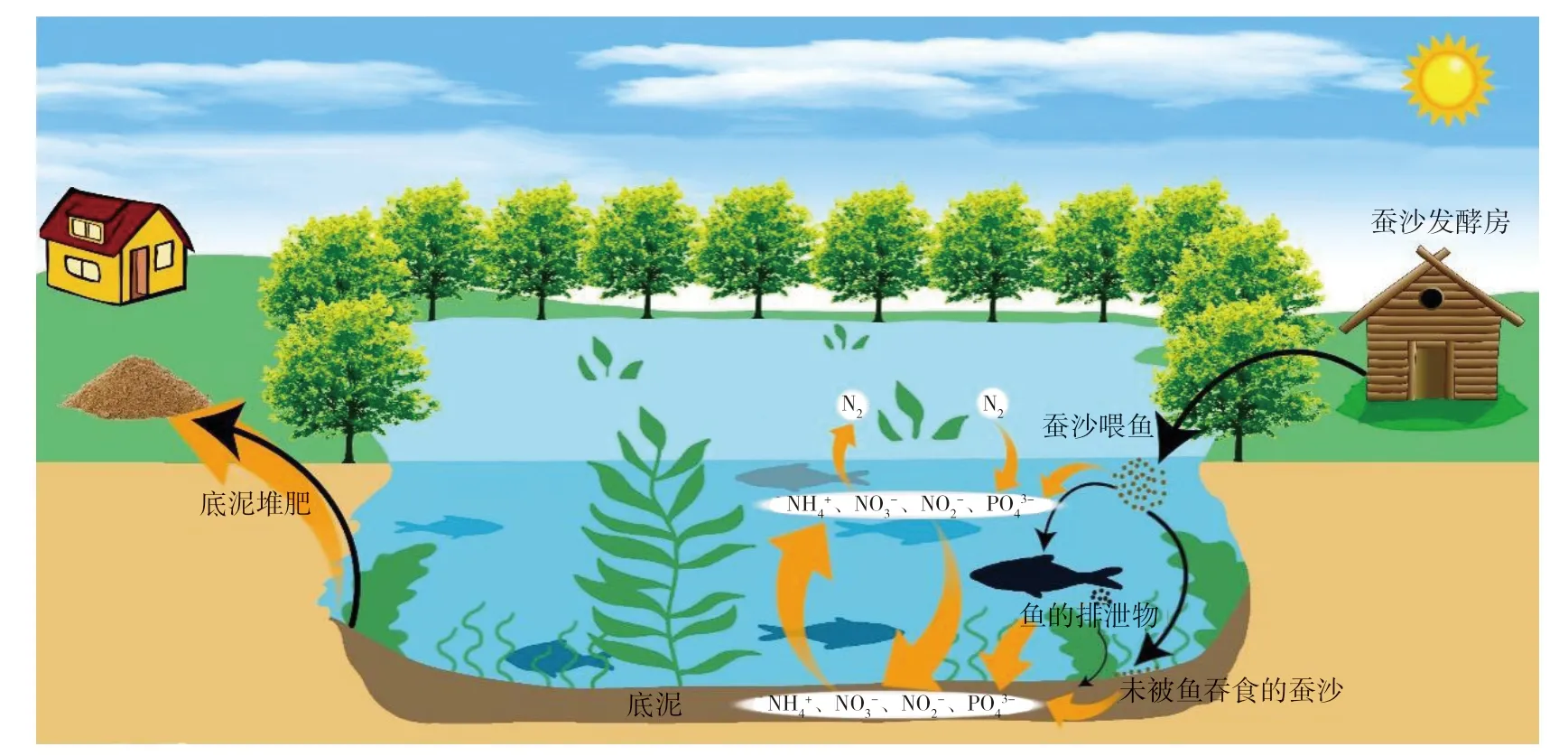
图1 桑基鱼塘池塘子系统基本模式Fig.1 Basic mode of mulberry-fish pond subsystem
鱼塘子系统是农户收入的最主要来源,在传统的桑基鱼塘经营模式中,鱼塘多采用立体混养养殖方式,即青鱼、草鱼、鲢鱼、鳙鱼分层养殖。随着混养养殖模式的演替,以及管理技术和方法的限制,目前管理者往往倾向于饲养单一鱼种,从而造成了桑基鱼塘内部养殖资源的低效利用,水产品的产出率低下。当前,鱼塘水产品产量的提升大多依赖于高密度、集约化的养殖模式。在大幅提高水产品的产出率和经济效益时,水体养殖环境的污染问题日益突出。研究表明,在养殖池塘中,鱼、虾等水产品的养分输出量不超过输入量的50%,剩余养分多以鱼类残饵、排泄物等形式沉积到底泥中[3]。
2 鱼塘水体和底泥中N、P 分布形态和基本特征
2.1 水体中N、P 的赋存形态特征
池塘水体是桑基鱼塘系统中营养元素的重要环境介质,给鱼类和其他水产品营造液体环境,直接提供丰富的溶解氧和营养物质。孙振中等[4]研究发现,养殖水体中总氮、总磷年平均质量浓度约为2.53、0.294 mg/L,但鱼塘中N、P 会在养殖过程中因有机质的累积在秋季养殖后期较高。因N 在水体中以氮气(N2)、硝态氮(NO3--N)、亚硝态氮(NO2--N)、氨态氮(NH4+-N)和有机氮5 种形态存在,在养殖水体中NH4+-N 的含量高于NO3--N 和NO2--N 的含量[5]。溶解于水的N2被水中的固氮菌和固氮蓝藻吸收作用,转变成可被水生植物利用的NH4+-N 和NO3--N。藻类直接吸收利用的NH4+-N、NO3--N 和NO2--N 等形式的N 被称为有效氮。绝大多数藻类总优先利用NH4+-N,当NH4+-N 消耗后才开始利用NO3--N,最后再利用NO2--N。因此,NH4+-N 是藻类生长中最重要的氮源,NH4+-N 含量不足通常成为水体初级生产力的限制因素[6]。
根据P 在天然水体中的物理性质和化学形态的不同,可将其分为可溶态磷(DP)和颗粒态磷(PP)[7]。其中DP 包括可溶态无机磷(DIP)和可溶态有机磷(DOP)。DIP 包括无机正磷酸盐和无机缩聚磷酸盐。正磷酸盐是浮游生物和细菌能直接吸收利用的磷形态[8],在pH 6.5~8.5的正常天然淡水中无机正磷酸盐的存在形态以HPO42-和H2PO4-为主。无机缩聚磷酸盐包括无机环状缩聚磷酸盐和无机线型缩聚磷酸盐或络合物[9]。DOP 主要包括植酸盐、磷蛋白、核蛋白、核酸、核苷酸、磷脂、磷酸单酯、磷酸双酯、肌醇和有机磷农药等[10-12]。天然水体中大部分DOP 可以被微生物群落和浮游动植物利用,将有机磷转化为正磷酸盐释放[13-15],碱性磷酸酶(APA)在这一过程中起重要作用[16]。因此,除了正磷酸盐外,溶解态有机磷在天然水体中也可作为初级生产者磷的来源[17]。PP 主要以矿物与无定形物质的共沉淀和吸附于矿物表面的形式存在,主要包含矿物颗粒的磷和无定形的磷、磷灰石态磷、颗粒有机物和生物体内的磷,是河流系统和河口地区磷的主要存在形态[18]。研究表明,随河流输入海洋的磷悬浮颗粒态占90%以上,径流中磷素输出以PP 为主,占总磷(TP)的比例超过75%[19]。
2.2 底泥中N 的赋存形态特征
底泥作为桑基鱼塘系统中营养物质的源和汇,沉积了大量残饵及鱼类排泄物,蕴藏丰富的营养物质成分。李煜等[20]研究湖区底泥表层中N、P 的浓度和分布现状,分别对底泥中总氮(TN)、总磷(TP)和总有机质(TOM)进行测定分析,结果表明,表层底泥TN 含量介于285~4 500 mg/kg 之间,均值为1 779.6 mg/kg,TOM含量介于17.13~189.30 g/kg 之间,均值为55.99 g/kg,空间分布与TN 具有相似格局;TP 含量介于261~709 mg/kg 之间,均值为639 mg/kg。N 是水生系统主要的营养元素,在底泥中以多种形态存在,且不同形态的N 对生物地球化学循环的贡献不同。底泥中N 分为有机态氮和无机态氮,有机态氮是底泥中N 的主要形态,组成复杂,以蛋白质形式存在的氨基酸为有机氮的主要形态;而无机态氮包括可交换态氮和固定态铵,可交换态氮是由无机氮通过离子交换等作用被吸附于有机质或矿物表面,可直接被水生生物吸收利用,是底泥-水界面水体-底泥界面N 交换的主要形态。一般而言,可交换态氮以NH4+-N 和NO3--N为主[21],其次为硝酸盐氮,亚硝盐氮含量最少。而固定态铵是指通过置换作用而进入矿物晶格内部的铵,结构稳定,是沉积物中主要的氮蓄积库,可在适宜的环境下释放并参与氮循环[21]。
有机氮占总氮含量的80%以上,是底泥中N 的主要形态,无机氮占总氮的比例较小。各形态无机氮以固定态铵含量最高,占总量的75%以上[22-23]。固定态铵是指非交换态的铵,通过置换矿物中的离子(如K+、Na+和Ca2+等)而被固定在矿物晶格中的铵,这种形态的N 也被认为是水生生态系统中N 的重要储存形式;与固定态按相比,相同物理化学条件下,可交换态氮更容易被初级生产者吸收利用,直接参与生命系统的循环,对研究水生系统具有重要意义。蒋国民等[24]发现,底泥中总氮(有机态氮和无机态氮)含量呈现由浅层向深层递增的趋势。有机氮在底泥中含量高,随着底泥中总氮含量的变化而变化;无机态氮以NH4+-N 和NO3--N 两种形态进行交换,交换的含量较低。有机氮是底泥中N 的主要形态,NH4+-N 和NO3--N 是底泥和上覆水之间进行N 营养物质交换的主要形式,固态氮则是底泥中无机氮的主要形式。
2.3 底泥中P 的赋存形态特征
底泥中的P 分为有机态磷和无机态磷,一定条件下,有机态磷会转化为无机态磷,而无机态磷以HPO42-、H2PO4-形态为主[25]。P 在底泥中存在形态复杂,研究底泥中P 的不同赋存形态及分布特征,有助于揭示水体中P 在水体-底泥界面的形态转化、迁移机制和生物地球化学循环过程。
P 对金属离子具有较高的亲和力,在一定环境条件下可与铁、铝、钙结合形成复杂的络合物[26-27]。Ruttenberg 等[28]提出将底泥中的P 分成5 种形态,分别是可交换态磷(E-P)、铁铝结合态磷(Fe/Al-P)、钙结合态磷(Ca-P)、有机态磷(OP)、残留态磷(residual-P)。其中可交换态磷一般附着在底泥表面,具有活性高、易被生物利用等特点,在浅层底泥中进行迁移。铁铝结合态磷是指与无定形和弱晶形铁矿物结合的磷,其活性受溶解氧质量分数影响较大。研究表明,Fe3+的还原可能引起内源磷的释放,当上覆水中磷酸盐浓度较低时,底泥中磷酸盐可从无定形铁矿物中释放[29]。在还原条件下,缺氧底泥中微生物可将胶体Fe(OH)3转化为溶解性Fe(OH)2,完成底泥中P 的吸附-释放过程。钙结合态磷可分为碎屑钙磷和自生钙磷,这部分P 在短时间内几乎不参与磷循环,难以被生物利用,被认为是持久性磷汇,仅在底泥环境为弱酸性时才可能会释放[30-31]。碎屑磷主要来源于水体中植物残渣,具有较强的稳定性,不易释放。有机P大多数来源于有机体细胞,以核酸、磷酸酯、磷蛋白等化合物的形式存在,其在矿化作用下可被分解为小分子态或可溶性磷酸盐。研究发现,瑞典Erken 湖底泥中50%~60%的有机磷可转化为生物有效磷的形态,因此可溶性磷酸盐常被用来表征磷的生物可利用性[32-33]。张小勇等[34]研究黄东海陆架区的表层底泥,发现其潜在的生物有效磷约占总磷13.55%,通过静态扩散及底泥再悬浮过程与水体发生物质交换及生物活化,进而影响水体初级生产力。
3 鱼塘子系统营养物质的释放行为与循环特征
3.1 水体和底泥中N、P 的释放行为
蚕沙等饵料投加到养殖水体后,未被鱼类完全摄食的残饵会悬浮于水体或沉淀到底泥中。饵料中蛋白质、糖类等有机大分子居多,并且经历矿化作用后会消耗大量溶解氧,从而导致水体-底泥界面形成厌氧环境。此外,矿化作用也产生N、P 等营养元素,进而促进水体中病毒和细菌大量繁殖[35]。饵料经过氨化作用后形成的NH4+-N 积存于底泥间隙水中,然后通过自由扩散进入上覆水。研究表明,饵料中蛋白质的含量与N、P 在水体中的释放量呈显著正相关,饲料蛋白质含量越高,N、P 释放量越高[33]。
当上覆水中的NH4+-N 浓度低于间隙水时,底泥就表现出N 释放,蕴含在间隙水中的NH4+-N 经过硝化反应向NO2--N 和NO3--N 转化,在反硝化作用下,NO3--N 被还原成N2进入上覆水并扩散到大气中。一般情况下,NO3--N 在水体中易被处于还原状态的表层底泥所还原,导致底泥中再生氮流失。
N、P 的释放速率具有相关性,且随着塘龄增长,底泥中N、P 呈现同步的释放规律。研究发现,底泥中N、P 的释放行为与底泥中的有机质含量相关,这表明N、P 的释放过程受底泥中有机质的分解作用驱动[36]。对于N 而言,有机氮是底泥中N 的主要形态,占总氮含量的98%以上。底泥N 在微生物作用下分解产生的NH4+-N,是底泥N 释放的主要形态。NH4+-N 通过交换吸附进入间隙水中,经扩散作用释放到上覆水体,最终在间隙水与上覆水之间形成动态平衡。厌氧条件下,底泥微生物可利用电子受体(如硝酸、铁和锰、硫酸以及二氧化碳等)来分解有机质。较高的氧化还原电位(ORP)条件下,底泥会分解产生大量NH4+-N。这归因于有机质的分解使大量NO3--N 作为电子受体被还原成NO2--N 和NH4+-N,NO2--N 浓度随ORP 下降而降低,说明当ORP 降低到一定程度后,微生物会选择NO2--N 作为新的电子受体,最终生成NH4+-N 进入底泥环境中。对于P 而言,PO43--P 是底泥中P 释放的主要形态[37],且与池塘运行时间无关。PO43--P 的释放与NH4+-N 释放机制稍微不同,P 除了和底泥有机质含量有关外,同时还受到底泥中Fe3+、Al3+等阳离子的影响。在微生物的参与作用下,底泥中有机磷被分解产生无机磷,无机磷顺着浓度梯度进入上覆水促进水体中P 的循环,位于间隙水中的P 被金属离子吸附后与水体参与反应,最终导致上覆水中的P 沉积于底泥中,表明在高ORP 条件下,底泥中PO43--P 会与Fe3+结合生成难溶性的Fe3PO4,抑制了P 的释放。
当水体为中性和碱性时,Fe(OH)3会吸附上覆水中的P;随着有机物的不断分解,底泥中ORP 下降,底泥中Fe3+被还原成Fe2+,进而导致被吸附的PO43-重新释放进入上覆水体。因此鱼塘长时间运行后,会出现P 释放速率急剧增长。
3.2 鱼塘子系统N、P 的来源
N、P 是池塘中浮游藻类生长必需的两种大量元素,通过调节水体的初级生产力,间接影响塘鱼产量。N、P 的收支平衡是池塘子系统物质循环的主要内容,可以从微观视角诠释桑基鱼塘生态系统的内在逻辑。因此,通过对生态系统中N、P 的收支平衡研究,不仅可以估算系统中N、P 的转化效率和养殖污染程度,还可以揭示其N、P 的来源和归宿。
养殖池塘中N、P主要来源为水体生物体的有机氮沉积及外源含N有机污染物的流入,底泥和上覆水中的有机氮大部分借助于微生物的矿化作用分解进入氮循环。输入来源具体包括饵料、肥料、养殖动物、降水、进水以及地表径流中N、P的输入。大量研究表明,饵料是池塘中N、P输入的主要来源。目前常见的桑基鱼塘生态系统混合养殖不同鱼类。在N、P输入方面,混养鱼塘全年N的输入总量为586 kg/hm2。N输入的主要途径是通过饵料携带(419.5 kg/hm2,71.57%),其次为进水输入(117.7 kg/hm2,20.08%)、苗种输入(43.68 kg/hm2,7.45%)和肥料输入(5.23 kg/hm2,0.89%)。混养鱼塘全年P输入量为114 kg/hm2。P输入的主要途径是通过饵料携带(86.03 kg/hm2,75.03%),其次为进水输入(14.79 kg/hm2,12.90%)、苗种输入(10.39 kg/hm2,9.06%)和肥料输入(3.45 kg/hm2,3.01%)[38]。
3.3 鱼塘子系统N、P 的归趋
研究表明,水产养殖生物对饲料的摄食效率为70%左右,剩余30%左右的饲料会以排泄物的形式排放到养殖环境中[39-40]。夏斌[41]研究发现,草鲢复合养殖池塘饲料中超过60%的N、P通过残饵、粪便和排泄物等形式进入养殖水体;王申等[42]研究发现,鲫鱼精养池塘的底泥沉积N、P 含量高达53%和87%;钟全福[43]研究发现,罗非鱼为主的罗非鱼-家鱼-对虾混养系统渔获物的N、P 含量占总输入量的45.45%和28.61%,其余大部分输入至养殖环境。陶冶等[44]发现不同养殖模式下进入池塘的N、P 营养盐都只有少数被养殖生物吸收利用,大部分未被摄食的营养盐会在水中溶解或者沉降进入底泥。潘红玺等[45]研究表明,相当大部分的饵料不能被鱼类有效吸收和利用,在池塘中形成残饵,加上生物碎屑和鱼类排泄等因素,导致底泥有机质含量逐渐增加,池塘子系统的污染负荷逐渐加大。
养殖池塘N、P 支出主要有渔获携带、排水、渗漏、挥发、底泥沉积等途径。在N 输出方面,通过渔获物携带输出165.1 kg/hm2、占总输出N 的28.17%,其次是排水输出(29.41 kg/hm2,15.29%)。通过其他途径输出的N 为248.7 kg/hm2,占总输出N 的42.43%。在P 输出方面,排水是P 输出的主要途径,为44.55 kg/hm2、占总输出P 的38.85%,其次是其他途径输出(35.82 kg/hm2,32.42%)和渔获物输出(34.29 kg/hm2,29.91%)。一般而言,底泥沉积是池塘N、P 输出的主要途径。此外,在物质输出的过程中还有N、P 的挥发和渗漏,这可能是N、P 通过其他途径输出的主要原因,从而造成鱼塘饵料中N、P 保留率分别达到28.94%和27.78%。底泥沉积是池塘养殖系统N、P 输出的主要形式,其输出量占总输出量的50%以上,其次是鱼获输出、占20%左右[46]。
4 N、P 在水体-底泥界面的迁移转化机制
严桦等[47]研究发现,南方河网地区猪鱼混养鱼塘及其附近河涌底泥的pH 值介于5.13~8.32之间,65.63% 的样品pH 值低于7.0,底泥主要偏酸性;有机质平均含量为17.94 g/kg ;全氮和全磷含量分别为0.393~4.155、0.191~9.111 g/kg。底泥中N 形态分为有机氮和无机氮,其中有机氮占绝大部分[48]。在厌氧环境下,厌氧氨氧化细菌利用亚硝酸盐为电子受体,将NH4+氧化成N2。同时,处于上覆水体中的NO3--N 和NO2--N 会迁移到底泥厌氧层中,经反硝化作用还原成N2O 和N2后散逸进入大气[49-50]。水体-底泥界面作为池塘子系统中N、P 循环的关键物质交换场所,N、P 迁移转化的环境行为受底泥性质、生物因素和环境因子影响。研究发现,天然湖泊中溶解态磷占比低于10%,绝大部分的P 以颗粒结合态悬浮在水相中[47]。界面条件发生变化时,底泥中的P 会再活化(解吸或溶解)释放进入上覆水中,在不同封闭水域系统中该过程的主导响应机制有所差异[51]。
如图2 所示,大部分有机氮经微生物降解为NH4+、NO3-等无机离子,在界面浓度梯度下向上覆水体释放。在溶解氧充足的情况下,水体中微生物活性较强且大量繁殖,同时伴随硝化作用。随着溶解氧的消耗,硝化作用产生的NO3-从上覆水中反向扩散到底泥缺氧层,NO3-在反硝化细菌作用下生成N2、N2O 等气体扩散到大气中。水体-底泥界面中硝化作用和反硝化作用耦合密切,不断消耗底泥中的有机氮和NO3-,这种脱氮过程可有效消除水体中内源N 负荷[26]。水体中颗粒态的有机磷经沉降作用聚集在底泥中,一部分在底栖生物和水动力的扰动下再悬浮重新进入水体中;另一部分有机磷经矿化分解生成磷酸盐,其中溶解性磷酸盐具有较强生化活性能够直接被水生植物和微生物所利用,当生物死亡后,体内的可降解性有机磷在生物化学作用下释放,继续参与水体中的磷循环,同时水体中磷酸盐可被底泥中活性Ca2+、Fe3+吸附结合成钙结合态磷和铁结合态磷。与其他形态磷相比铁结合态磷稳定性较差,当底泥中氧化还原条件改变时,易发生解吸现象。
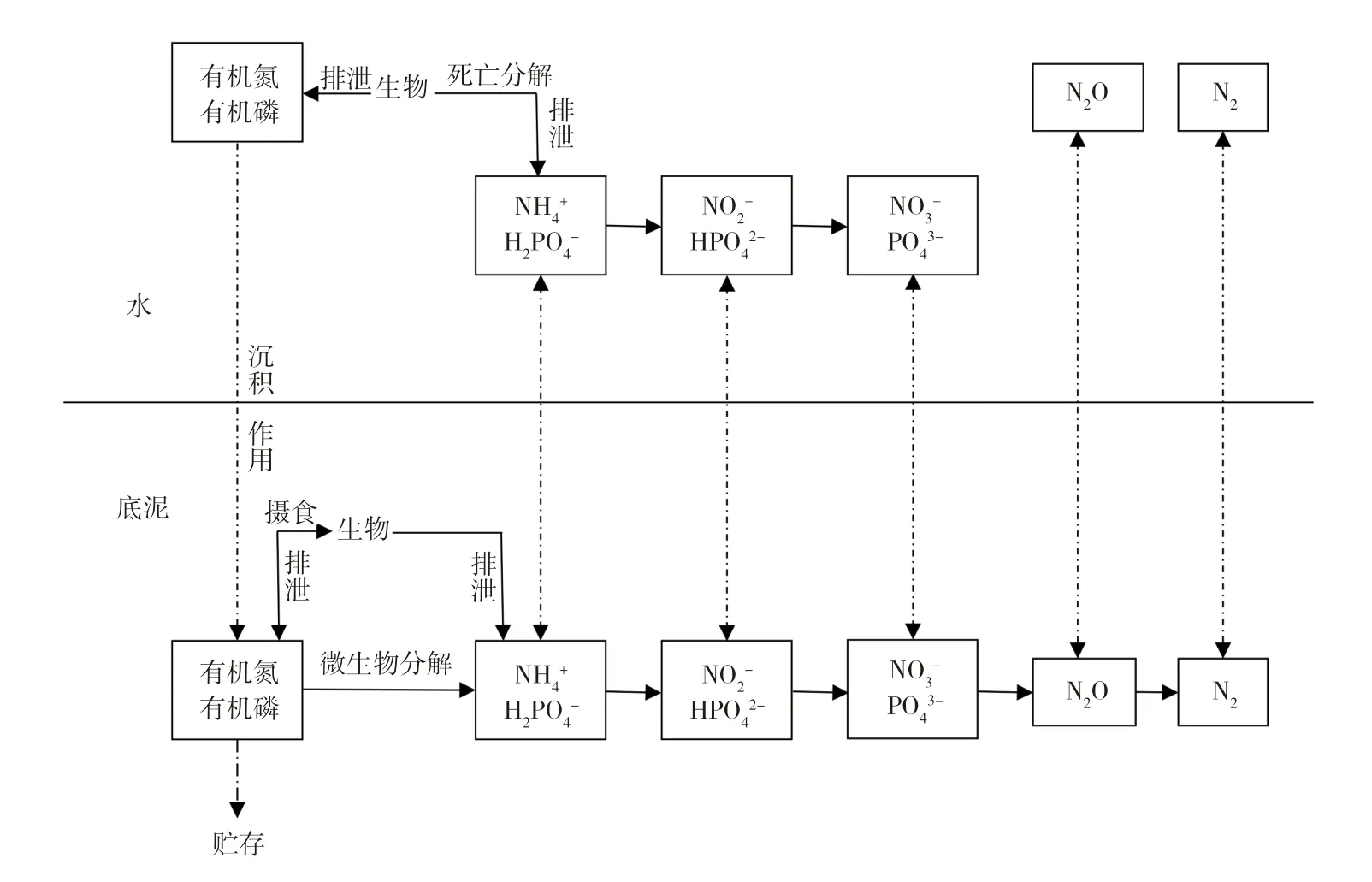
图2 N、P 在水体-底泥界面的迁移转化机制Fig.2 Migration and transformation mechanism of nitrogen and phosphorus at water-sediment interface
4.1 底泥性质对水体-底泥界面N、P 迁移转化的影响
底泥性质受有机质组成、粒径结构、微生物群落和金属离子等物质影响,并且底泥的组成结构、浓度均会影响水体-底泥界面N、P 的迁移转化过程。底泥颗粒再悬浮的程度受湖泊地形和水动力循环影响,底泥颗粒越细,总氮、总磷和有机质含量越大,比表面积越大,可转化态氮的含量越高,无机氮的含量则越低,吸附特性更好,对N、P 在水界面交换的影响存在差异。研究发现,底泥中总氮、总磷、有机质以及各形态磷中85%以上分布在细颗粒中,而无机磷在黏粒、细砂粒、粗砂粒以及粉砂粒中的含量逐渐减少[52-53]。不同深度底泥中颗粒物以无机磷为主,可交换态磷的含量相对较低,粒径较细的底泥可吸附较多的可交换态磷[54]。李乐等[55]发现底泥间隙水中磷浓度与黏粒、粉砂粒的丰度呈显著负相关,表明底泥细颗粒物含量越高,间隙水P 含量越低,P 的释放越少。
底泥中P 的释放速率与有机质含量呈显著正相关,有机质含量会影响底泥P 的最大释放速率和释放量[56]。底泥中的有机质包含脂肪酸、腐殖质和蛋白质等成分,主要来源于湖泊中藻类、动植物残体。有机质矿化作为湖泊磷循环的重要驱动力,其产物会不同程度影响P 的吸附速率和吸附量,如腐殖质能在无机物表面形成胶膜,减缓P 的迁移速率;富里酸会与磷酸盐形成竞争吸附,使P 从底泥向上覆水迁移。底泥微生物通过矿化分解有机磷和分泌有机酸会降低pH 值,促进底泥中P 的释放。钱燕[57]研究发现,灭菌处理的底泥会吸附上覆水的P 素,而未灭菌的底泥由于微生物的活动促进底泥P 的释放。目前,铁磷耦合作用被认为是调控P 迁移的主要机制,即Fe3+被还原会促进底泥中P 的释放。此外,有机质还能与Fe、Al 等金属形成络合物复合体,占据P 的结合位点,抑制P 的释放。
4.2 生物对水体-底泥界面N、P 迁移转化的影响
池塘内蓝藻、硅藻等浮游植物的生长可以同化吸收上覆水中的营养元素,降低水相中N、P的浓度,促进底泥中N、P 的释放。此外,浮游植物的生长也会引起上覆水pH 值升高和氧化还原电位下降,并能改变底泥微生物的群落结构,促进底泥中N、P 向上覆水迁移。
一般而言,水体中藻类的生长与底泥中P 的释放过程两者相互促进[24]。底泥微生物是池塘子系统中能量循环的主要驱动者,对水体氮循环和P 元素的价态转化过程起不可替代的作用。章海鑫等[58]研究发现,草鱼养殖池塘中的底泥微生物主要是变形菌门、厚壁菌门、放线菌门、硝化螺旋菌门以及分类地位未定的种类。张植强等[59]研究发现,草鱼池塘中的厌氧氨氧化菌在厌氧条件下以NO2-为电子受体把NH4+氧化成N2,并通过硝化作用把氨氧化成硝酸盐,降低水体氮污染以及游离态氨和亚硝酸盐的含量。微生物的生命活动可促进底泥中P 的释放,将有机磷转化为H2PO4-、HPO42-和PO43-等无机磷,被藻类吸收后同化为有机磷。研究发现底泥释放的大部分P 会被藻类重新吸收,但藻类的生长与N、P 的摄取过程并不同步,藻类吸收P 素后将其储存在体内,维持其在低P 环境中的生长[60]。P以有机磷的形态储存在生物体中,随着浮游植物的衰亡,P 重新释放进入上覆水参与磷循环过程。
水生植物对水体-底泥界面P 迁移的调控机制种类繁多。由于生长习性特殊,沉水植物会促进底泥中有机物的矿化,降低底泥中N、P 含量,影响不同形态N、P 的迁移转化。马久远等[61]发现沉水植物区域的底泥中,总氮、有机氮、氨氮和硝态氮含量均比群落外有不同程度降低,其中总氮和有机氮含量下降幅度明显。同时,沉水植物抑制沉积物中氮向上覆水体释放。研究表明,苦草与菖蒲都能降低沉积物中P 的迁移速率,但菖蒲对总磷的抑制效果相对更好,这是由于菖蒲的根系能为微生物与菌胶团营造适宜生境,抑制了PP 和DOP 的释放;苦草通过同化吸收DOP、改变溶解氧浓度和pH,影响水体-底泥界面P循环[62]。易文利等[63]发现穗花狐尾藻的生长会促进底泥中活性有机磷和NaOH-P 的释放,而衰亡过程则以有机磷的沉积为主。研究发现,黑藻能显著降低底泥的总磷含量,但对P 也起到了活化作用,增加可交换性磷的含量,但这种分歧可能与沉积物中P 的形态和构成有关[64-65]。
此外,底栖动物的爬行、摄食等生理活动也会影响底泥的理化性质,进而影响水体-底泥界面的P 迁移。常见的底栖动物有摇蚊幼虫、水丝蚓和河虮等,如摇蚊幼虫能增加底泥中溶解氧的渗透深度,促进Fe(OH)3生成,从而抑制底泥中活性P 的释放。水丝蚓活动会增加沉积物含水率和氧化还原电位,研究发现水丝蚓对太湖梅梁湾和大浦口沉积物P 迁移作用效果相反,可能是因为大浦口间隙水Fe2+浓度是梅梁湾的数倍。但有学者发现河蚬扰动促进了大浦口沉积物活性P 的释放,这是因为河蚬扰动促进了有机质和含P 代谢产物的分解,这些过程释放的P 远多于Fe(OH)3吸附的P[66]。
4.3 环境因子对鱼塘子系统N、P 迁移转化的影响
4.3.1 水体温度对鱼塘子系统N、P 释放的影响温度是影响N、P 释放的主要因素,通过调控水体-底泥界面的物质迁移速率和改变微生物活性来影响底泥中N、P 的释放。研究表明,低温会抑制N、P 的释放,高温会促进N、P 向上覆水体释放[67]。N、P 释放行为对温度变化的影响规律类似,N 的释放量并没有P 的变化明显,这可能是温度持续升高,而生物活性不再提高,造成N 的释放速率增长平缓。
温度会影响氨的形态和吸附特性,由于氨的吸附是一个放热过程,升高温度会降低其吸附性能,导致底泥吸附能力下降。此外,氨主要以NH3和NH4+两种形态存在于水体中。当温度升高时,由于微生物参与了水体-底泥界面N 迁移转化过程中的多个关键反应,如氨化、硝化和反硝化反应,使得水体中NH4+的浓度增大,导致水体中NH4+通过水体-底泥界面向底泥迁移[68]。大部分微生物对温度的变化相当敏感,温度可以通过改变微生物的活性抑制或者加速不同形态氮的释放速率。当温度升高时,一方面微生物氨化能力增强,有机氮快速氨化生成NH4+-N,另一方面强烈的微生物活动减少了溶解氧含量,硝化作用受到抑制。此时底泥间隙水中NH4+-N 的浓度远大于上覆水体,致使以NH4+-N 为主的无机氮由底泥向上覆水体释放。当温度降低时,微生物活动受到抑制,NH4+-N 的扩散减弱,硝化作用占主导作用,NO3--N 成为底泥-水界面N 素扩散的主要形态,并且水体-底泥界面的N 素释放整体减弱。研究表明,夏季(25℃)太湖中N 素是由沉积物向上覆水释放,底泥作为释放源,而冬季(5℃)则表现为上覆水向底泥转移[43],即底泥表现为汇[69]。魏羽含[70]研究发现,随着水温从5 ℃上升至25 ℃,总氮浓度从8.5 mg/L 上升至25 mg/L。然而,当温度持续升高到一定程度后,水体中N 的含量变化不明显。由此可见,温度的改变能影响氮循环的整个过程。
水温的升高有利于沉积物向水体中释放P,温度越高,P 的释放越明显。研究发现,其他条件相同的情况下,当水温从10 ℃上升至25 ℃时,上覆水的总磷浓度从0.06 mg/L 上升至0.09 mg/L[71-72]。这归因于高温使得微生物代谢活动增强,造成水体溶解氧含量下降,致使水体氧化还原电位降低,促进了Fe3+向Fe2+方向转化,导致吸附在Fe(OH)3胶体及磷酸盐中的P 重新释放出来,造成了Fe-P 的释放。此外,高温会促进微生物的代谢和藻类的降解过程,加快一些含P 有机物的分解。由于有机质矿化过程会产生有机酸,与铁、铝等金属通过螯合作用,促进金属结合态磷的释放。一般而言,高温下底泥中P 的释放量高于低温,这可能由于池塘在高温季节富营养化程度加强所致。Sendergaard 等[73]对丹麦一些浅水湖泊的研究也得到了类似规律,发现夏季上覆水的总磷浓度是冬季的2~4 倍。
4.3.2 水体pH 值对鱼塘子系统N、P 释放的影响 水体的pH 值能显著影响底泥-水界面营养盐释放行为,通过改变水环境中N、P 的物理吸附、化学转化过程和微生物活性,可对水体-底泥界面各形态N、P 的迁移转化过程造成影响。研究表明,当底泥上覆水的pH 值为酸性或碱性时,氨氮和总磷的释放强度都会增大[74-75]。
pH 值与TN 浓度呈显著负相关,酸性或碱性条件下都不利于底泥中微生物(如氨化细菌、硝化细菌等)的生命活动,会抑制底泥中N 的释放。在中性或弱碱性的环境下,微生物的活动最为活跃,有利于N 素释放。当pH 值较低时,游离态H+易与铵盐中NH4+发生竞争吸附,使NH4+在铵盐的结合位置被取代后释放进入水体;而当pH值较大时,OH-会与NH4+发生反应,NH4+转化为NH3,并以气体的形式脱离水体,因此N 含量较小。吕拥军等[72]对比研究了pH 5、7、9 下N的释放通量,发现上覆水体的总氮含量在16 d 后基本稳定,N 释放随着pH 值的升高而减小。高湘等[76]研究发现,中性条件下底泥中氨氮释放量达到最高,当pH ≥8 时氨氮的释放量随pH 值增大而迅速减小。
李晔等[77]研究表明,在酸性范围内,底泥中总磷处于负释放状态,这是由于矿物表面被质子化,促进了P 的吸附,即水中H+浓度越高,底泥吸附P 的能力越强;在碱性范围内,底泥中总磷处于正释放状态,离子交换是影响总磷释放的主要机制,体系中的OH-与金属离子结合的磷酸盐发生交换,使得HPO42-和H2PO4-的释放强度增大。因此,随着pH 值升高,底泥中P 的释放量增大;大量游离的OH-离子与底泥胶粒的阴离子竞争吸附位置,迫使底泥中的P 以HPO42-和H2PO4-的形态释放出来[78]。而在中性范围内,P的主要存在形式是HPO42-和H2PO4-,易与底泥中的金属元素结合,从而被底泥吸附,此时总磷的释放强度较小[79]。整体而言,pH 值对底泥总磷释放强度的影响大小表现为碱性>中性>酸性。
4.3.3 底泥对池塘子系统N、P 释放的影响底泥的厚度也会对池塘N、P 的释放造成影响。NH4+-N 是底泥中N 最主要的形态,鱼塘刚形成时,水中NH4+-N 水平比较低。随着鱼塘养殖时间的积累,底泥逐渐变厚,NH4+-N 的持续释放能力增强。研究表明,底泥厚度与水中NH4+-N 浓度显著正相关[80-81]。因此,鱼塘需要定期清理塘泥,将含有大量营养元素的塘泥用于桑树供肥,有利于避免水体富营养化,导致鱼因缺氧死亡。残饵与塘鱼排出的粪便沉积到底泥中,造成底泥中有机质的积累,而底泥中有机质分解的产物与金属离子结合后促进磷酸盐的溶解,造成HPO42-和H2PO4-等基团从底泥中释放进入上覆水中。
李程亮[67]通过模拟实验研究了底泥厚度对底泥中N、P 释放特性的影响,不同厚度底泥的TP 和SOP 的释放强度表明,模拟系统运行5 d 后,5、10 cm厚的底泥TP和SOP的释放强度降幅较大,而15 cm 厚的底泥TP 和SOP 的释放强度降幅较小;运行2 d 后,10、15 cm 厚的底泥TP 和SOP的释放强度相当,均明显高于5 cm 厚的底泥TP和SOP 的释放强度;综合来看,底泥TP 和SOP的释放强度排序依次为15 cm >10 cm >5 cm,说明底泥越厚越有利于P 素留存在水体中。因此,在实际水产养殖施肥过程中,定期清除过厚塘泥是避免池塘子系统水体富营养化的关键。
5 结语
桑基鱼塘是一种历史悠久,兼具经济、生态、社会文化的混合生态农业生产方式,其主要利用基面与池塘构成水陆交互作用的农业系统,从而实现水产养殖、桑种植及各类加工业相结合的一种生态型农业。随着科技和工业文明的发展,桑基鱼塘的面积逐渐缩小,并且传统桑基鱼塘景观遭到破坏,经济效益大不如前。本综述梳理相关文献,明确桑基鱼塘生态系统鱼塘子系统的基本特征、鱼塘中水体和底泥中的N、P 分布形态和基本特征,鱼塘子系统营养物质的释放行为与循环特征,以及N、P 在水体-底泥界面的迁移转化机制,旨在探究桑基鱼塘内部物质循环、能量流动规律和迁移转化机制,为构建桑基鱼塘高效新型生态利用新模式,促进特色生态农业系统可持续发展提供理论依据。
桑基鱼塘农业生产模式历经千百年来的形成、发展和延续至今,仍具有巨大的经济、文化和科学价值,不仅具有农业生产功能,同时承载了一个地区的历史发展和文化。目前,大多数学者研究专注于桑基鱼塘的生产、管理和经营等经济文化发展方面,但对于桑基鱼塘内部营养物质的迁移转化规律缺乏深入了解,并且现阶段桑基鱼塘发展缺乏动力、面临发展困难。因此,我们不能仅关注桑基鱼塘表面产生的经济效益,更应该从桑基鱼塘历史悠久的发展进程、社会、文化和生态方面,探索新的发展生机。因此迫切需要更多相关专家学者的关注和开展相关研究,并从更深层次揭示桑基鱼塘生态系统内部潜在的科学规律,从而为桑基鱼塘的传承和保护提供参考依据和解决方案。
